ABSTRACT
This study sought to understand the importance of variation of steeping and germination conditions (temperature, pH and salts) on the quality of African finger millet malt in terms of diastic power (a-amylase and β-amylase), endo- (1,3) (1,4)-β-D-glucanase, b-glycan content and protein profile. The results show that the physiological responses of African finger millet malted seeds are correlated to pH (acidity and alkalinity) but inversely correlated to temperature stress. The effect of the stresses on the activity of a-amylase, b-amylase and endo-(1,3)(1,4)-b-D-glucanase as well as b-glycan content was significantly different in magnitude except for the β-amylase activities obtained after acidic and alkaline treatment at 40°C which are not statistically different. Alkaline pH and heat stress at 30°C were the dominant factors for malting optimization from the result of diastic power indices. a-Amylase activity is a better predictor of diastic power. The grains subjected to the steeping and germination process carried out in Tris-HCl buffer solution (25 mM, pH 9) containing 100 mM NaCl at 30ºC during 96 h showed higher α-amylase and β-amylase activity. This shows that for a salt–alkali-heat mix stress, a reciprocal enhancement among salt stress, alkali and heat stress was a characteristic feature with no significant change in the hordein protein expression. The influential effect of the stress conditions indicate that alkaline pH steeping and 30°C malting is the most effective condition for producing malted African millet flour with a promising potential of distinct malting quality metrics.
Key words: African finger millet, diastatic enzymes, malting, stresses.
African finger millet (Eleusine coracana (L.) Gaertn. subsp. coracana) is an important cereal in West Africa and India. African finger millet is grown mainly by subsistence farmers in Nigeria and serves as a food security crop because of its high-nutritional value, flavor and excellent storage qualities (Mbithi-Mwikya et al., 2000). It is used either in its native or processed forms in the preparation of weaning foods, pharmaceutical products and low cost ready-to-eat food items (Narayanaswamy et al., 1971). African finger millet sprout flour has been used as a source of enzyme for production of togwa, a traditional nonalcoholic beverage consumed in East Africa (Yasuyuki and Kitabatake, 2003).
Malting of African finger millet is a well-known household technology in West and East Africa and India (Gimbi and Kitabatake, 2002). The traditional malting technique of African finger millet and sorghum are similar (Nout and Davies, 1982). This involves 24 h continuous immersion steeping, 2-3 days germination and lastly 1 or 2 days solar dehydration at 25-30°C. This has posited to improve the nutritional quality and reduce the level of anti-nutrients in African finger millet flour (Mbithi-Mwikya et al., 2000). Malting is the controlled and partial germination of cereals grains under conducive to control environmental conditions of steeping, germination and kilning (Mark et al., 2013). The biotechnology of malting uses the natural germination events to provide favourable mashing material hence, intact enzymes, starch availability and viability of the seeds are the main criteria for malt quality (Gorzolka et al., 2012). The primary objective of cereal malting is to promote the development of hydrolytic enzymes. However, some malted cereals may contain low activity of starch-hydrolysing enzymes (β-amylase and α-amylase) and low activity of desirable enzymes degrading endosperm cells walls like β-glucanase (Agu and Palmer, 1997). In addition, the possibility of successfully eliminating specific fractions of non-digestible soluble fibre such as β-glucan, from different types of cereals or cereal by-products has also posed challenges to maltsters (Charalampopoulos et al., 2002). The findings of Hounhouigan et al. (2007) showed that malt enzymatic power is highly variable with the variety of grain in consideration and the malting (steeping, germination and drying) conditions employed. The joint amylolytic activity of primarily β-amylase and α-amylase, termed diastatic power, is a major malting quality parameter of malted products (Clancy et al., 2003). In the same vein, the extent of endosperm cell wall degradation by the β-glucanase enzymatic activity and the β-glucan content left after processing are vital determinant factors of malting quality especially in the friability of the malt product (Wang et al., 2004). The protein content of malted products is also pivotal to the determination to their quality (Jamar et al., 2011). The full knowledge on these parameters and conditions which bring about their improvement is very important to maltsters. The quality of the malt is of primary significance in the manufacture of brewed products. To increase the brewing yield and efficiency, malts with high extract values, high enzymatic activities, and good modification are essential. To produce malt that meets these requirements, the grains are expected to be able to germinate rapidly and uniformly (Riss and Bang-Olsen, 1991), producing a correspondingly high diastatic power, high β-glucanase activity and low protein content. Lately, attention has been placed on optimization of African finger millet malting process for quality malt (Gimbi and Kitabatake, 2002). In Nigeria, attempt is underway to use malted African millet for beer production. This prompted a high and incessant demand for malted African finger millet flours for beer brewing, preparation of non-alcoholic beverages, weaning foods, infant foods formula (Nzelibe and Agbojo, 2005; Hounhouigan et al., 2007) and for the development of novel foods in cereal based industries (Charalampopoulos et al., 2002).
The study seeks to understand the importance of variation of steeping and germination conditions (tempera-ture, pH and salts) on the quality of African finger millet malt in terms of diastic power (a-amylase and β-amylase), endo- (1,3) (1,4)-β-D-glucanase, b-glycan content and protein profile. The study revealed that careful control of germination environments and conditions coupled with monitoring and optimizing diastatic enzymes (a-amylase, β-amylase), β-glucanase activities and reducing the β-glucan and protein contents, one will be able to reliably produce malt with finely tuned characteristics as well as an appropriate physical state that together meet the needs of their brewing customers.
African Finger Millet (Eleusine coracana) (local variety) was acquired from the Farin Gada market, Jos, Nigeria in September 2011. The seeds (moisture content of 11g/100 g and germination capacity of ≥ 92%) were screened to remove broken grains and foreign materials and then stored at £ 4°C until used. Enzyme activity kits of a-amylase (K-CERA), β- amylase (K-BETA), endo- (1,3) (1,4)-β-D-glucanase (K-MBGL) and β-glucan (K-BGLU) were obtained from Megazyme International Ireland Ltd., Bray Business Park, Bray, Co. Wicklow, Ireland. All other materials used were of analytical grade from authorized commercial suppliers. All spectrophotometric assays were monitored using Shimadzu UV 1800 double beam UV–visible spectrophotometer. All measurements were performed thrice unless otherwise stated.
The malting procedure
The micro-malting was done as reported elsewhere (Macnicol et al., 1993). The grains were then weighed (100 g per group), washed (in distilled water) and sterilized using 1% sodium hypochlorite solution for 20 min at room temperature The malted grains were pulverized using a thermoline® blender.
Design of simulated acidic, alkaline and salt conditions
The sterilized grain samples, in their respective groups were steeped overnight at 25°C under acidic pH stress condition (25 mM Glycine-HCl Buffer, pH 3.0), alkaline pH stress condition (25 mM Tris-HCl, pH 9.0) and salt stress condition (0 - 200 mM NaCl, a neutral salt, in deionized water). The grains steeped in distilled water serve as a control. The grains were germinated at these stress conditions under constant moisture content at either 30 or 40°C (room temperature was 25°C). Sampling was at 24 h for 96 h and immediately kept at -20°C until used.
Measurement of diastatic enzymes indices
Malt a-amylase, b-amylase and β-glucanase activities were measured according to the enzymatic method of a commercial assay kit (Megazyme International Ireland, Bray, Ireland). The results are expressed as U/g. Total β-glucan content in malted finger millet was measured according to the enzymatic method of McCleary and Shameer (1987) using a commercial assay kit (Megazyme International Ireland, Bray, Ireland). The calculation of the β-glucan content of the malt was carried out using the Megazyme Mega-CalcTM (www.megazyme.com).
Protein content assay and SDS-PAGE
Total protein concentration was determined according to the dye binding method of Bradford (Bradford, 1976) with bovine serum albumin as standard. Aliquots of the millet protein extract were taken from the 96 h germinated samples for SDS-PAGE. This was achieved by mixing 100 mg of each germinated millet sample with 1.0 mL of 20 mM sodium phosphate buffer (pH 6.5) under mechanical stirring for 3 h. The mixtures were centrifuged at 10, 000 g for 10 min at 4°C. 400 μL of the supernatants containing protein extract were precipitated with 100% trichloroacetic acid (TCA) in cold acetone 1 g/ml (w/v) and stored overnight at 0°C. These were later centrifuged at 10, 000 X g to collect the pellets formed. The pellets were washed repeatedly with cold acetone to remove all TCA and then air-dried. The obtained residue was dissolved in electrophoretic sample buffer containing 20 gL−1 SDS, 0.0625 mol L−1 Tris, 100 mL L−1 glycerol and 0.1 gL−1 bromophenol blue (pH 6.8) and boiled at 100°C for a minute. The hordeins were similarly extracted three times for 1 h each with 55% (v/v) propan-2-ol, 1% (v/v) acetic acid and 2% (v/v) 2-mercaptoethanol (15–30 ml) at 60°C. The hordein fraction was reduced with 1% (v/v) 2- mercaptoethanol and alkylated with 1·5% (v/v) 4- vinyl-pyridine to disrupt permanently the disulphide bonds within the B and D hordeins. The discontinuous Tris-HCl/glycine buffer system was employed (Laemmli, 1970), with 12 % SDS-PAGE.
Gels were scanned and the digital pictures subjected to densitometric analysis using the Gene Tools software 3.07.03 Package (Synoptics, Cambridge, England).
Statistical analysis
Data from triplicate results was presented as mean values ± standard deviation (SD). The results were statistically evaluated by one way analysis of variance analysis (ANOVA) followed by Duncan's New Multiple Range Test (DMRT) using SPSS 16.0 (SPSS Inc., Chicago, IL, USA)statistical package. LSD test as post hoc tests with a p = 0.05 significance, to compare the treatment effect. Graphical analysis was performed with performed using Kaleidagraph 4.1 (Synergy software).
α-Amylase
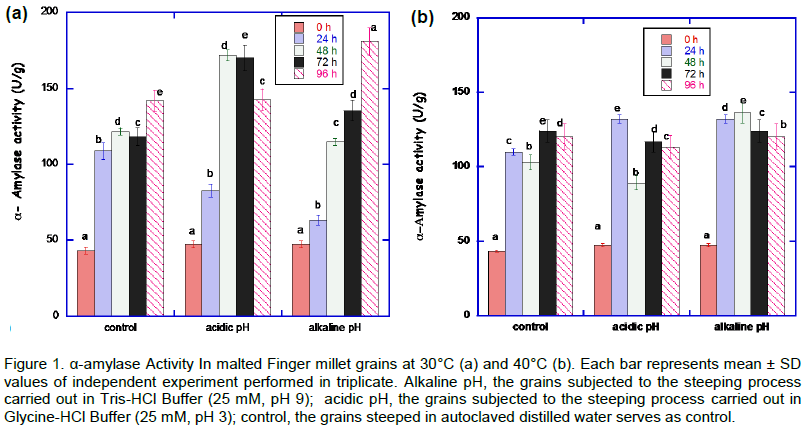
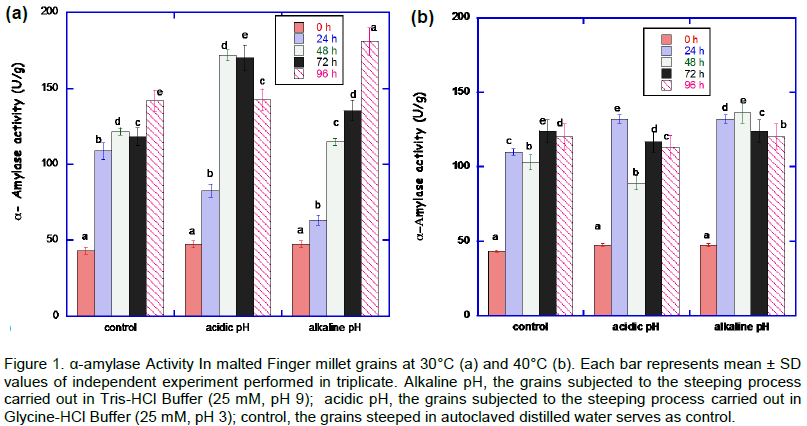
The result of α-amylase activity malted African finger millet during various stress condition is shown in Figure 1. A steady increase (from 0 to 96 h) in the α-amylase activity was noticed at 30°C at alkaline pH stress condition. The highest activity (180.04±0.20 U/g) was observed at 96 h for alkaline pH steeped and germinated at 30°C. At 40°C, the highest activity (136.11 ± 2.02 U/g) for the malted Finger millet was at alkaline pH at 48 h.
β-Amylase activity
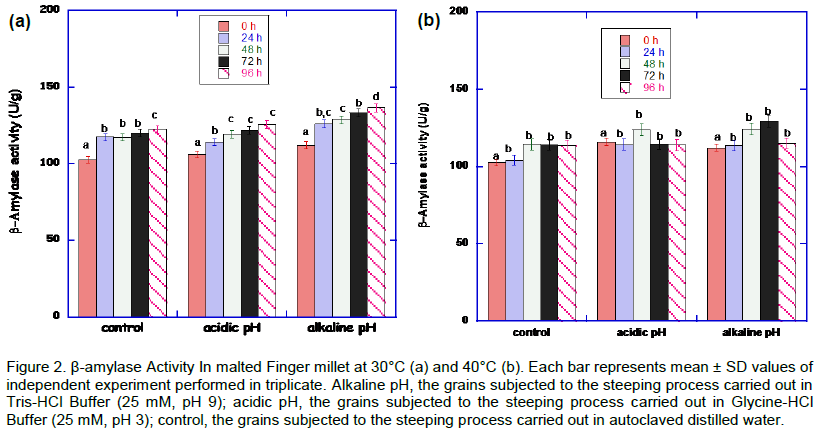
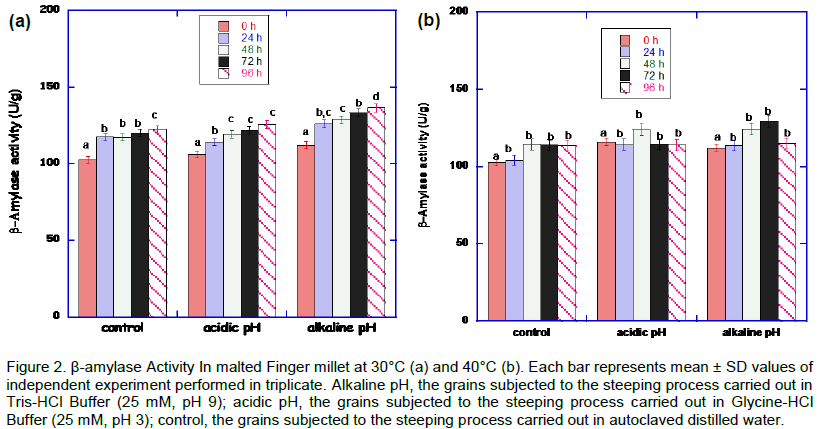
The results of the effect of pH and heat stresses on β-amylase activity is shown in Figure 2. There was increase in the activity at alkaline and acidic pH at 30°C. The optimum β-amylase activity (136.27 ± 2.0 U/g) was at 30°C at alkaline pH condition at 96h. At 40°C germinating condition, there was a decline in β-amylase activity after 48 h and 72 h at acidic and alkaline pH stress conditions, respectively.
β-Glucanase content
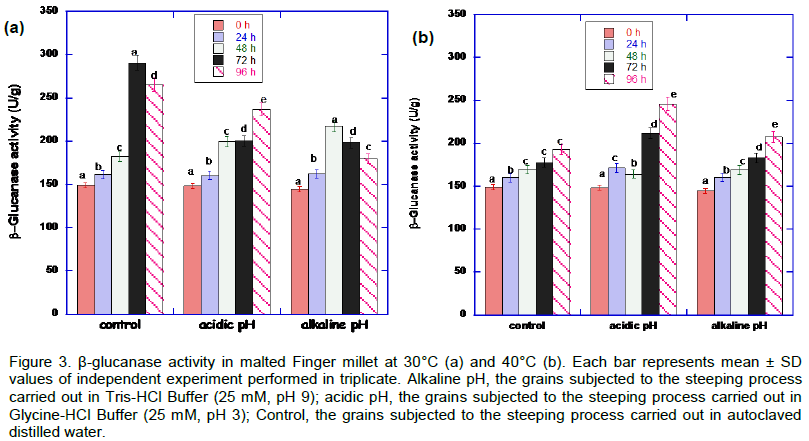
The result in Figure 3 shows the effect of heat stress and pH stress on β-glucanase activity in African finger millet malt. Malting at 30°C showed optimum β-glucanase activity compared with 40°C. There was statistically significant increase for pH stress conditions from 0 to 48 h period.
β-Glucan content estimation
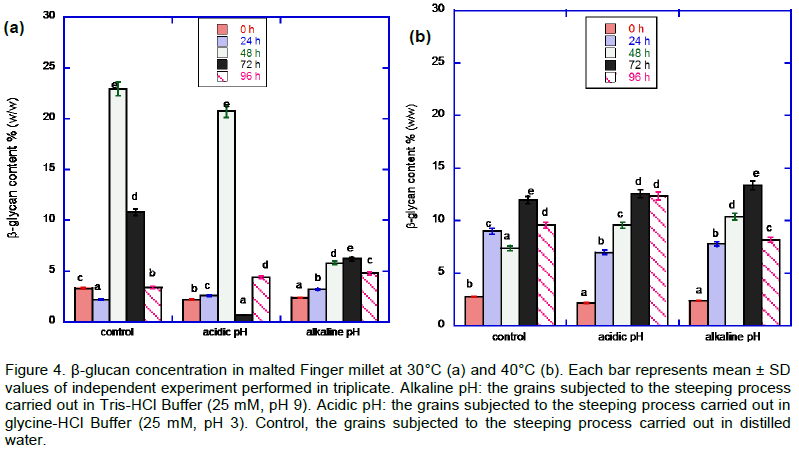
The result of the effect of heat and pH stresses on the β-glucan activity of the malted African finger millet grain is presented in Figure 4. There was a general increase in the β-glucan content at 40°C compared to 30°C.
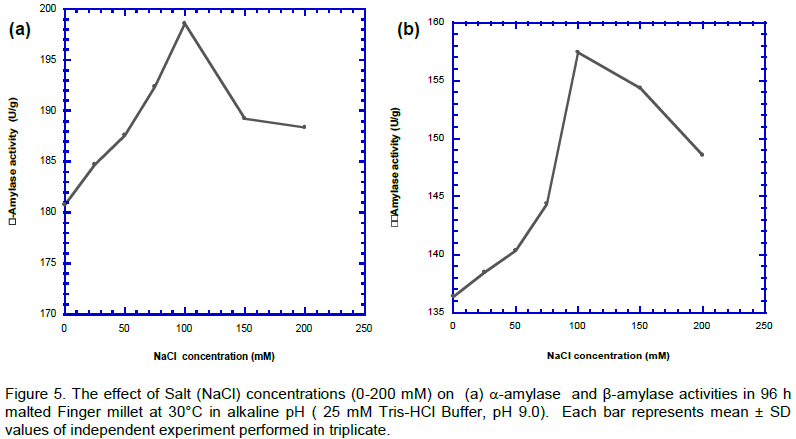
Salinity treatment
The simulated salt–alkaline-30°C stress conditions were a combinatorial approach employed to study the complex salt–alkaline (30°C) stress conditions in malting (Figure 5). a-Amylase assay was used as indicator, because of its sensitivity, to monitor the effect. The result indicates an increase in the a-amylase activity from 0- 100 mM but no further significant change till 200 mM. b-amylase activity was further used to revalidate the result of the a-amylase activity. The result showed to be the same.
Protein profile of malted Finger millet using SDS-PAGE
SDS-PAGE separation of hordein polypeptides from individual 96 h malted African finger millet grains in alkaline pH at 30°C, alkaline pH at 40°C, acidic pH at 30°C and distilled water at 30°C is shown in Figure 6. The hordein fraction expressions were not significantly different from the stress conditions applied. It is predominantly characterised by 44-66 kDa hordein and 20-27 kDa hordein fractions. This indicates hordein B and hordein A respectively.
There has been a high and constant demand for malted cereals for beverage production, infant foods formulation and for novel food development in cereal based ndustries (Charalampopoulos et al., 2002). However, the problems usually encountered are inadequate expressed enzyme levels of starch-hydrolysing enzymes and endosperm cell wall degrading enzymes (Agu and Palmer, 1997). In addition, the possibility of successfully eliminating non-digestible β-glucan fractions from different types of cereals or cereal by-products has also posed challenges to industrialists and technologists (Charalampopoulos et al., 2002). Malt enzymatic power is highly variable within
the varieties of cereals grain and the malting (steeping, germination and drying) conditions employed (Hounhouigan et al., 2007; Ukwuru, 2010).
The acidic and alkaline pH treatment during steeping and germination of African finger millet grains at 30°C induced higher α-amylase activity compared to control (grains steeped in autoclaved distilled water). Samples of malted African finger millet grains treated at pH 9.0 during 96 h at 30°C showed higher α-amylase activity. Malted grains containing high α-amylase activity is important for the hydrolysis of starch and production of maltose, maltotriose and dextrin (Kanensi et al., 2013). Previous study showed that the maximum development of α-amylase activity usually occurs after 96 h in germinating millet (Mbithi-Mwikya et al., 2000). This is of great value because the proper development of the enzyme ultimately leads to complete sacchari?cation and high extract yield (Ukwuru, 2010). Heat stress of 40°C appears not to be appropriate compared to 30°C from the result of α-amylase activity. 30°C could be a more suitable temperature for the production of high levels of reducing sugars. This is in agreement with earlier report (Agu and Palmer, 1997).
The β-amylase activity in the malted millet is not well expressed compared to α-amylase. This is not unusual. Similar result was obtained from malted sorghum (Uriyo and Eigel, 1999). They posited that low β-amylase activity is attributed to the interaction of β-amylase with polyphenols to form insoluble polyphenol enzyme complexes. β-amylase activities is the best expressed at 30°C as against those of 40°C. The malted African finger millet grains treated at pH 3 and pH 9.0 showed lower β-amylase activity (125 and 130 U/g), compared to α-amylase activity (143 and 180 U/g), respectively, after 96 h incubation at 30ºC. Dreier et al. (1995) reported that β-amylase activity is known to increase in response to heat stress. In contrast, it was observed that the activity of β-amylase in the millet species malted was rather low as the temperature increased from 30 to 40°C. As suggested by Bwanganga et al. (2012), this may be due to the synthesis rate of the enzyme which is easily influenced by the germination temperature. (Taylor and Robbins, 1993) reported that a low temperature is optimal to obtain high β-amylase activities. This requirement of a relatively low germination temperature may be related to the fact that the β-amylase is thermolabile. It is glaringly evident that α-amylase and β-amylase activities work in tandem in the malted millet grains during the heat and pH stresses. Both α-amylase and β-amylase, have a joint amylolytic function in that α-amylase catalyses the hydrolysis of internal α-1, 4-O-glycosidic bonds in polysaccharides to produce soluble oligosaccharides with retained α-anomeric configuration which are in turn hydrolysed by β-amylase, liberating maltose; this joint amylolytic activity is known as the diastatic power (Usha et al., 2011).
The significance of the results obtained in the present study is that the finger millet grains that were subjected to alkaline pH stress at 30°C have a correspondingly high diastatic activity when compared to that of the other groups due to their respective α-amylase activity levels. Since the α-amylase activity is a good indicator of the diastatic power in the malted millets. In line with these observations, reports have it that diastatic activity increase is caused by an increase in activity of the amylase enzymes developed during germination (McCleary and Shameer, 1987). Another significance of the high α-amylase activity recorded in the present study is that the different fractions of the oligosaccharides obtained from the starch hydrolysis could also be separated and their functionality could be tested and patented in the food industry. Some infant products contain them in the hope that they might provide some of the benefits attributed to oligosaccharides in human milk (Rivero and Santamaria, 2001).
There is no significant difference in the endo-(1,3)(1,4)-β-D-glucanase enzyme activities in the finger millet grains subjected to alkaline and acidic pH stress and germinating at 30°C, especially those sampled between 0 to 24 h of germination. Figure 3 illustrates that malted millet grains obtained from steeping process in autoclaved distilled water (control) at 30ºC showed highest β-glucanase activity, while at the same temperature the enzyme appeared to be stable under the acid stress conditions at the 48th and 72nd h of germination. From their work, Gupta et al. (2010), observed that better malting performance is expected to be associated with lower levels of β-glucan in grains and higher levels of β-glucanase in malt. Recent findings also show that the degradation of endosperm cell walls and subsequent changes in β-glucan levels during malting is to a great extent, related to β-glucanase activity, which depolymerizes β-glucan (Gupta et al., 2010). Such levels of β-glucanase activity as observed among millet grains steeped under alkaline and acid pH stress conditions and germinated at 30°C (Figure 3) with a corresponding decrease in β-glucan concentration (Figure 4) suggests the possibility of the malted products being of good quality and may be a useful alternative to the malted products produced from other cereals or under other conditions. The quality (in terms of its viscosity) of the malt produced in the present study could be predicted from the results obtained on the β-glucan content of the malted finger millet grains (Figure 4). This is due to the fact that β-glucan molecules have the ability to self-associate and form aggregates, hence contributing to increased viscosity (Cui and Wood, 2000). Moreover, the starch content and alpha-amylase and beta-amylase activity may also influence the viscosity of the malted grains since poor degradation of β-glucans and poor hydrolysis of starch can lead to slow wort filtration, reduced extraction efficiency and haze development of the finished brewed product (Correia et al., 2008).
Stress due to salinity generally might not be unconnected to osmotic stress and possibly ion-induced imbalance (Munns, 2002). Our results show the induction of the amylolytic enzymes under this present study and attain saturation above 100 mM NaCl. Our result is not unusual. Salt stress has been reported to stimulate induction of b-amylase protein accumulation and activity in maize, pearl millet and barley (Kaplan et al., 2006). Here, there appear to be an added effect of salinity to alkali stress. The synergistic comparative mechanism of salinity-alkaline pH stress still remains unclear. However it is plausible to suggest that ionic imbalance and pH homeostasis could stimulate the induction of amylolytic enzymes to enhance malting. Malting quality is related to the hordein (cereal storage protein) fractions (Echart-Almedia and Cavalli-Molina, 2001). There was no significant change in the expression of the predominantly hordein polypeptide A and B during the stress conditions applied. There were no significant changes in polypeptide fractions of hordein A and B after acid and alkaline treatment of African finger millet malted grain. Here, it becomes clearer that change in hordein fraction might not be a good variable to malting potential in African finger millet.
In summary, the findings from this study reveals that, steeping practices are critical for good malt production when considering finger millet as the grain source. Finger millet malt quality is significantly affected by pH and temperature. Malt quality within the range of parameters studied increases with steeping time (24-72 h), the most effective condition for producing malted millet flour could be under alkaline pH (pH 9.0) and at 30°C. Thus, finger millet if properly harnessed under optimized conditions could be a very promising replacement for barley malt.
The authors did not declare any conflict of interest.
REFERENCES
|
Agu RC, Palmer GH (1997). The effect of temperature on the modification of sorghum and barley during malting. Process Biochem. 32(6):501-507.
Crossref
|
|
|
|
Bradford MM (1976). A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principal of protein dye-binding. Anal. Biochem. 38: 248-252.
Crossref
|
|
|
|
|
Bwanganga TJ, Bera F, Thonart P (2012). Modelling the beta-amylase activity during red sorghum malting when Bacillus subtilis is used to control mould growth. J. Cereal Sci. 57(1):115-119.
Crossref
|
|
|
|
|
Charalampopoulos D, Wang R, Pandiella SS, Webb C (2002). Application of cereals and cereal components in functional foods: a review. Int. J. Food Microbiol. 79:131-141.
Crossref
|
|
|
|
|
Clancy JA, Han F, Ullrich SE (2003). Comparative mapping of β-amylase activity QTLs among three barley crosses. Crop Sci. 43: 1043–1052.
Crossref
|
|
|
|
|
Correia I, Nunes A, Barros AS, Delgadillo I (2008). Protein profile and malt activity during sorghum germination. J. Sci. Food Agric. 88:2598–2605.
Crossref
|
|
|
|
|
Cui W, Wood PJ (2000). Relationships between structural features, molecular weight and rheological properties of cereal β-D-glucan. Amsterdam,Elsevier In K Nishinari (Ed.), Hydrocolloids, Part 1:159-168.
|
|
|
|
|
Dreier W, Schnarrenberger C, Borner T (1995). Light and stress-dependent enhancement of amylolytic activities in white and green barley leaves: beta-amylases are stress-induced proteins. J. Plant Physiol. 145:342-348.
Crossref
|
|
|
|
|
Echart-Almedia C, Cavalli-Molina S (2001). Hordein polypeptide patterns in relation to malting quality in Brazillian barley varieties. Pesq. Agropec. Bras. 36(2):211-217.
Crossref
|
|
|
|
|
Gimbi DM, Kitabatake N (2002).Changes in alpha-and beta-amylase activities during seed germination of African finger millet. Int. J. Food Sci. Nutr. 53:481-488.
Crossref
|
|
|
|
|
Gorzolka K, Lissel M, Kesslerb N, Loch-Ahring C, Niehaus C (2012). Metabolite fingerprinting of barley whole seeds, endosperms, and embryos during industrial malting. J. Biotechnol. 159:177-187.
Crossref
|
|
|
|
|
Gupta M, Abu GN, Gallaghar E (2010). Barley for brewing: Characteristic changes during malting, brewing and applications of its by-products. Compr. Rev. Food Sci. Food Saf. 9:318-328.
Crossref
|
|
|
|
|
Hounhouigan DJ, Nout MJR, Nago CM, Houben JH, Rombouts FM (2007). Composition and microbiological and physical attributes of mawe, a fermented maize dough from Benin. Int. J. Food Sci. Technol. 28:513-517.
Crossref
|
|
|
|
|
Jamar C, Jardin P, Fauconnier M (2011). Cell wall polysaccharides hydrolysis of malting barley (Hordeum vulgare L.): a review. Biotechnol. Agron. Soc. Environ. 15(2):301-313.
|
|
|
|
|
Kanensi OJ, Ochola S, Gikonyo NK, Makokha A (2013). Effect of steeping and germination on the diastatic activity and sugar content in amaranth grains and viscosity of porridge. J. Agric. Food. Technol. 3(1):1-7.
|
|
|
|
|
Kaplan F, Sung D, Guy CL (2006). Roles of ß-amylase and starch breakdown during temperatures stress. Physiol. Plant. 126:120-128.
Crossref
|
|
|
|
|
Laemmli UK (1970). Cleavage of structure proteins assembly of the head of bacteriophage. T4. Nature 22:680-685.
Crossref
|
|
|
|
|
Macnicol PK, Jacobsen JV, Keys MM, Stuart IM (1993). Effects of heat and water stress on malt quality and grain parameters of schooner barley grown in cabinets. J. Cereal Sci. 18:61-68.
Crossref
|
|
|
|
|
Mark RS, Ronald WS, Allen DB (2013). Protein mobilization and malting-specific proteinase expression during barley germination. J. Cereal Sci. 58(2):324-332.
Crossref
|
|
|
|
|
Mbithi-Mwikya S, Van CJ, Yiru Y, Huyghebaert A (2000). Nutrient and antinutrient changes in finger millet (Eleusine coracana) during sprouting. Lebensm. Wiss. u-Technol. 33:9-14.
|
|
|
|
|
McCleary BV, Shameer I (1987). Assay of malt β-glucanase using azo barley glucan: an improved precipitant. J. Inst. Brew. 93:87-90.
Crossref
|
|
|
|
|
Munns R (2002). Comparative physiology of salt and water stress. Plant Cell Environ. 25:239–250.
Crossref
|
|
|
|
|
Narayanaswamy D, Kurian S, Daniel VA, Rajalakshmi D, Swaminathan M, Parpia HA (1971). Supplementary value of a low cost protein food based on a blend of wheat and soyabean flours to poor rice and ragi diets. Nutr. Rep. Int. 4:109-116.
|
|
|
|
|
Nout MJ, Davies R (1982). Malting characteristics of finger millet, sorghum and barley. J. Inst. Brew. 88:157-163.
Crossref
|
|
|
|
|
Nzelibe HC, Agbojo DA (2005).Malting characteristics of different varieties of ï¬nger millet (Eleusine coracana) compared with Fonio millet (Digitaria exilis). J. Sci. 7(2):183-187.
|
|
|
|
|
Riss P, Bang-Olsen K (1991). Germination profile – A new term in malting barley analysis. In: Proceedings 23rd European Brewery Convention. Lisabon: 100-108.
|
|
|
|
|
Rivero UM, Santamaria OA (2001). Oligosaccharides: application in infant food. Early Hum. Dev. 65:43-52.
Crossref
|
|
|
|
|
Taylor JRN, Robbins DJ (1993). Factors influencing beta-amylase activity in sorghum malt. J. Inst. Brew. 99:413-416.
Crossref
|
|
|
|
|
Ukwuru MU (2010). Effect of steeping conditions on the amylolytic development of some Nigerian improved sorghum cultivars. J. Food Sci. Technol. 47(1):61-66.
Crossref
|
|
|
|
|
Uriyo M, Eigel WE (1999). Duration of kilning treatment on α-amylase, β-amylase and endo-(1,3)(1,4)-β-D-glucanase activity of malted sorghum (Sorghum bicolor). Process Biochem. 35:433-436.
Crossref
|
|
|
|
|
Usha B, Krishna VG, Muni KD, Hemalatha KPJ (2011). Partial characterization of α-amylase from germinating little millets (Panicum Sumatrense). J. Phytol. 3(1):1-8.
|
|
|
|
|
Wang J, Zhang G, Chen J, Wu F (2004). The changes of β-glucan content and β -glucanase activity in barley before and after malting and their relationships to malt qualities. Food Chem. 86:223-228.
Crossref
|
|
|
|
|
Yasuyuki OI, Kitabatake N (2003).Chemical composition of an east African traditional beverage, togwa. J. Agric. Food Chem. 51:7024-7028.
Crossref
|
|