ABSTRACT
The selection of resistance is inevitable whenever chemotherapy is necessary for pathogen control. Notably, Plasmodium falciparum has developed multifaceted means to overcome the toxicity of nearly all antimalarial medicines. To bypass this challenge, not only should novel drugs be developed, but the resistance mechanisms to new and existing drugs need should be fully explored. Pyronaridine is a companion drug in Pyramax®, a blend of artesunate (ASN)-pyronaridine (PRD) which is the WHO prequalified alternative for malaria treatment in the African setting. However, half-life mismatch predisposes the PRD to swift emergence of resistance especially in high malaria transmission settings. However, there are no well-characterized PRD-resistant parasite lines. Previously, stable PRD- resistant P. berghei ANKA lines were selected by in vivo drug pressure and preliminary results showed cross-resistance with quinolines, therefore, hypothetically the activity of PRD and chloroquine or other quinolines may be comparable, hence, the resistance mechanisms may be parallel. Consequently, genetic polymorphisms and expression profiles of PbMDR-1 that could be associated with pyronaridine resistance were examined by PCR amplification, sequencing and transcript quantification by RT-qPCR. The transcripts level increased during resistance selection while translated PbMDR-1 sequence alignment of PRD-sensitive and PRD-resistant was the same, the expression may be linked to PRD resistance but not mutations.
Key words: Quinolines, malaria, Pyronaridine, Pyramax®, resistance, expression, MDR-1 gene.
Malaria is one of the most devastating infectious diseases faced by the humanity in the 21st century. Over the past two decades, numerous strains of Plasmodium falciparum have developed resistance to nearly all anti-malarial drugs presented for clinical treatment of malaria illness (Hanboonkunupakarn and White, 2015; Menard and Dondorp, 2017).This development has prompted a quest for new effective anti-malarial compounds with the least side effects (Tang et al., 2020). One such strategy for plummeting the malaria prevalence is the usage of twin- or triple-anti-malarial drug combinations, which is thought to protect each drug from the development of resistance and reduce the overall transmission of malaria (Dipanjan et al., 2017; Tse et al., 2019; Mishra et al., 2017). In the last 20 years, over 60 countries and territories have officially adopted artemisinin-based combination therapy (ACT) for the treatment of falciparum malaria (WHO, 2017). The artemisinin derivatives cause a rapid and effective reduction in parasite biomass as well as gametocyte carriage, while the partner drug, which has a longer duration of action, achieves effective clinical and parasitological cure. Despite evaluation of different forms of ACT (Henrich et al., 2014; Mishra et al., 2017), clinical failures or at least longer parasite clearance times have been described in western Thailand, southern Myanmar, and possibly in the Vietnam and Cambodia (Kyaw et al., 2013; Hien et al., 2012). This emergence of parasite resistance to some forms of ACT indicates that novel compounds and combinations must be discovered and developed (Ouji et al., 2018).
To overcome drug resistance challenge, several forms of Artemisinin Combinational Therapies (ACTs) have been evaluated and now the currently WHO prequalified combination therapy, PRD/ASN (Ashley and Phyo, 2018; Tse et al., 2019; Henrich et al., 2014), which deployment is under a pharmacovigilance system in countries that may consider the drug in their national treatment guidelines (WHO, 2020). Pyronaridine (PRD) is the bis-mannich base, an analog of amodiaquine and like lumefantrine, it has been found to act through the inhibition of β-haematin formation although the mechanisms remain unclear (Chang et al., 1992; Croft et al., 2012). Recently, attention has been renewed in pyronaridine as a likely partner for use in artemisinin-based combination therapy (ACT) for malaria treatment (Croft et al., 2012; Tse et al., 2019). Indeed, PRD is highly effective against CQ-sensitive and CQ resistant parasites (Gupta et al., 2002; Vivas et al., 2008). Although PRD has not been commercially available as monotherapy or extensively deployed outside China, the in vitro sensitivity to this drug decreased in China between 1988 and 1995, suggested the emergence of PRD resistance (Croft et al., 2012). Pyronaridine resistance has previously been selected in Plasmodium berghei and Plasmodium yoelii (Croft et al., 2012; Kimani et al., 2014; Peters and Robinson, 1992); however, the molecular organization of the phenotype was not investigated. Pyramax® (PRD/ASN), remains active against widespread Chloroquine (CQ) and Sufadoxine-Pyrimethamine (SP) drug-resistant P. falciparum clearance of malaria infection, studies indicate that use of drug combination with mismatched pharmacokinetics does not prevent selection of the resistance against long-acting drug (Hastings and Hodel, 2014; Li and Hickman, 2015). Artesunate is a short-acting artemisinin derivative with half-life of less than 2 h, while PRD is long-acting with half- life of 16 to 17days (Park and Pradeep, 2010).
Indeed, PRD which is left trailing in sub-therapeutic doses within the body provides strong selection pressure for fast resistance emergence. Thus the need to understand the mechanism of PRD resistance towards the elucidation of molecular surveillance and health policy tools before or during drug use.
Single and multiple mutations, elevated transcript profiles and increased copy numbers of the P. falciparum multidrug resistance gene 1 (Pf PbMDR-1) have been linked with most antimalarial drug resistance in P. falciparum (Kiboi et al., 2014; Pradines et al., 2010; Tang et al., 2020). Therefore, to evaluate the role of point mutation and expression profiles in antimalarial pyronaridine resistance, we used previously generated stable PRD- resistant P. berghei ANKA lines by in vivo drug pressure (Kimani et al., 2014). Documented studies hypothesize that PRD mode of action may be comparable to that of CQ or other quinolines drug; consequently, they may share similar modes of action and resistance mechanisms (Dorn et al., 1998; Hanboonkunupakarn and White, 2015). As a result, the goal of this study is to identify genetic polymorphisms and expression profiles that could be associated with PRD resistance in selected genes associated with quinoline or any other anti-malarial drugs- preliminarily the multi-drug resistant gene 1 (MDR-1). This study is aware that the selected gene may not certainly be accompanying PRD resistance, thus proposes employment of whole-genome shotgun (WGS) sequencing approach and whole transcriptome profiling (WTP) of the dilution cloned resistant parasite using next sequencing generation sequencer (Illumina) to identify novel genes and copy number variation that may have accumulated during the drug selection process.
Parasites and experimental animals
The drug sensitive parasite lines of P. berghei ANKA acronymed as PRDS was used as reference line, while PRDR is as previously submitted to drug selection pressure and cloned as described in detail (Kimani et al., 2014). Male Swiss albino mice weighing 20-22 g out-bred at KEMRI, Animal house Nairobi, utilized for this study. The animals warehoused in the animal house in hard plastic cages, standard polypropylene, and placed on commercial rodent feeds and water ad libitum. Antimalarial PRD drug was newly made by dissolving it in a solvent consisting of 70% Tween-80 (d = 1.08 g/ml) and 30% ethanol (d = 0.81 g/ml) and successively diluted ten-fold with double distilled water. All experiment in this study involving use of animals was conducted in accordance with KEMRI guidelines, as well as, internationally accepted principles for laboratory animal use and care.
Dilution cloning of PRD resistant parasite
To generate genetically homogenous PRDR parasites, four different generations from pyronaridine resistant lines (Table 1) were dilution cloned as detailed (Janse et al., 2004). Briefly, a mouse with
parasitemia between 0.3 and 1% was selected as a donor mouse. Then, 5 μl of infected blood (parasitized red blood cells, RBCs) was collected from the tail of the mouse using μl of heparinized syringe and diluted in 1 ml of 1 × PBS buffer. The number of infected erythrocytes per 1 μl was estimated from 20 μl of diluted blood. The cell suspension was then diluted further with 1 × PBS buffer to an estimated final concentration of 0.5 parasites/200μl PBS. Fifteen mice were intravenously injected each with 200μl /mouse of parasitized RBCs. Cloning was considered effective when about fifth to half of the inoculated mice tested positive and bared a parasitemia of between 0.3 and 1% at day 8 post infection.
Determination of indices of PRD resistance
The stability of PRD resistant line was evaluated by: (i) measuring drug responses after making five drug-free passages and (ii) freeze-thawing of parasites from -80°C stored for a period of four weeks followed by the measurement of effective doses in the 4-Day suppressive test. Stable resistance was defined as the maintenance of the resistance phenotype when drug-selection pressure was removed for at least five passages in mice (Kimani et al., 2014). Acquisition of resistance was assessed after every five drug passages using standard 4-DT to confirm the response levels of the parasite to the pyronaridine compound. 4-DT permits the measurement of the ED50 and ED90, as well as the index of resistance at the 90% levels (I90). The indices of resistance (I90) were defined as the ratio of the ED90 of the resistant line to that of the parent strain as described by Kimani et al. (2014). Resistance was classified into three categories based on an earlier work (Merkli and Richle, 1980): I90 = 1.0, sensitive, (2) I90 = 1.01-10.0, slight resistance, (3) I90 = 10.01-100.0, moderate resistance and (4) I90> 100.0, high resistance.
Pyronaridine sensitivity profiles tests
To assess the resistance profile of individual clones generated by dilution cloning, the fastest growing clone among the four generation was selected and evaluated for its response to PRD in the 4-DT (Fidock et al., 2004). Briefly for each selected clone, mice were infected intraperitoneally with 1 × 106 parasites/mouse. Oral treatment of drug was initiated on day 0 (4 h post infection) and continued for 24, 48 and 72 h post infection. Parasite density was estimated microscopically (×100) after 96 h post parasite inoculation using thin blood films made from tail blood. Parasite growth was then followed for at least 15 days post-infection to assess the recrudescence of the parasites after cessation of drug treatment as earlier described elsewhere (Kiboi et al., 2014).
Parasite preparation and extraction of parasitic DNA
Parasitized blood cells were harvested from mice under general anaesthesia, when trophozoite stages were most prevalent into PBS (PH 7.2). In preparing parasitic DNA, mouse white blood cells was removed by successive filtration of infected blood using Plasmodipur filters (Euro-Diagnostica) as previously described (Janse et al., 2004). Intact parasites were released from their host red blood cells (RBCs) using RBC lysis buffer (Roche). Briefly, packed cells was re-suspended in 5 volumes of cold (4°C) erythrocyte lysis buffer for 5 min until the suspension becomes a clear red colour before spinning at 2000rmp for 8 min to pellet the parasite. Total genomic DNA of both the sensitive and resistant parasites was extracted using commercially available QiAamp DNA Blood kit (Qiagen) according to manufacturer’s instructions.
Amplification and sequencing of PbMDR-1 gene
A 4260-nucleotide length fragment of theMDR-1 gene was amplified by Polymerase Chain Reaction (PCR). Briefly, MDR-1 gene was amplified by PCR and sequenced using the primer pairs described by Kiboi et al. (2014) and also provided in Supplementary Table S1. The reaction mixture consisted of approximately 200 ng of genomic DNA, 0.5 μM of forward and reverse primers, 1X PCR buffer (Promega), 2 mM MgCl2, 200 μM deoxynucleotide triphosphate (dNTP) and 0.025 U DreamTaq polymerase (Eurogentec) in a final volume of 25µ μl. The thermo-cycler (Applied Biosystems)) was programmed as follows: an initial 95°C for 5 min followed by 30 cycles of 95°C for 1 minute, 48°C for 30 s and 68°C for 5 min. A final 10-min extension step was done at 72°C. A MDR-1 gene sequence was obtained from http://plasmodb.org. Accession No. PBANKA_123780. Primers for amplification and sequencing were manually designed (Table 1). Products of PCR amplification were analysed using gel electrophoresis system. After PCR products were purified by GeneJETTM PCR purification kit, sequencing was conducted using ABI Prism Big Dye Terminator v3.1 (Applied Biosystems, CA, and U.S.A) cycle sequencing ready reaction kits according to the manufacturer’s instructions.
Evaluation of gene expression of PbMDR-1gene
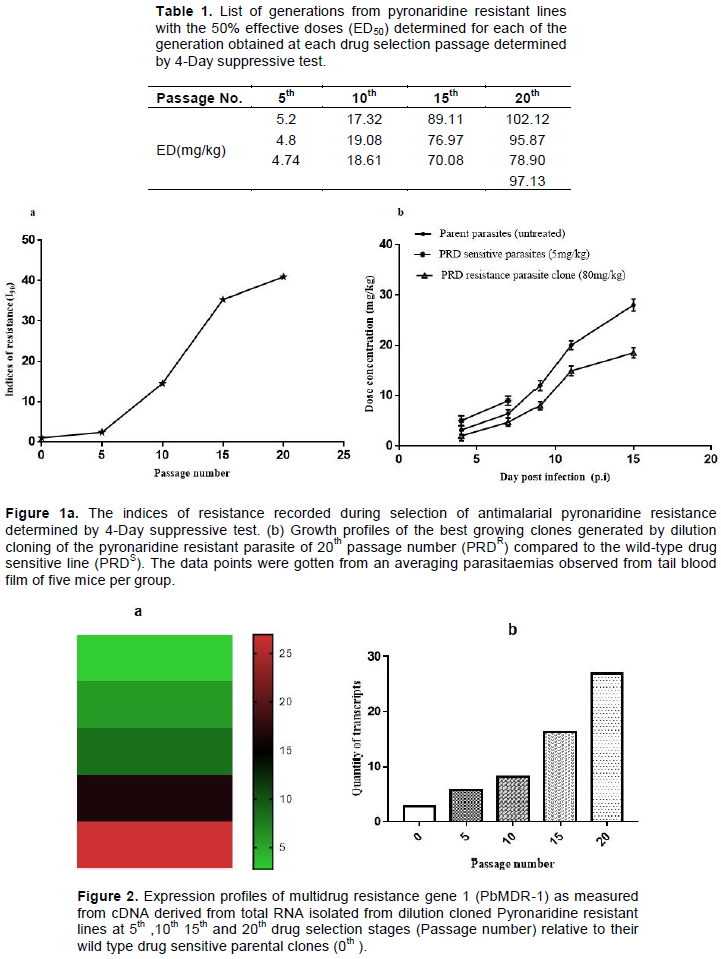
To determine the expression profile of the PbMDR-1 at specific selection stage presented as 5th, 10th, 15th and 20th drug pressure passage, clones were generated by dilution cloning and the best growing clone among the generations were selected as described earlier (Janse et al., 2004). To assess the mRNA transcripts quantity of PMDR-1gene, briefly fresh parasites were centrifuged and the pellet prepared. The total RNA was extracted from at least 1 × 106 parasites based on High Pure RNA extraction kit (Roche™). The RNA was immediately used for cDNA synthesis following the Transcriptor First Strand cDNA synthesis kit (Roche™).Briefly, the Transcriptor Reverse Transcriptase (20 U/μl) were added to first strand cDNA and RT reaction mix was incubated at 50°C for 60 min, then at 85°C for 5 min and finally chilled on ice. The cDNA was used as template for RT-PCR assays or stored at −15 to −20°C for longer period. Primers (Supplementary Table S2) were designed to run the reactions for PbMDR-1 in the same plate by using similar cycling conditions. Relative quantification of transcripts was performed using the housekeeping gene Pbβ-actin (PBANKA_145930) as a reference gene. The difference in relative expression levels of PbMDR-1 was calculated from 2−ΔΔCt value after normalization of data to actin (Livak and Schmittgen, 2001). All analyses were performed triplicate. The conditions for these reactions were: 94°C for 4 min; followed by denaturation at 94°C for 15 s and annealing/extension at 56°C for 30 s, for 40 cycles.
Data analysis
Gene sequences were manually compiled, then compared between drug selected and unselected clones (Altschul et al., 1990) and then analysed using MEGAX software (Kumar et al., 2018). The relative expression level data was normalized using Pbβ-actin as endogenous control gene using the formula 2-ΔΔCT (Livak and Schmittgen, 2001).
Previously, stable PRD- resistant P. berghei ANKA lines were selected by in vivo drug pressure (Figure 1a). The fastest growing clone of PRD resistant line (PRDR) was selected and subjected to drug sensitivity profiles (Table 1, Figure 1b). Subsequently, the best growing clone with high resistance profile was considered for the evaluation of genetic polymorphism and quantification of transcripts. Then the MDR-1 coding sequence of both PRD-Sensitive and PRDR clones were amplified and sequenced (Supplementary Figure S1). To explore the potential modulatory and compensatory role of MDR-1, the mRNA transcript level was measured. Consequently, these results demonstrated that the transcripts level increased during selection of pyronaridine resistance (Figure 2), while the translated PbMDR-1sequence alignment of PRD-sensitive and PRD-resistant was 100% identical (Supplementary Figure S2). Consistence with part of in vivo results, Pradine et al. reported the absence of association between pyronaridine in vitro responses and polymorphisms in genes involved in quinoline resistance in Plasmodium falciparum (Pradines et al., 2010).
The global dissemination of drug-resistant P. falciparum is spurring intense efforts to implement artemisinin (ART)–based combination therapies for malaria, including ASN/PRD. Nevertheless, resistance to at least one component of some forms of ACT currently in clinical use has been documented, and it is feared that ACT will gradually lose its clinical efficacy due to widespread use. Individual P. falciparum parasites with longer clearance times have been described in Cambodia (Rogers et al., 2009).
The biggest challenge at present is genetic elasticity and high frequency of human malaria parasite P. falciparum to develop resistance to structurally and mechanistically related and unrelated drugs (accelerated resistance to multiple drugs, ARMD) (Rathod et al., 1997), ARMD is characterized by the ability of a strain to generate a drug-resistant clone when put under drug pressure. This results from the high mutation rate during parasite multiplication. The goal of the present study was to investigate the role of multidrug resistance gene -1 which is known to be associated with reduced quinoline susceptibility in modulating PRD resistance. Previously, PRD resistance line was developed for over a period of six months and cloned the parasite which was highly growing and with the highest resistance; however, the mechanisms underlying this resistance was never investigated (Kimani et al., 2014).

It has been proposed that the selective pressure for resistance to PRD-ASN combinations would be exerted by the longer-acting PRD component. The mechanism by which resistance to pyronaridine develops is unknown, but may be due to a direct effect on the pyronaridine mechanism of action or modulation of targeted gene. For instance, Wu et al. (1988) described an increase in the number of food vacuoles in trophozoites from a pyronaridine-resistant P. berghei (RP) line, some of which were fusing. There was also a marked reduction in the digestive food vesicles containing malaria pigment granules for both trophozoites and schizonts and typical hemozoin grains were not formed in the pyronaridine-resistant parasites (Auparakkitanon et al., 2006; Wu et al., 1988). These and other ultra-structural differences suggested that resistance may be due to a direct effect on the pyronaridine mechanism of action. Another study by Li et al., found over-expression of a 54 kDa protein in a pyronaridine-resistant strain of P. berghei (ANKA) (Li et al., 1995). The protein was localized mainly in the cytoplasm of erythrocytic stage trophozoites, schizonts and merozoites and less commonly in the cytoplasm of infected erythrocytes(Li et al., 1995). Interestingly, a 54 kDa protein is also expressed in chloroquine-resistant P. berghei (ANKA) suggesting a common effect, though whether this is related to resistance development remains unknown (Li et al., 1995). Notably, Qi et al. proposed that antimalarial pyronaridine could be an inhibitor of P-glycoprotein mediated multidrug resistance in tumor cells (Qi et al., 2002; Qi et al., 2004).
The P. falciparum multidrug (MDR) resistance protein (PfMDR-1) has been implicated in altering parasite susceptibility to a variety of currently available antimalarial drugs. Point mutations in PfMDR-1 have been associated with changes in parasite susceptibility to Chloroquine, Quinine, Mefloquine, and Artemesinin derivatives in both laboratory lines and clinical isolates; however, these mutations have limited use as molecular markers (Duraisingh and Cowman, 2005; Woodrow and Krishna, 2006).
Comparison of coding region of parent strain and pyronaridine resistant line revealed that the nucleotide and translated protein sequence of the PbMDR-1 was identical. Thus, point mutation in MDR-1 was not involved in the generation of pyronaridine resistance. However, the expression levels of the gene increased in tandem with the increase of the resistance implying that the expression profile may be linked to PRD resistance.
The authors have not declared any conflict of interests.
The research was supported by the Government of Kenya through National Commission of Science, Technology and Innovation award to SKK grant NCST/ST&I/RCD/4th Call M.Sc./196.
REFERENCES
|
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990). Basic Local Alignment Search Tool. Journal of Molecular Biology 215:403-10.
Crossref
|
|
|
|
Ashley Elizabeth A, Phyo PA (2018). Drugs in Development for Malaria. Drugs 78(9):861-879.
Crossref
|
|
|
|
|
Auparakkitanon S, Chapoomram S, Kuaha K, Chirachariyavej T, Wilairat P(2006). Targeting of Hematin by the Antimalarial Pyronaridine.Antimicrobial Agents and Chemotherapy 50(6):2197-2200.
Crossref
|
|
|
|
|
Chang C, Lin-Hua T, Jantanavivat C (1992). Studies on a New Antimalarial Compound: Pyronaridine. Transactions of the Royal Society of Tropical Medicine and Hygiene 86(1):7-10.
Crossref
|
|
|
|
|
Croft SL, Duparc S, Arbe-barnes SJ, Carl Craft J, Shin C, Fleckenstein L, Borghini-fuhrer I, Rim H (2012). Review of Pyronaridine Anti-Malarial Properties and Product Characteristics. Malaria Journal 11(4):270.
Crossref
|
|
|
|
|
Dipanjan B, Shivaprakash G, Balaji O (2017). Triple Combination Therapy and Drug Cycling-Tangential Strategies for Countering Artemisinin Resistance. Current Infectious Disease Reports 19(7):1-7.
Crossref
|
|
|
|
|
Dorn A, Rani SV, Matile H, Jaquet C, Vennerstrom JL, Ridley RG (1998). An Assessment of Drug-Haematin Binding as a Mechanism for Inhibition of Haematin Polymerisation by Quinoline Antimalarials. Biochemical Pharmacology 55(97):727-736.
Crossref
|
|
|
|
|
Duraisingh MT, Cowman AF (2005). Contribution of the Pfmdr1 Gene to Antimalarial Drug-Resistance. Acta Tropica 94:181-190.
Crossref
|
|
|
|
|
Fidock DA, Rosenthal PJ, Croft SL, Brun R, Nwaka S (2004). Antimalarial Drug Discovery: Efficacy Models for Compound Screening. Nature Reviews Drug Discovery 3(6):509-520.
Crossref
|
|
|
|
|
Gupta S, Thapar MM, Mariga ST, Wernsdorfer WH,Björkman A (2002). Plasmodium falciparum: In Vitro Interactions of Artemisinin with Amodiaquine, Pyronaridine, and Chloroquine. Experimental Parasitology 100(1):28-35.
Crossref
|
|
|
|
|
Hanboonkunupakarn B, White NJ (2015). The Threat of Antimalarial Drug Resistance. Tropical Diseases, Travel Medicine and Vaccines 2(1):1-5.
Crossref
|
|
|
|
|
Hastings IM, Hodel EM (2014). Pharmacological Considerations in the Design of Anti-Malarial Drug Combination Therapies - Is Matching Half-Lives Enough? Malaria Journal 13 (1):1-15.
Crossref
|
|
|
|
|
Henrich PP, Brien CO, Sáenz FE, Cremers S, Kyle DE, Fidock DA (2014). Evidence for Pyronaridine as a Highly Effective Partner Drug for Treatment of Artemisinin-Resistant Malaria in a Rodent Model. Antimicrobial Agents and Chemotherapy 58(1):183-95.
Crossref
|
|
|
|
|
Hien TT, Thuy-Nhien NT, Phu HN, Boni MF, Thanh NV, Nha-Ca NT, Thai L (2012). In Vivo Susceptibility of Plasmodium falciparum to Artesunate in Binh Phuoc Province, Vietnam. Malaria Journal 11:1-11.
Crossref
|
|
|
|
|
Janse C, Ramesar J, Waters A (2004). Plasmodium Berghei: General Parasitological Methods.Leiden University Medical Center, The Netherlands pp. 26-27.
|
|
|
|
|
Kiboi, DM, Irungu B, Orwa J, Kamau L, Ochola-oyier LI, Ngángá J, Nzila A (2014). Experimental Parasitology Piperaquine and Lumefantrine Resistance in Plasmodium berghei ANKA Associated with Increased Expression of Ca 2 + / H + Antiporter and Glutathione Associated Enzymes. Experimental Parasitology 147:23-32.
Crossref
|
|
|
|
|
Kimani SK, Ng JK, Kariuki DW, Kinyua J, Kimani FT, Kiboi DM (2014). Plasmodium berghei ANKA : Selection of Pyronaridine Resistance in Mouse Model. African Journal of Biochemistry Research 8(6):111-17.
Crossref
|
|
|
|
|
Kumar S, Stecher G, Li M, Knyaz C, Tamura K (2018). MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Molecular Biology and Evolution 35(6):1547-1549.
Crossref
|
|
|
|
|
Kyaw MP, Nyunt MH, Chit K, Aye MM, Aye KH, Aye M, Lindegardh N (2013). Reduced Susceptibility of Plasmodium falciparum to Artesunate in Southern Myanmar. PLoS ONE 8(3).
Crossref
|
|
|
|
|
Li GD, Liu SQ, Ye XY, Qu FY (1995). Detection of 54-KDa Protein Overexpressed by Chloroquine-Resistant Plasmodium berghei ANKA Strain in Pyronaridine-Resistant P. berghei ANKA Strain] (in Chinese). Pharmacologica Sinica 16(1):17-20.
|
|
|
|
|
Li Q, Hickman M (2015). The Impact of Pharmacokinetic Mismatched Antimalarial Drug Combinations on the Emergence and Spread of Drug Resistant Parasites. Basic Pharmacokinetic Concepts and Some Clinical Applications 1:1-32.
Crossref
|
|
|
|
|
Livak KJ, Schmittgen TD(2001). Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2-ΔΔCT Method. Methods 25(4):402-408.
Crossref
|
|
|
|
|
Menard D, Dondorp A (2017). Antimalarial Drug Resistance: A Threat to Malaria Elimination. Cold Spring Harb Perspect Medicine.
Crossref
|
|
|
|
|
Merkli B, Richle RW (1980). Studies on the Resistance to Single and Combined Antimalarials in the Plasmodium berghei Mouse Model. Acta Tropica 37:228-231.
|
|
|
|
|
Mishra M, Mishra VK, Kashaw V, Iyer AK, Kashaw SK (2017). Comprehensive Review on Various Strategies for Antimalarial Drug Discovery. European Journal of Medicinal Chemistry 125:1300-1320.
Crossref
|
|
|
|
|
Ouji M, Augereau JM, Paloque L, Benoit-Vical F (2018). Plasmodium falciparum Resistance to Artemisinin-Based Combination Therapies: A Sword of Damocles in the Path toward Malaria Elimination. Parasite 25:24-30.
Crossref
|
|
|
|
|
Park SH, Pradeep K (2010). Absorption, Distribution, Excretion, and Pharmacokinetics of C 14 -Pyronaridine Tetraphosphate in Male and Female Sprague-Dawley Rats. Journal of Biomedicine and Biotechnology 59:1-9.
Crossref
|
|
|
|
|
Peters W, Robinson BL (1992). The Chemotherapy of Rodent Malaria. XLVII. Studies on Pyronaridine and Other Mannich Base Antimalarials. Annals of Tropical Medicine and Parasitology 86(5):455-465.
Crossref
|
|
|
|
|
Pradines B, Briolant S, Henry M, Oeuvray C, Baret E, Amalvict R, Didillon E, Rogier C (2010). Absence of Association between Pyronaridine in Vitro Responses and Polymorphisms in Genes Involved in Quinoline Resistance in Plasmodium falciparum. Malaria Journal 9:339.
Crossref
|
|
|
|
|
Qi J, Yang CZ, Wang CY, Wang SB, Yang M, Wang JH (2002). Function and Mechanism of Pyronaridine: A New Inhibitor of P-Glycoprotein-Mediated Multidrug Resistance. Acta Pharmaceutica Sinica 23:544-550.
|
|
|
|
|
Qi J, Wang S, Liu G, Peng H, Wang J, Zhu Z, Yang C (2004). Pyronaridine, a Novel Modulator of P-Glycoprotein-Mediated Multidrug Resistance in Tumor Cells in Vitro and in Vivo. Biochemical and Biophysical Research Communications 319(4):1124-1131.
Crossref
|
|
|
|
|
Ramharter M, Kurth F, Schreier AC, Nemeth J, Von Glasenapp I, Schlie M, Kammer J (2008). Fixed-Dose Pyronaridine-Artesunate Combination for Treatment of Uncomplicated Falciparum Malaria in Pediatric Patients in Gabon. Journal of Infectious Diseases 198:911-19.
Crossref
|
|
|
|
|
Rathod P, Mcerlean T, Lee P (1997). Variations in Frequencies of Drug Resistance in Plasmodium falciparum. Proceedings of the National Academy of Sciences 94:9389-9393.
Crossref
|
|
|
|
|
Rogers WO, Sem R, Tero T, Chim P, Lim P, Muth S, Socheat D, Ariey F, Wongsrichanalai C(2009). Failure of Artesunate-Mefloquine Combination Therapy for Uncomplicated Plasmodium falciparum Malaria in Southern. Malaria Journal 9:1-9.
Crossref
|
|
|
|
|
Tang Y, Ye Q, Huang H, Zheng W (2020). An Overview of Available Antimalarials: Discovery, Mode of Action and Drug Resistance Current Molecular Medicine 20(8):583-592.
Crossref
|
|
|
|
|
Tse EG, Korsik M, Todd MH (2019). The Past, Present and Future of Anti-Malarial Medicines. Malaria Journal 18(1):1-21.
Crossref
|
|
|
|
|
Vivas L, Rattray L, Stewart L, Bongard E, Robinson BL, Peters W, Croft SL (2008). Anti-Malarial Efficacy of Pyronaridine and Artesunate in Combination in Vitro and in Vivo. Acta Tropica 105(3):222-228.
Crossref
|
|
|
|
|
World Health Organization (WHO) (2017). World malaria report 2017.
Crossref
|
|
|
|
|
World Health Organization (WHO) (2020). The use artesunate-pyronaridine for the treatmnt of uncomplicated malaria.
View
|
|
|
|
|
Woodrow CJ, Krishna S (2006). Antimalarial Drugs : Recent Advances in Molecular Determinants of Resistance and Their Clinical Significance. Cell Molecular Life Science 63:1586-1596.
Crossref
|
|
|
|
|
Wu LJ, Rabbege JR, Nagasawa H, Jacobs G, Aikawa M (1988). Morphological Effects of Pyronaridine on Malarial Parasites. American Journal of Tropical Medicine and Hygiene 9:87-89.
|
|