Full Length Research Paper

ABSTRACT
Seawater changing chemistry has consequences on coastal ecosystems and their living resources. Future projections suggest the pH could drop ~0.2-0.3 pH units by the year 2100 under a business-as-usual (BAU) CO2 emission scenario. Marine calcifying organisms such as corals, calcifying algae, crustaceans, mussels, oysters and clams are most likely to be impacted by ocean acidification. The Isognomon alatus (flat tree oyster) is an important species that can be negatively affected by the lowering of seawater pH. Isognomon alatus is an important food source, a substrate for other benthic organisms (e.g., stone crab, Menippe mercenaria) and contribute to nutrients recycling in coastal ecosystems. The study was conducted to test the impacts acidified seawater CO2 on the growth of I. alatus under controlled laboratory conditions as well as field experiment. The Isognomon alatus lost weight and experienced negative growth rates of –0.56 ± 0.36 mg g-1day-1 under average pH values of 7.8 expected by the end of this century compared to a loss of –0.26 ± 0.23 mg g-1day-1 under ambient pH (value 8.1) conditions. In contrast, I. alatus incubated in a field experiment showed a gain in weight and positive growth of 3.30 ± 0.23 mg g-1day-1 despite exposure to pH levels (~7.4) during low tide significantly lower than those experienced in the laboratory. Overall, the results showed concern on the impacts of acidification flat tree oyster (Bivalvia:Isognomonidae). A decline of calcifying bivalves populations can impact coastal ecosystems function and indirectly affect the human beings that depend on them as a food source.
Key words: Ocean acidification, climate change, Isognomonidae, shell dissolution, bivalve's growth, estuarine.
INTRODUCTION
MATERIALS AND METHODS



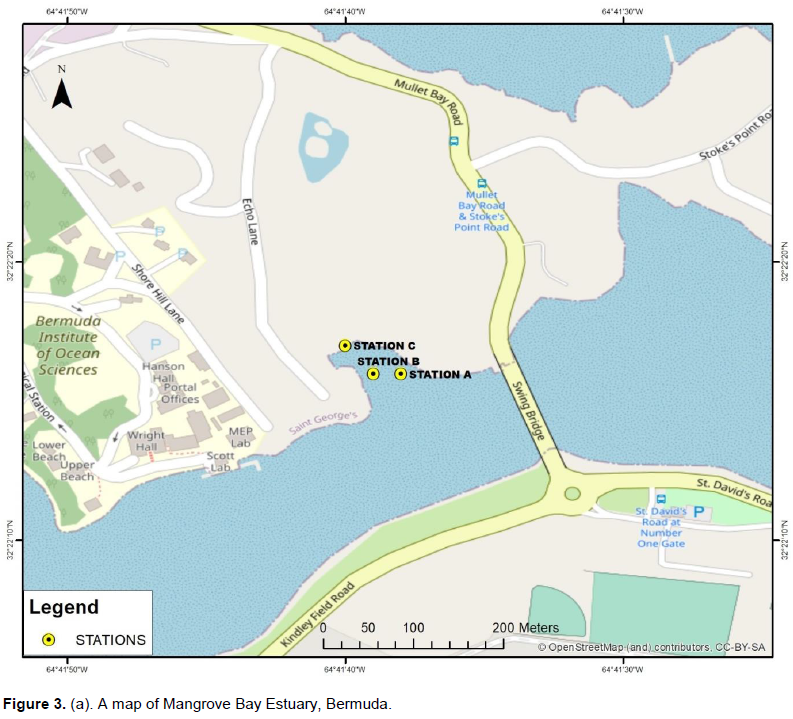



RESULTS














DISCUSSION
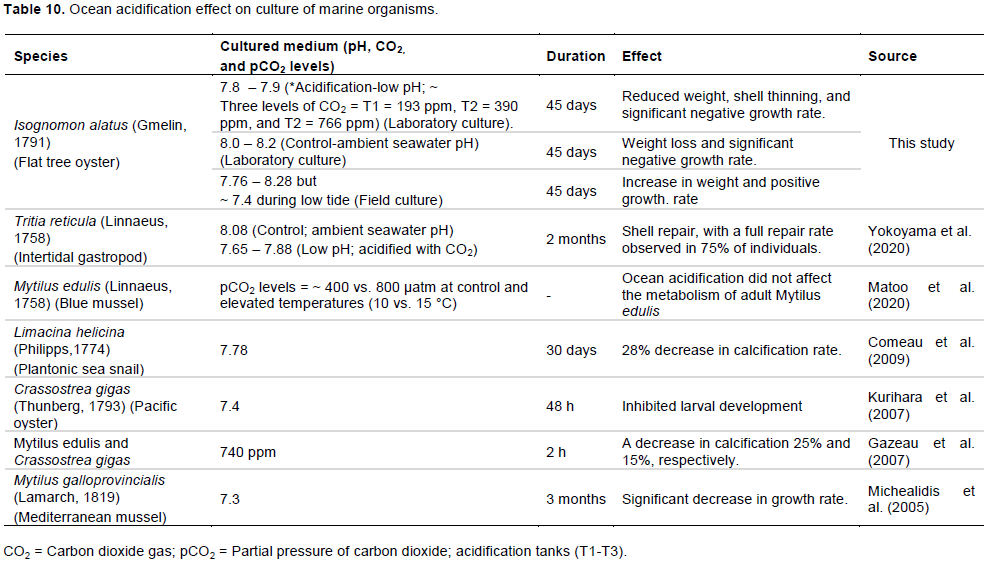
CONCLUSION

CONFLICT OF INTERESTS
ACKNOWLEDGMENTS
REFERENCES
|
Abbott RT, Morris PA (1995). A Field Guide to Shells: Atlantic and Gulf Coasts and the West Indies. Houghton Mifflin, New York. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Andersson AJ, Gledhill D (2013). Ocean acidification and coral reefs: effects on breakdown, dissolution, and net ecosystem calcification. Annual Review of Marine Science 5:321-348. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Andersson AJ, Mackenzie FT, Bates NR (2008). Life on the margin: Implications of ocean acidification on Mg-calcite, high latitude and cold-water marine calcifiers. Marine Ecology Progress Series pp. 265-273. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Baillie CJ, Grabowski JH (2019). Factors affecting recruitment, growth and survival of the eastern oyster Crassostrea virginica across an intertidal elevation gradient in southern New England. Marine Ecology Progress Series 609:119-132. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Baldry K, Saderne V, McCorkle DC, Churchill JH, Agusti S, Duarte CM (2020). Anomalies in the carbonate system of Red Sea coastal habitats. Biogeosciences 17:423-439. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Bamber RN (1987). The effects of acidic seawater on young carpet-shell clams Venerupis decussata (L.)(Mollusca: Veneracea). Journal of Experimental Marine Biology and Ecology 108:241-260. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Bamber RN (1990). The effects of acidic seawater on three species of lamellibranch mollusc. Journal of Experimental Marine Biology and Ecology 143:181-191. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Bates NR, Astor YM, Church MJ, Currie K, Dore JE, González-Dávila M, Lorenzoni L, Muller-Karger F, Olafsson J, Santana-Casiano JM (2014). A time-series view of changing ocean chemistry due to ocean uptake of anthropogenic CO2 and ocean acidification. Oceanography 27:126-141. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Bates NR, Best MHP, Neely K, Garley R, Dickson AG, Johnson RJ (2012). Detecting anthropogenic carbon dioxide uptake and ocean acidification in the North Atlantic Ocean. Biogeosciences 9:2509-2522. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Bates NR, Michaels AF, Knap AH (1996). Alkalinity changes in the Sargasso Sea: geochemical evidence of calcification?. Marine Chemistry 51:347-358. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Berge JA, Bjerkeng B, Pettersen O, Schanning MT, Oxnevad S (2006) Effects of increased seawater concentrations of CO2 on growth of the bivalve Mytilus edulis L. . Chemosphere 62:681-687. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Bibby R, Cleall-Harding P, Rundle SD, Widdicombe S, Spicer JI (2007). Ocean acidification induced defenses in the intertidal gastropod Littorina littorea. Biology Letters 3:699-701. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Bibby R, Widdicombe S, Parry H, Spicer JI, Pipe R (2008). Impact of ocean acidification on the immune response of the blue mussel Mytilus edulis. Aquatic Biology 2:67-74. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Bijma J, Portner HO, Yesson C, Rogers AD (2013). Climate change and the oceans--what does the future hold? Marine Pollution Bulletin 74:495-505. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Bindoff NL, Cheung WWL, Kairo JG, Arístegui J, Guinder VA, Hallberg R, Hilmi N, Jiao N, Karim MS, Levin L, O'Donoghue S, Purca Cuicapusa SR, Rinkevich B, Suga T, Tagliabue A, Williamson P (2019). Changing Ocean, Ecosystems, and Dependent Communities. In: Pörtner H-O, Roberts DC, Masson-Delmotte V, Zhai P, Tignor M, Poloczanska E, Mintenbeck K, Alegría A, Nicolai M, Okem A, Petzold J, Rama B, Weyer NM (eds) IPCC Special Report on the Ocean and Cryosphere in a Changing Climate pp. 477-587. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Bradshaw AL, Brewer PG (1988). High precision measurements of alkalinity and total carbon dioxide in seawater by potentiometric titration-1. Presence of unknown protolyte(s)?. Marine Chemistry 23:69-86. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Braga AC, Pereira V, Marçal R, Marques A, Guilherme S, Costa PR, Pacheco M (2020). DNA damage and oxidative stress responses of mussels Mytilus galloprovincialis to paralytic shellfish toxins under warming and acidification conditions - elucidation on the organ-specificity. Aquatic Toxicology 228:105619. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Brierley AS, Kingsford MJ (2009). Impacts of climate change review on marine organisms and ecosystems. Current Biology 19:R602-R614. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Brown J (1988). Multivariate analyses of the role of environmental factors in seasonal and site-related growth variation in the Pacific oyster Crassostrea gigas. Marine Ecological Progress Series 45:225-236. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Brown JR (1986). The Influence of Environmental Factors upon the Growth and Survival of the Pacific Oyster Crassostrea Gigas Thunberg. In: Department of Biological Sciences. Simon Fraser University. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Byrne M (2011). Impact of ocean warming and ocean acidification on marine invertebrate life history stages: Vulnerabilities and potential for presistence in a changing ocean. In: Gibson RN, Atkinson RJA, Gordon JDM (eds) Oceanography and Marine Biology: An Annual Review pp. 1-42. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Byrne M, Przeslawski R (2013). Multistressor impacts of warming and acidification of the ocean on marine invertebrates' life histories. Integrative and Comparative Biology 53:582-596. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Byrne M, Soars NA, Ho MA, Wong E, McElroy D, Selvakumaraswamy P, Dworjanyn SA, Davis AR (2010). Fertilization in a suite of coastal marine invertebrates from SE Australia is robust to near-future ocean warming and acidification. Marine Biology 157:2061-2069. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Caldeira K, Wickett ME (2003). Anthropogenic carbon and ocean pH. Nature 425(6956):365-365. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Caldeira K, Wickett ME (2005). Ocean model predictions of chemistry changes from carbon dioxide emissions to the atmosphere and ocean. Journal of Geophysical Research 110(C9):C09S04-1. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Campbell MD, Hall SG (2019). Hydrodynamic effects on oyster aquaculture systems: a review. Reviews in Aquaculture 11:896-906. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Campoy AN, Cruz JE, Ramos JB, Viveiros F, Range P, Teodósio MA (2020). Ocean acidification impacts on zooplankton. In: Teodosio MA, Branco Barbosa AM (eds) Zooplankton Ecology. CRC Press. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Cole JJ (2013). Chapter 6 The carbon cycle: With a brief introduction to global biogeochemistry. Fundamentals of Ecosystem Science. Academic Press, Oxford, UK, 109135. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Comeau S, Gorsky G, Jeffree R, Teyssié JL, Gattuso JP (2009). Key Arctic pelagic mollusc (Limacina helicina) threatened by ocean acidification. Biogeosciences Discussions 6:2523-2537. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Convey P, Peck LS (2019). Antarctic environmental change and biological responses. Science Advances Review 11:1-16. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Costa GB, Ramlov F, Koerich G, Navarro BB, Cabral D, Rodrigues ERO, Ramos B, Fadigas SD, Rörig LR, Maraschin M, Horta PA (2019). The effects of mining tailings in the physiology of benthic algae: understanding the relationship between mud's inductive acidification and the heavy metal's toxicity. Environmental and Experimental Botany 167:103818. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Curry A (2020). Effects of Multiple Stressors on the Development and Performance of Decapod Crustaceans. In: School of Ocean Sciences. Bangor University, Bangor, Wales, United Kingdom. P. 202. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
de Moel H, Ganssen GM, Peters FJC, Jung SJA, Brummer GJA, Kroon D, Zeebe RE (2009). Planktontic foraminiferal shell thinning in the Arabian Sea due to anthropogenic ocean acidification.?Biogeosciences Discussion 6:1811-1835. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Dekshenieks MM, Hofmann EE, Powell EN (1993). Environmental effects on the growth and development of eastern oyster, Crassostrea virginica (Gmelin, 1791), larvae: A modeling study. Journal of Shellfish Research 12:241-254. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Diaz R, Lardies MA, Tapia FJ, Tarifeño E, Vargas CA (2018). Transgenerational effects of pCO2-driven ocean acidification on adult mussels Mytilus chilensis modulate physiological response to multiple stressors in Larvae. Frontiers in Physiology 1349:1-12. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Dickson AG (1981). An exact definition of total alkalinity and a procedure for the estimation of alkalinity and total inorganic carbon from titration data. . Deep-Sea Research 28:609-623. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Dickson AG, Afghan JD, Anderson GC (2003). Reference materials for oceanic CO2 analysis: a method for the certification of total alkalinity. Marine Chemistry 80:185-197. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Dickson AG, Goyet C (1994). Handbook of methods for the analysis of the various parameters of the carbon dioxide system in seawater. In, United States. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Dickson AG, Millero FJ (1987). A comparison of the equilibrium constants for the dissociation of carbonic acid in seawater media. Deep Sea Research Part A 1,733-731,743. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Dickson AG, Sabine CL, Christian JR (2007). Guide to best practices for ocean CO2 measurements, Sidney, British Columbia, North Pacific Marine Science Organization. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Dissanayake A (2014). Ocean Acidification and Warming Effects on Crustacea: Possible Future Scenarios. In: Goffredo S, Dubinsky Z (eds). The Mediterranean Sea:Its history and present challenges. Springer Dordrecht, The Netherlands pp. 363-372. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Dissanayake A, Ishimatsu A (2011). Synergistic effects of elevated CO2 and temperature on the metabolic scope and activity in a shallow-water coastal decapod (Metapenaeus joyneri; Crustacea: Penaeidae). ICES Journal of Marine Science 68:1147-1154. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Dissard D, Nehrke G, Reichart GJ, Bijma J (2009). Impact of seawater pCO2 changes on calcification and on Mg/Ca and Sr/Ca in benthic foraminifera calcite (Ammonia tepida): results from culturing experiments. Biogeosciences Discussions 6:3771-3802. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
DOE (1994). Handbook of methods for the analysis of the various parameters of the carbon dioxide system in seawater. In: Dickson AG, Goyet C (eds). ORNL/CDIAC-74, 2nd Edition. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Doney S, Fabry V, Feely R, Kleypas J (2009). Ocean acidification: The other CO2 problem. Annual Review of Marine Science 1:169-192. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Doney SC, Balch VJ, Fabry VJ, Freely RA (2015). Ocean acidification: A critical emerging problem of the ocean sciences. Oceanography 22:16-25. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Doney SC, Busch DS, Cooley SR, Kroeker KJ (2020). The impacts of ocean acidification on marine ecosystems and reliant human communities. Annual Review of Environment and Resources 45. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Dong Z-G, Chen Y-H, Ge H-X, Li X-Y, Wu H-L, Wang C-H, Hu Z, Wu Y-J, Fu G-H, Lu J-K, Che H (2018). Response of growth and development of the Pacific oyster (Crassostrea gigas) to thermal discharge from a nuclear power plant. BMC Ecology 18(1):1-11. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Dupont S, Thorndyke MC (2009). Impact of CO2-driven ocean acidification on invertebrates early life-history. Biogeosciences Discussion 6:3109-3131. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
e Ramos BJ, Muller M, Riebesell U (2010). Short-term response of the coccolithophore Emiliania huxleyi to abrupt changes in seawater carbon dioxide concentrations. Biogeosciences 7: 4739-4763. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Egilsdottir H, Spicer JI, Rundle SD (2009). The effect of CO2 acidified sea water and reduced salinity on aspects of the embryonic development of the amphipod Echinogammarus marinus (Leach) Marine Pollution Bulletin 58:1187-1191. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
El Haïkali B, Bensoussan N, Romano J-C, Bousquet V (2004). Estimation of photosynthesis and calcification rates of Corallina elongata Ellis and Solander, 1786, by measurements of dissolved oxygen, pH and total alkalinity. Scientia Marina 68:45-56. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Emanuel MP, Pillay D, van der Merwe M, Branch GM (2020). Interactive effects of pH and temperature on native and alien mussels from the west coast of South Africa. African Journal of Marine Science 42(1):1-12. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Emerson CE, Reinardy HC, Bates NR, Bodnar AG (2017). Ocean acidification impacts spine integrity but not regenerative capacity of spines and tube feet in adult sea urchins. Royal Society Open Science 4(5):170140. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Fabricius KE, De'ath G, Noonan S, Uthicke S (2014). Ecological effects of ocean acidification and habitat complexity on reef-associated macroinvertebrate communities. Proceedings of Biological Science 281(1775):20132479. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Fabry VJ, Seibel BA, Feely RA, Orr JC (2008). Impacts of ocean acidification on marine fauna and ecosystem processes. Ices Journal of Marine Science 65:414-432. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Feely RA, Doney SC, Cooley SR (2009). Ocean acidification: Present conditions and future changes in a high-CO2 world. Oceanography 22(4):36-47. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Findlay HS, Wood HL, Kendall MA, Spicer JI, Twitchett RJ, Widdicombe S (2011). Comparing the impact of high CO2 on calcium carbonate structures in different marine organisms. Marine Biology Research 7:565-575. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Gazeau F, Quiblier C, Jansen JM, Gattuso JP, Middelburg JJ, Heip CHR (2007). Impact of elevated CO2 on shellfish calcification. Geophyical Research Letters 34(7). |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Goyet C, Snover AK (1993) High-accuracy measurements of total dissolved inorganic carbon in the ocean:comparison of alternate detection methods. . Marine Chemistry 44:235-242. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Gran G (1952). Determination of the equivalence point in potentiometric titrations. Part II. Analyst 77:661-667. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Green MA, Aller RC, Aller JY (1993). Carbonate dissolution and temporal abundances of foraminifera in long-Island sound sediments. Limnology and Oceanography 38:331-345. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Gutiérrez JL, Jones CG, Strayer DL, Iribarne, O.O. I (2003). Molluscs as ecosystem engineers: the role of shell production in aquatic habitats. Oikos 101:79-90. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Havenhand JN, Schlegel P (2009). Near-future levels of ocean acidification do not affect sperm motility and fertilization kinetics in the oyster Crassostrea gigas. Biogeosciences Discussions 6:4573-4586. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Herler J, Dirnwöber M (2011). A simple technique for measuring buoyant weight increment of entire, transplanted coral colonies in the field. Journal of Experimental Marine Biology and Ecology 407:250-255. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Hofmann GE, Barry JP, Edmunds PJ, Gates RD, Hutchins DA, Klinger T, Sewell MA (2010). The effect of ocean acidification on calcifying organisms in marine ecosystems: an organism-to-ecosystem perspective. Annual Review of Ecology, Evolution, and Systematics 41:127-147. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Holman JD, Burnett KG, Burnett LE (2009). Effects of hypercapnic hypoxia on the clearance of Vibrio campbellii in the blue crab, Callinectes sapidus. . Biological Bulletin 206:188-196. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Huang W-J, Wang Y, Cai W-J (2012). Assessment of sample storage techniques for total alkalinity and dissolved inorganic carbon in seawater. Limnology and Oceanography: Methods 10:711-717. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Intergovernmental Panel on Climate Change (IPCC) (2007). Climate change 2007: Synthesis Report. A contribution of Working Groups I, II and III to The Third Assessment Report of the Intergovernmental Panel on Climate Change. In: Watson RT (ed). Cambridge Press. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Intergovernmental Panel on Climate Change (IPCC) (2013). Climate Change 2013. In: Stocker TF, Qin D, Plattner G-K, M. Tignor, Allen SK, Boschung J, Nauels A, Xia Y, Bex V, Midgley PM (eds). The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Intergovernmental Panel on Climate Change, Cambridge University Press, Cambridge, United Kingdom and New York, NY. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Intergovernmental Panel on Climate Change (IPCC) (2014). Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. In: Pachauri RK, Meyer LA (eds). Intergovernmental Panel on Climate Change Geneva, Switzearland. P 151. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Intergovernmental Panel on Climate Change (IPCC) (2018). Global Warming of 1.5°. An IPCC Special Report on the impacts of global warming of 1.5°C above pre-industrial levels and related global greenhouse gas emission pathways, in the context of strengthening the global response to the threat of climate change, sustainable development, and efforts to eradicate poverty. In: Masson-Delmotte. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
V, Zhai P, Pörtner H-O, Roberts D, Skea J, Shukla PR, Pirani A, Moufouma-Okia W, Péan C, Pidcock R, Connors S, Matthews JBR, Chen Y, Zhou X, Gomis MI, Lonnoy E, Maycock T, Tignor M, Waterfield T (eds). |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Intergovernmental Panel on Climate Change (IPCC) (2019). IPCC Special Report on the Ocean and Cryosphere in a Changing Climate In: Pörtner H-O, Roberts DC, Masson-Delmotte V, Zhai P, Tignor M, Poloczanska E, Mintenbeck K, Alegría A, Nicolai M, Okem A, Petzold J, Rama B, Weyer NM (eds). |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Ishimatsu A, Hayashi M, Kikkawa T (2008). Fishes in high-CO2, acidified oceans. Marine Ecology Progress Series 373:295-302. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Jaffe R, Leal I, Alvarado J, Gardinali PR, Sericano JL (1998). Baseline study on the levels of organic pollutants and heavy metals in bivalves from the Morrocoy National park, Venezuela. Marine Pollution Bulletin 36:925-929. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Johnson KM, Sieburth JM, Williams PJL, Brändström L (1987). Coulometric total carbon dioxide analysis for marine studies: Automation and calibration. Marine Chemistry 21:117-133. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Johnson KM, Wills KD, Butler DB, Johnson WK, Wong CS (1993). Coulometric total carbon dioxide analysis for marine studies: maximizing the performance of an automated gas extraction system and coulometric detector. Marine Chemistry 44:167-188. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Jokiel PL, Maragos JE, Franzisket L (1978). Coral growth: buoyant weight technique. In: Stoddart DR, Johannes RE (eds). Coral reefs: research methods. UNESCO, Paris, Italy pp. 529-541. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Jones JM, Sweet J, Brzezinski MA, McNair HM, Passow U (2016). Evaluating carbonate system algorithms in a nearshore system: Does total alkalinity matter? . PLoS ONE 11(11):11. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Kawahata H, Fujita K, Iguchi A, Inoue M, Iwasaki S, Kuroyanagi A, Maeda A, Manaka T, Moriya K, Takagi H, Toyofuku T, Yoshimura T, Suzuki A (2019). Perspective on the response of marine calcifiers to global warming and ocean acidification-Behavior of corals and foraminifera in a high CO2 world "hot house". Progress in Earth and Planetary Science 6(1):1-37. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Keeling CD (1993). Surface Ocean CO2. In: Heimann M (ed). Global Carbon Cycle. Kluwer Publishers pp. 22-29. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Kinnear PR, Gray CD (1999). SPSS for Windows Made Simple. Psychology Press Ltd, Hove, United Kingdom. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Kleypas JA, Feely RA, Fabry VJ, Langdon C, Sabine CL, Robbins LL (2006). Impacts of Ocean Acidification on Coral Reefs and Other Marine Calcifiers: A Guide for Future Research. In. NSF, NOAA, and the U.S. Geological Survey, St. Petersburg P 88. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Kurihara H, Asai T, Kato S, Ishimatsu A (2009). Effects of elevated pCO2 on early development in the mussel Mytilus galloprovincialis. Aquatic Biology 4(3):225-233. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Kurihara H, Kato S, Ishimatsu A (2007). Effects of increased seawater pCO2 on early development of the oyster Crassostrea gigas. Aquatic Biology 1(1):91-98. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Kurihara H, Shirayama Y (2004). Effects of increased atmospheric CO2 on sea urchin early development. Marine Ecology Progress Series 274:161-169. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
La Peyre ML, Geaghan J, Decossas G, La Peyre JF (2016). Analysis of environmental factors influencing salinity patterns, oyster growth, and mortality in Lower Breton Sound Estuary, Louisiana, using 20 years of data. Journal of Coastal Research 32(3):519-530. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lacoue-Labarthe T, Martin S, Oberhänsli F, Teyssie JL, Markich S, Jeffree R, Bustamante P (2009). Effects of increased pCO2 and temperature on trace element (Ag, Cd and Zn) bioaccumulation in the eggs of the common cuttlefish, Sepia officinalis Biogeosciences Discuss 6:4865-4894. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Langdon C, Atkinson MJ (2005). Effect of elevated pCO2 on photosynthesis and calcification of corals and interactions with seasonal change in temperature/irradiance and nutrient enrichment. . JournalJournal of Geophysical Research C: Oceans 110:1-16. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Leal A, A.V.Flores AAV, Collin R, Tremblaya R (2019). Drifting in the Caribbean: Hints from the intertidal bivalve Isognomon alatus. Estuarine, Coastal and Shelf Science 227:106333. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Linto N, Barnes J, Ramachandran R, Divia J, Ramachandran P, Upstill-God RC (2014). Carbon dioxide and methane emissions from Mangrove-Associated Waters of the Andaman Islands, Bay of Bengal. Estuaries and Coasts 37(2):381-398. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Liu Z, Zhang Y, Zhou Z, Zong Y, Zheng Y, Liu C, Kong N, Gao Q, Wang L, Song L (2020). Metabolomic and transcriptomic profiling reveals the alteration of energy metabolism in oyster larve during initial shell formation and under experimental ocean acidification. Scientific Reports 10(1):1-11.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


