ABSTRACT
A bacterial strain named DDK6 was isolated from diesel-contaminated soil from a petrol station in Al-Hofuf city, after enrichment on diesel oil. The strain DDK6 formed a reddish-pink colony with a 2 to 3 mm diameter after two days of incubation at 30°C. Cells were Gram-positive coccoid and formed no endospores. Phenotypic identification by the automated bacterial system, Vitek compact II, identified the DDK6 as Kocuria sp. at 95% probability level. The 16s rRNA gene sequencing analysis confirmed the identity of the strain as K. sedimins at an identity level of 99.15%. Results of Gas chromatography-mass spectrometry (GC-MS) revealed that the DDK6 degraded the C14-C19 compounds in diesel. In addition, the DDK6 strain consumed the majority (68%) of the carbon sources tested, including monosaccharides, disaccharides, polysaccharides, and sugar alcohols as noticed by biochemical characterization using the API 50CH. The cultural, biochemical, and molecular characteristics were in general agreement with the strain identification. The results confirmed the metabolic versatility of the strain DDK6, in addition to its ability to degrade diesel oil, thereby providing ecological and environmental merits for its application in bioremediation of hydrocarbon pollutants.
Key words: Bioremediation, hydrocarbon, Kocuria, diesel oil.
There is an increasing concern globally about potential environmental consequences arising from contamination by accidental petroleum release during storage, transport, or exportation. Saudi Arabia is one of the largest oil-producing countries worldwide, therefore marine and terrestrial biota are negatively affected. Diesel oil is a common fuel for diesel engines. Chemically, it is a mixture of aliphatic and aromatic hydrocarbons produced during petroleum separation by fractional distillation. Leakages of diesel oil can occur from storage tanks when the oil seeps into soils and groundwater causing severe environmental problems (Das and Chandran, 2010). The ecological effects of diesel on plants growing on diesel-contaminated soils result in reduction of seed germination, plant growth and productivity. Additionally, diesel sticks with soil particles causing bad soil aeration, leading to a decline in microbial biodiversity. Consequently, the ecosystem functioning is negatively affected and the biogeochemical cycling of essential elements become truncated (Ciric et al., 2010).
One promising and efficient strategy to remove diesel oil from soil and water is via microbial degradation. Bioremediation is an ecofriendly, cost-cheap, versatile and efficient treatment of hydrocarbons. It has been confirmed in many laboratories worldwide that several bacterial groups possess the catabolic machinery for diesel oil. Bacterial species, which belong to Acinetobacter, Bacillus, Citrobacter, Corynebacterium, Flavobacterium and Micrococcus are representatives of oil-degrading bacteria (Jirasripongpun, 2002; Das and Chandran, 2010). Furthermore, hydrocarbon-degrading bacteria that have recently been documented in different localities in Saudi Arabia, include Stenotrophomonas maltophilia (Arulazhagan et al., 2017), Cupriavidus taiwanensis, Ochrobactrum intermedium, Pseudomonas aeruginosa and P. citronellolis, (Oyehan and Al-Thukair 2017).
Kocuria is Gram-positive cocci arranged in pairs, short chains, tetrads, cubical packets of eight and irregular clusters. Kocuria belongs to the phylum Actinobacteria, class Actinobacteria, order Actinomycetales, sub order Micrococcinae and family Micrococcaceae. The genus Kocuria was coined by Stackebrandt et al. (1995) and separated from Micrococcus based on chemotaxonomic and phylogenetic features (Stackebrandt et al., 1995). Currently, there are more than 18 species of Kocuria identified based on the 16S rRNA phylogenetic studies. In general, Kocuria spp. are non-pathogenic however, some species have been isolated from infected superficial and deep human tissues. At the time of writing, 20 different species with validated names are included under the genus Kocuria. The completely detailed list of the species is outlined in http://www.bacterio.cict.fr/k/kocuria.html. Most of these bacterial species have been isolated from diverse ecological niches. For example, K. dechangensis obtained from saline and alkaline soils (Wang et al., 2015), K. salsicia from salt-fermented seafood (Yun et al., 2011), K. gwangalliensis from seawater (Seo et al., 2009), K. palustris and K. rhizophila from rhizoplane of Typha angustifolia (Kovács et al., 1999), K. pelophila rhizosphere of a mangrove (Hamada et al., 2016). Additionally, K. sedimins has been isolated from a sediment sample from Kerala, India and described by polyphasic approaches (Bala et al., 2012) but its ability to degrade diesel oil was not investigated. Researchers have showed the role of Kocuria spp. in bioremediation of hydrocarbon (Esmaeil et al., 2009), removal of copper from copper- contaminated soils (Achal et al., 2011) and production of probiotics (Sharifuzzaman et al., 2014),biocontrol agents (Sharifuzzaman and Austin 2010), plant-growth-promoting activities (Egamberdieva, 2008). Members of the genus Kocuria have been shown to produce, kocurin, a novel thiazolyl peptide antibiotic, which exhibited anti-bacterial activities against clinically relevant strains (Bacillus subtilis, methicillin-resistant Staphylococcus aureus (MRSA), Acinetobacter baumannii and Candida albicans) (Palomo et al., 2013).
Hydrocarbon-degrading bacteria that have recently been investigated in different localities in Saudi Arabia, include Stenotrophomonas maltophilia (Arulazhagan et al., 2017),Cupriavidus taiwanensis, Ochrobactrum intermedium, Pseudomonas aeruginosa and P. citronellolis, (Oyehan and Al-Thukair 2017). Jeddah, Khobar and Duhran. However, little is known about the biodegradability of diesel-oil in Al-Houf, eastern region, Saudi Arabia. In addition, the search for local diesel-oil-degrading bacteria from soils exposed to the prevailing conditions in Al-Houf city has a pivotal importance in bioremediation approaches. Therefore, the current study aimed at isolation and characterization of a bacterial strain, designated DDK6, which was able to grow on diesel as a sole energy and carbon sources. To fulfill this aim, a soil sample was collected from a diesel-contaminated soil from a petrol station.
Collection of soil sample
A diesel contaminated soil sample was collected a petrol station in Al-Hofuf Saudi Arabia, in a sterilized screw-capped test tube. In the laboratory, 0.5 g of soil sample was enriched with diesel oil (1%v/v) as a sole carbon source in a 200 ml conical flask containing 50 ml mineral salts (MS) medium with the composition; 1 g (NH4)2SO4, 0.8 g K2HPO4, 0.2 g KH2PO4, 0.2 g MgSO4·7H2O, 0.1 g CaCl2·2H2O, and 5 mg FeSO4·7H2O in 1 L distilled water, pH 7) (Wu et al., 2013). Flasks were placed in a shaking-incubator at 100 rpm for three weeks. To ensure oil degradation ability, subsequent transfer of inoculum (3%) into a fresh MS medium was carried out.
Isolation, purification and preservation of the strain
Aliquots (100 µl) from the diesel-enriched media were streaked into MS agar plates and sprayed with diesel on the surface. Plates were incubated at 30°C for 7 days. Single colonies were picked and sub-cultured into fresh MS agar amended with diesel.
Morphology of colony and cells
The morphological characteristics of the colony; pigmentation, diameter, elevation and transparency was determined visually of 24 h old colony growing on soya agar medium and incubated at 30°C. Cell shape, arrangement and reaction to the gram staining were assessed.
Biochemical characterization using the API50Ch strip kit
In order to investigate the biochemical characteristics of the strain DDK6 API50Ch strip kit (Biomerieux, France) was used following the guidelines of the manufacturer. Results were recorded after 48 h of incubation of API50Ch.
Identification of the strain using Vitek compact II
The bacterial strain DDK6 was identified using the automated system for identification of bacterial strains, Vitek compact II. Gram-Positive Card was used and the data analysis were carried out using the Software version: 05.02 (Biomerieux, Mary L’Etoile, France).
Identification of the strain DDK6 using 16S rRNA gene sequencing
PCR amplification of the 16S rRNA gene
16S rRNA gene was amplified using the universal primers; 27F 5'-AGAGTTTGATCMTGGCTCAG-3' and 1492R 5'-TACGGYTACCTTGTTACGACTT-3’ (Weisburg et al., 1991) in 20 µl total PCR reaction. Genomic DNA was extracted from the strain using the InstaGene Matrix (Bio-Rad, USA) following the instructions of the manufacturer. PCR conditions were adjusted as previously described (Khalifa et al., 2015).
16S rDNA sequencing and construction of phylogenetic tree
For sequencing of the 16S rRNA gene, the Big Dye terminator cycle sequencing kit (Applied BioSystems, USA) was used. Sequencing products were resolved on an Applied Biosystems model 3730XL automated DNA sequencing system (Applied BioSystems, USA).
Blast search and calculations of similarity values of pairwise nucleotide 16s rRNA gene sequence were performed using EzTaxon server (http://www.eztaxon.org/; Chun et al., 2007). Multiple alignments with sequences of the most closely related recognized Kocuria strains and construction of phylogenetic tree were carried out by using the Neighbor-joining method based on the Tamura-Nei model (Tamura and Nei, 1993). Evolutionary analyses were conducted in MEGA5.02 (Tamura et al., 2011). The 16s rRNA gene sequence of strain DDK6 was deposited in the NCBI database under the accession number KY307788.
Detection of diesel oil degradation using the Gas chromatography-mass spectrometry (GC-MC)
The ability of the strain DDK6 to degrade diesel oil was further detected using GC-MS. DDK6 was inoculated into 50 ml MS medium containing 1% (V/V) diesel oil and incubated at 30°C with shaking 150 rpm for 5 days. Non-inoculated flask containing the same growth medium was used as control. After incubation period, diesel oil was extracted using equal volume of dichloromethane and analyzed using a Shimadzu GCMS –QP2010 SE instrument. The flow rate of the Helium as a carrier gas was set at 6.0 ml/min. The temperatures of injector and detector were adjusted at 250 and 300°C, respectively. The temperature program was as followings: 2-min hold at 60°C, ramp to 300°C at 20°C/min and 6-min hold at 300°C.
K. sediminis strain DDK6 was isolated from diesel-contaminated soils from a petrol station in Al-Hofuf. The cultural characteristics are outlined in Table 1. DDK6 formed a reddish-pink, convex colony, with a diameter of 2 to 3 mm, and entire margin. Cells of the strain DDK6 were coccoid, formed no endospores and exhibited positive reaction to the Gram staining. These features were typical to the type strain of K. sediminis FCS-11T (Bala et al., 2012) confirming the identity of the DDK6 strain.
Biochemical characterization using the API50Ch strip kit
The biochemical characteristics of the strain DDK6 using the API50Ch (Biomerieux, France) were shown in Table 2. API 50 CH is a phenotypic-based system, comprising 50 biochemical tests designated to estimate the carbohydrate metabolism ability of a microorganism. The system is exploited in the current study as a fast, effective a reliable tool to study the metabolic versatility of the strain DDK6 towards different sources of carbohydrate.
DDK6 exhibited the ability to utilize a wide range of different carbon including monosaccharides (e.g., D-galactose and L-arabinose) disaccharides (e.g., D-maltose and D-lactose), polysaccharides (e.g., Starch and Glycogen) and sugar alcohols (e.g., D-galactose, D-mannitol). Out of 49, DDK6was able to utilize 33 (~67%) different carbon compounds tested. However, 33% of the carbon compounds were not metabolized by the strain under study. D-turanose, D-lyxose, D-Tagatose, D-Fucose, Arabitol, potassium gluconate and Inulin are representatives of carbon sources that were not consumed by the strain DDK6. The biochemical profile of the strain DDK6 is shown by the API 50CH kit strip is typical to that of the K. sediminis (Bala et al., 2012) providing another evidence to the reliable identification of the strain DDK6. Genome sequences of members of Kocuria genus such as K. marina SO9-6 (Castro et al., 2015) and K. rhizophila strain TPW45 (Tan et al., 2016) revealed the existence of many gene clusters involved in catabolic pathways of carbohydrate. The ability of the strain DDK6 to grow using a wide range of different biochemical compounds. This range of metabolic versatility could provide an explanation about the ubiquity of Kocuria sp. in diverse ecological niches particularly, in oil-contaminated ecosystems.


Identification of the strain using mass spectrometry technology
Vitek II is a powerful and accurate automated platform with an expanded identification database for rapid microbial identification, and antibiotic susceptibility testing based on biochemical analysis using colorimetry. The bacterial strain DDK6 was identified as Kocuria sp. (at 98%) by using the Vitek compact II indicating that this tool is efficient for bacterial identification at the genus and specific levels. The efficiency of this system to correctly identify Staphylococcus spp. and other bacterial genera has been proven previously (Chatzigeorgiou et al., 2011; Paim et al., 2014). Nonetheless, discordance between VitekII and 16S rRNA gene sequencing for bacterial identification has been reported. For example, VitekII identified a bacterial strain from human blood as K. kristinae (score of 98%) whereas 16s rRNA sequencing identified it as Rothia amarae (Abouseada et al., 2016).
16S rDNA sequencing and of phylogenetic analysis
The 16S rRNA gene sequencing is a well-established technique for a robust and accurate bacterial identification and for inferring phylogenetic relationships among species. Comparative analyses of the 16S rRNA gene sequencing revealed that the strain DDK6 was related to the genus Kocuria. Sequence analysis showed that DDK6 was most closely related to K. sediminis strain FCS-11T (99.15% identity), followed by K. flava HO-9041T (98.9%), K. turfanensis HO-9042T (98.9%), K. dechangensis NEAU-ST5-33T (98.41%), K. polaris CMS 76orT (98.28%), K. rosea DSM 20447T (98.27%), K. aegyptia YIM 70003T (98.28%), K. himachalensis K07-05 T (97.51%), K. atrinae P30 T (97.1%) at similarity levels of gene sequences. As can be seen in Figure 1, the neighbour-joining phylogenetic tree (Figure 1) clearly highlighted that the strain DDK6 grouped with the K. sediminis strain FCS-11. The 16S rRNA gene sequencing confirmed phenotypic identification of the strain DDK6 based on Vitek II. Similar results have been obtained by Hassan et al. (2016) who showed that both Vitek2 and 16S rRNA gene sequencing correctly identified K. kristinae (99%) (Hassan et al., 2016).

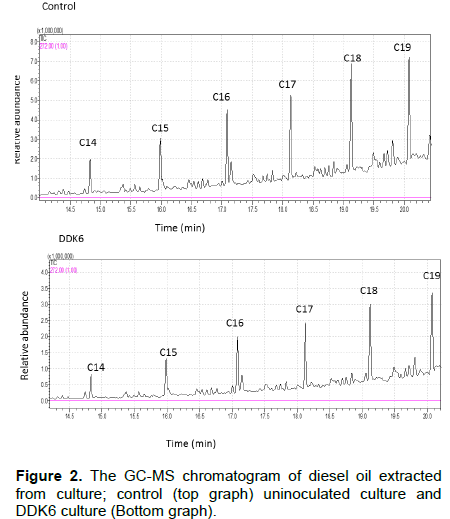
As can be seen in Figure 2, the major components of the diesel oil were C14-C19 alkanes. The peak areas of the C14-C19 were significantly lower than those in the control indicating that DDK6 was able to degrade this fraction of diesel oil (Figure 2). Our reseults are in accordance with those obtained by Mariano et al. (2007) who demonstrated that K. palustris was able to efficiently degrade diesel oil (Mariano et al., 2007). Microbial biodegradation of diesel oil and/or its components is a common process in terrestrial and aquatic ecosystems (Austin and Groves et al., 2011). Two main classes of metalloenzymes are involved in this multi-step process. The first class comprises membrane-associated enzymes such as alkane hydroxylase and the latter composed of cytoplasmic –soluble enzymes such as cytochrome P540. The existence of the alkane hydroxylase and cytochrome P540 in the strain DDK6 is consistent with previous studies on other bacterial species such as Acinetobacter sp. (Hou et al., 2013), Enterobacter cloaecea (Ramasamy et al., 2017), Pseudomonas aeruginosa and Bacillus subtilis (Safdari et al., 2017). Both enzymes exhibited wide range of alkane substrates and could have synergistic effect. The ability of other Kocurial species to grow on oil and other hydrocarbon as a sole carbon and energy sources has been documented. For example, K. flava and K. rosea were shown to degrade naphthalene, phenanthrene and fluoranthene and crude oil (Tumaikina et al., 2008). Apparently, the efficiency of local bacterial strains in hydrocarbon-degradability was found to be substantially higher than that of the introduced strains (Wu et al., 2013), this could be attributed to outcompeting, biotic and/or abiotic interacting factors. Therefore, isolation of new bacterial strains adapted to the local conditions of a particular area is crucial for the efficiency of hydrocarbon clean up in that area. It has been reported that production of biosurfactants enhances the bacterial oil degradation (Matvyeyeva et al., 2014). Biosurfactants are active molecules that lower the interfacial tension between two immiscible liquids. Sarafin et al. (2014) have highlighted that K. marina BS-15 produced biosurfactants reflecting its efficiency in oil degradation via increasing its water solubility to facilitate enzyme attack.

Generally, microbial degradation of diesel oil and other hydrocarbon is attributed to cytochrome P450, haem-thiolate monooxygenases, which catalyze the oxidative addition of atomic oxygen to the C-H or C-C bond of the organic compound (Van Beilen and Funhoff, 2007). Data mining for the Kocurial species that possess Cytochrome P450 showed that 17 different putative Cytochrome P450 proteins were found in 11 species (Table 3). Testing these species is crucial to confirm the ability to degrade diesel. Other enzymes such as soluble and particulate methane monoxygenases are involved in hydrocarbon degradation (Das and Chandran, 2011).

In conclusion, the cultural, biochemical and molecular characteristics were in general agreement for the strain DDK6 identification as K. sedimins. DDK6 exhibited a metabolic versatility and ability to degrade diesel oil indicating ecological and environmental merits for its application in bioremediation of hydrocarbon pollutants. Nonetheless, further studies are required to investigate the efficiency of the strain DDK6 for biodegradation of different organic pollutants and enzymes involved in this process at the molecular level, to explore the potentialities of the strain for biotechnological exploiting. Bearing in mind the efficiency of local bacterial strains in hydrocarbon-degradability was found to be substantially higher than that of the introduced strains (Wu et al., 2013), this could be attributed to outcompeting, biotic and/or abiotic interacting factors. This is the first report addresing K. sediminis as an oil-degrading bacterium isolated from an oil-contaminated soil exposed to the prevailing conditions in Al-Hofuf, Saudi Arabia.
The authors has not declared any conflict of interests.
REFERENCES
|
Abouseada N, Moheb M, Moez P (2016). Impact of MALDI-TOF in the Routine Diagnostic Microbiology Laboratory in Alexandria University, Egypt. Egypt. J. Med. Microbiol. 3:616-620.
|
|
|
|
Achal V, Pan X, Zhang D (2011). Remediation of copper-contaminated soil by Kocuria flava CR1, based on microbially induced calcite precipitation. Ecol. Eng.10:1601-1605.
Crossref
|
|
|
|
Arulazhagan P, Al-Shekri K, Huda Q, Godon JJ, Basahi JM, Jeyakumar D (2017). Biodegradation of polycyclic aromatic hydrocarbons by an acidophilic Stenotrophomonas maltophilia strain AJH1 isolated from a mineral mining site in Saudi Arabia. Extremophiles, 1:163-174.
Crossref
|
|
|
|
Austin RN, Groves JT (2011). Alkane-oxidizing metalloenzymes in the carbon cycle. Metallomics, 8:775-87.
Crossref
|
|
|
|
Bala M, Kaur C, Kaur I, Khan F, Mayilraj S (2012). Kocuria sediminis sp. nov, isolated from a marine sediment sample. Antonie Van Leeuwenhoek. 3:469-478.
Crossref
|
|
|
|
Castro DB, Pereira LB, e Silva MVM, da Silva BP, Palermo BRZ, Carlos C., ... and Gomes MB (2015). High-quality draft genome sequence of Kocuria marina SO9-6, an actinobacterium isolated from a copper mine. Genomics data, 5:34-35.
Crossref
|
|
|
|
Chatzigeorgiou KS, Sergentanis TN, Tsiodras S, Hamodrakas SJ, Bagos PG (2011). Phoenix 100 versus Vitek 2 in the identification of gram-positive and gram-negative bacteria: A comprehensive meta-analysis. J. Clin. Microbiol. 49:3284-3291.
Crossref
|
|
|
|
Chun J, Lee JH, Jung Y, Kim M, Kim S, Kim BK, Lim YW (2007). Extaxon: a web-based tool for the identification of prokaryotes based on 16S ribosomal RNA gene sequences. Int. J. Syst. Evol. Microbiol. 57(10):2259-2261.
Crossref
|
|
|
|
Ciric L, Philip JC, Whiteley AS (2010). Hydrocarbon utilization within a diesel-degrading bacterial consortium. FEMS Microbiol. Lett. 303:116-122.
|
|
|
|
Das N, Chandran, P (2010). Microbial degradation of petroleum hydrocarbon contaminants: an overview. Biotechnol. Res. Int. 2011:1-13
|
|
|
|
Esmaeil AS, Drobiova H, Obuekwe C (2009). Predominant culturable crude oil-degrading bacteria in the coast of Kuwait. Int. Biodeter. Biodegr. 4:400-406.
|
|
|
|
Egamberdieva D (2008). Plant growth promoting properties of rhizobacteria isolated from wheat and pea grown in loamy sand soil. Turk. J. Biol. 1: 9-15.
|
|
|
|
Hamada M, Shibata C, Tamura T, Nurkanto A, Ratnakomala S, Lisdiyanti P, and Suzuki KI (2016). Kocuria pelophila sp. nov, an actinobacterium isolated from the rhizosphere of a mangrove. Int. J. Syst. Evol. Microbiol. 9:3276-3280.
|
|
|
|
Hassan RM, Bassiouny DM, Matar Y (2016). Bacteremia Caused by Kocuria kristinae from Egypt: Are There More? A Case Report and Review of the Literature. Case Reports in Infectious Diseases. 31:1-4.
Crossref
|
|
|
|
Hou D, Shi Z, Shen X, He Y, Sun M, Wang Q (2013). Isolation, identification and alkane hydroxylase genes detection of a marine diesel-degrading bacterial strain (F9). Afr. J. Microbiol. Res. 22:2794-2802.
|
|
|
|
Jirasripongpun K (2002). The characterization of oil-degrading microorganisms from lubricating oil contaminated (scale) soil. Lett. Appl. Microbiol. 35:296-300.
Crossref
|
|
|
|
Khalifa AY, Almalki MA (2015). Isolation and characterization of an endophytic bacterium, Bacillus megaterium BMN1, associated with root-nodules of Medicago sativa L. growing in Al-Ahsaa region, Saudi Arabia. Ann. Microbiol. 2:1017-1026.
Crossref
|
|
|
|
Kovács G, Burghardt J, Pradella S, Schumann P, Stackebrandt E, Màrialigeti K (1999). Kocuria palustris sp. nov. and Kocuria rhizophila sp. nov, isolated from rhizoplane of the narrow-leaved cattail (Typha angustifolia). Int. J. Syst. Bacteriol. 49:167-173.
Crossref
|
|
|
|
Mariano AP, Kataoka AG, Angelis DF, BonottoDM (2007). Laboratory study on the bioremediation of diesel oil contaminated soil from a petrolstation, Braz. J. Microbiol. 2:346-352.
Crossref
|
|
|
|
Matvyeyeva OL, Vasylchenko OA, Aliieva OR (2014). Microbial biosurfactant role in oil products biodegradation. Int. J. Environ. Biorem. Biodegrade. 2:69-74.
|
|
|
|
Oyehan, TA, Al-Thukair AA (2017). Isolation and characterization of PAH-degrading bacteria from the Eastern Province, Saudi Arabia. Mar. Pollut. Bull. 2:39-46.
Crossref
|
|
|
|
Paim TGDS, Cantarelli VV, d'Azevedo PA (2014). Performance of the Vitek 2 system software version 5.03 in the bacterial identification and antimicrobial susceptibility test: evaluation study of clinical and reference strains of Gram-positive cocci. Rev. Soc. Bras. Med. Trop. 3:377-81.
Crossref
|
|
|
|
Palomo S, González I, de la Cruz M, Martín J, Tormo JR, Anderson M, Hill R, Vicente F, Reyes F, Genilloud O (2013). Sponge-Derived Kocuria and Micrococcus spp. as sources of the new thiazolyl peptide antibiotic Kocurin. Mar. Drugs. 4:1071-1086.
Crossref
|
|
|
|
Ramasamy S, Arumugam A, Chandran P (2017). Optimization of Enterobacter cloacae (KU923381) for diesel oil degradation using response surface methodology (RSM). J. Microbiol. 2:104-111.
Crossref
|
|
|
|
Safdari MS, Kariminia HR, Nejad ZG, Fletcher TH (2017). Study Potential of Indigenous Pseudomonas aeruginosa and Bacillus subtilis in Bioremediation of Diesel-Contaminated Water. Water Air Soil Pollution 1:37-41.
Crossref
|
|
|
|
Sarafin Y, Donio MBS, Velmurugan S, Michaelbabu M, Citarasu T (2014). Kocuria marina BS-15 a biosurfactant producing halophilic bacteria isolated from solar salt works in India. Saudi J. Biol. Sci. 6:511-519.
Crossref
|
|
|
|
Seo YB, Kim DE, Kim GD, Kim HW, Nam SW, Kim YT, Lee JH (2009). Kocuria gwangalliensis sp. nov, an actinobacterium isolated from seawater. Int. J. Syst. Evol. Microbiol. 59:2769-2772.
Crossref
|
|
|
|
Sharifuzzaman SM, Austin B (2010). Kocuria SM1 controls vibriosis in rainbow trout (Oncorhynchus mykiss, Walbaum). J. App. microbial. 6:2162-2170.
|
|
|
|
Sharifuzzaman SM, Al-Harbi AH, Austin B (2014). Characteristics of growth, digestive system functionality, and stress factors of rainbow trout fed probiotics Kocuria SM1 and Rhodococcus SM2. Aquaculture, 418:55-61.
Crossref
|
|
|
|
Stackebrandt E, Koch C, Gvozdiak O, Schumann P (1995). Taxonomic dissection of the genus Micrococcus: Kocuria gen. nov, Nesterenkonia gen. nov, Kytococcus gen. nov, Dermacoccus gen. nov, and Micrococcus Cohn 1872 gen. emend. Int. J. Syst. Bacteriol. 45:682-692.
Crossref
|
|
|
|
Tumaikina YA, Turkovskaya OV, Ignatov VV (2008). Degradation of hydrocarbons and their derivatives by a microbial association on the base of Canadian pondweed. Appl. Biochem. Microbiol. 45:382-388
Crossref
|
|
|
|
Tamura K, Nei M (1993). Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 10:512-526.
|
|
|
|
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S (2011). MEGA5, Molecular Evolutionary Genetics Analysis using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods Mol. Biol. Evol. 28:2731-2739.
Crossref
|
|
|
|
Tan-Guan-Sheng Adrian PW, Tan JWC, Wai-Fong Yin KGC (2016). Draft genome sequence of Kocuria rhizophila strain TPW45, an actinobacterium isolated from freshwater. J. Genomics, 4:16-18.
Crossref
|
|
|
|
Van Beilen JB, Funhoff EG (2007). Alkane hydroxylases involved in microbial alkane degradation. Appl. Microbiol. Biotechnol. 1:13-21.
Crossref
|
|
|
|
Wang K, Zhang L, Liu Y, Pan Y, Meng L, Xu T, Zhang C, Liu H, Hong S, Huang H, Jiang J (2015). Kocuria dechangensis sp. nov, an actinobacterium isolated from saline and alkaline soils. Int. J. Syst. Evol. Microbiol. 65:3024-3030.
Crossref
|
|
|
|
Weisburg WG, Barns SM, Pelletier DA, Lane DJ (1991). 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 17:697-703.
Crossref
|
|
|
|
Wu M, Chen L, Tian Y, Ding Y, Dick WA (2013). Degradation of polycyclic aromatic hydrocarbons by microbial consortia enriched from three soils using two different culture media. Environ. Pollut. 178:152-158.
Crossref
|
|
|
|
Yun JH, Roh SW, Jung MJ, Kim MS, Park EJ, Shin KS, Nam YD, Bae JW (2011). Kocuria salsicia sp. nov, isolated from salt-fermented seafood. Int. J. Syst. Evol. Microbiol. 61:86-289.
Crossref
|