Full Length Research Paper

ABSTRACT
This study was conducted to isolate and characterize soil microorganisms capable of solubilizing Chilembwe and Sinda rock phosphates readily available in Zambia. Single isolates were obtained by direct plating and enrichment cultures with succinate, cellulose and glucose as the carbon sources. Isolates were differentiated using colony morphology (form, margin, elevation and colour) and a subset of 20 was chosen for identification using the 16S rRNA gene sequences. After being cultured in nutrient broth and defined medium with tricalcium phosphate as the sole phosphate source, ten isolates were characterized for acid phosphatase enzyme activity. Over 80 isolates were obtained from different rock phosphate-sugar combinations. Isolates had varying morphological characteristics and the 20 strains identified showed a large diversity of organisms belonging to the genera Enterobacter, Burkholderia, Arthrobacter, Bacillus, Beijerinckia and two genera not previously reported to have P-solubilizing capabilities, Dyella and Curtobacterium. Both the observed growth and genera to which isolates belonged were dependent on the type of rock phosphate. Almost all isolates tested showed acid phosphatase activity that were similar to previously reported levels, ranging from 0.62 to 4.8 µg p-nitrophenol per 108 cells per hour. This work will be useful as the basis for the development of appropriate seed inocula that can enhance Sinda and Chilembwe rock phosphate dissolution and increased plant available P for crop production.
Key words: Chilembwe, Sinda, Rock Phosphate, Burkholderia, Enterobacter, Rhizobia, Dyella, Arthrobacter, Curtobacterium, Dyella.
INTRODUCTION
Phosphorus is second only to nitrogen as an inorganic nutrient required by both plants and microorganisms. Its major physiological role is in the storage and transfer of biochemically useful energy through the phosphate
anhydride bond (Plante, 2007). It is also found in different cellular components as a constituent of genetic material and phospholipids. Phosphorous is primarily rock derived and therefore, large amounts of P are found either in soil minerals or organic matter (Syer et al., 2008). There is very little P present in soil solution, with concentrations usually 0.1 to 1 µg/g at any given time (Plante, 2007), accounting for about 0.1% of the total soil P (Sharma et al., 2013).
The availability of native P in most Zambian agricultural soils is relatively low, especially in highly weathered and high P-sorption soils such as Alfisols, Ultisols and Oxisols (Yerokun, 2008). The limiting levels of available P necessitates the use of commercially produced fertilizers to supplement P for crop production. Alternatives to commercially available fertilizers include crop residues and animal manure; these sources cannot, however, on their own mitigate soil fertility decline because of limited quantities available on farms (Ademiluyi and Omotoso, 2007) and alternate uses of the residues as forage, silage and fuel. For example, the green biomass of Tithoniadiversifoloia has been recognized to be high in nutrients and effective as a nutrient source for lowland rice and maize (Ademiluyi and Omotoso, 2007). Particularly, T. diversifolia has been shown to have relatively high concentrations of N, P and K in its green biomass (Jama et al., 2000) and tissue concentrations of P can be greater than the critical 2.5 g kg-1 for net mineralization (Palm et al., 1999). Guano excreted by seabirds, seals or bats is another source of organic P that can be used as fertilizer. In Zambia, Guano enriched with P can contain as high as 4.03% P and contains beneficial fungi and bacteria that act as a natural control of plant diseases and that participate in organic matter decomposition (Sikazwe and de Waele, 2004).
Phosphorus can also be obtained from minerals; this mineral form of P is known as rock phosphate (RP), which consists of different forms of mineral apatite. Most of the world’s fertilizers are produced from RP resources (Van Straaten, 2007). There are several deposits of RP in sub-Sahara Africa, including Zambia which are found in two types of igneous rocks: carbonatitesand syenites (Van Straaten, 2002).
Plant roots associated with certain soil microorganisms can enhance the dissolution of these mineral forms of P by the production of organic acids such as acetic, citric, lactic, malic, oxalic, succinic, tartaric, glycolic, formic, and ketogluconic acids (Whalem and Sampedro, 2010). Organic acids dissolve Al- and Fe-P complexes and hasten the weathering of apatite-rich rock fertilizers as well as other primary minerals. Carbonic acid produced from the respiration of plant roots and soil microorganisms promotes the dissolution of calcium and magnesium P compounds (Plante, 2007). The production of other organic acids, such as gluconic acid, seems to be the most frequent means of mineral phosphate solubilisationbybacteria suchasErwiniaherbicola, Burkholderiacepacia, Rhizobium leguminosarium, Rhizobium meliloti, and Bacillus firmus (Brempong, 2013).In vitro assays have shown reduction in the pH of cultures by at least one unit due to the production of acid by phosphate solubilizing microorganisms (Kang et al., 2002). Microorganisms are also able to release inorganic P from minerals through other reactions such as the formation of chelating agents which are able to sequester aluminium, iron, calcium and magnesium ions from the mineral (Altomare et al., 1999). Several works have demonstrated that phosphate solubilising strains of Rhizobium, Bradyrhizobiumand Azotobacter increase the growth and phosphate content in both non-leguminous and leguminous plants (Khan et al., 2007).
Soil microorganisms also produce other compounds such as acid and alkaline phosphatase enzymes, which hydrolyse ester bonds of organic matter releasing P into solution for plant and microbial uptake (Nannipieri et al., 2011). While acid phosphatase is derived mainly from soil microbial communities, production by roots has also been observed (Dick et al., 1983; Juma and Tabatabai, 1988a, b, c).Recent studies have shown that some microorganisms that have the capability of producing organic acids involved in RP solubilisation also have enzyme activity through the production of acid phosphatase and pyrophosphatase enzymes (Traoré et al., 2013).
The activities of phosphatases, and organic acids and chelating compounds in making organic and mineral P sources available, respectively, depend on several factors such as soil properties and interactions among soil microorganisms. For example, the effect of soil type on the ability of the microorganisms to solubilise P has been shown in Ghana on rice fields (Brempong, 2013). This suggests the importance of selecting microor-ganisms appropriate for particular soil types when developing inoculants.
This study was conducted to isolate and characterize Chilembwe and Sinda RP solubilizing soil microor-ganisms and to determine if these organisms can make organic forms of P available through acid phosphatase activity. This work serves as the first step to the develop-ment of inoculants that would increase the agronomic effectiveness and value of Chilembwe and Sinda RP readily available in Zambia for crop production.
MATERIALS AND METHODS
Soil sampling, processing and characterization
Soils were collected from the Pinney Purdue Agricultural Center in Wanatah, in northwestern Indiana, USA (latitude 41.44278, longitude -86.92905). Six random soil samples (approximately 3 kilograms each; P1-P6) were collected from the periphery of a soybean field and immediately put on ice for transportation back to the laboratory. Soils were then processed by sieving and apportioned for different tests before storing at either 4°C or -10°C. About 250 g of soil previously stored at 4°C was prepared and sent for chemical analysis at the A & L Great Lakes Laboratories (Fort Wayne, IN). Soils were characterized for organic matter, phosphorous, potassium, magnesium, and calcium concentrations, pH, and cation exchange capacity using standard laboratory procedures.
Culture medium preparation
To isolate soil microorganisms that have the capability to solubilize P and that use different carbon sources, two rock phosphates readily available in Zambia were used: Sinda (S-RP) and Chilembwe (C-RP) (Figure 1). Minimal media (MM) were prepared according to the procedure of Traoré et al. (2013) with modifications to the carbon source and their amounts. Three carbon sources were used to enrich for growth of soil microorganisms: succinate (S), glucose (G) and cellulose (C). The media contained per litre of distilled water: carbon source (1.9 g glucose, 1.4 g succinate or 1.9 g cellulose), 5 g rock phosphate (Sinda or Chilembwe), 5 g MgCl2.6H2O, 0.25 g MgSO4.7H2O, 0.2 g KCl, and 0.1 g (NH4)2SO4. Plates for isolating rock phosphate dissolving microorganisms were made with the same medium with 10 g agar added. All media preparations were adjusted to pH 7 before autoclaving at 121°C and 15 psi for 30 min. With the three carbon sources and two rock phosphates, a total of six treatments were used and each replicated six times (according to the soil samples collected). No-soil and no rock-phosphate controls were also included.

Soil enrichments and direct plating
Culturing of soil microorganisms was done either as soil enrichments or as direct plating of soil dilutions. Direct plating was achieved using 100 µl of a 100-fold dilution of soil solution (in deionized water) that was spread evenly on a plate; plates were then sealed with parafilm and incubated upside down at 30°C until colonies developed. The soil enrichments were prepared by diluting soil a 100-fold into a total volume of 120 ml of MM solution. Flasks were then sealed with cotton plugs and incubated at 30°C in an environmental shaker (120 rpm) for a week before sub-culturing into fresh medium. At the time of sub-culturing, sub samples were obtained from the week old cultures and diluted serially to a million fold and 100 µl was plated onto corresponding agar plates made with corresponding carbon sources. Microorganism growth on agar plates and in enrichment cultures was then observed and recorded.
Isolation and characterization of single isolates
As previously mentioned, plating of sub samples was conducted at each subculture and growth was observed. Single colonies were obtained from the plates, streaked onto fresh individual plates and purified. Morphological characterization of isolates was done based on colony form, colour, elevation, and margin. Colonies that showed growth of hyphae were recorded as ‘fungal’ and the colour of the mycelia recorded.
For genetic characterization and identification of isolates, 60 isolates were selected initially.Selected isolates were cultured overnight in Difco nutrient broth (NB); cultures with OD600 readings of 1-1.5 were immediately used for DNA extraction. DNA was isolated from only 36 out of the 60 selected isolates due to poor growth of some of the isolates in the nutrient broth. DNA was extracted from the cells using the FastDNA SPIN Kit (Q-Biogene; Solon, OH). The isolated DNA was then assessed for quality using gel electrophoresis (0.8% agarose gel run for 10 minutes at 100 V) and the 260/280 nm OD and quantified using a NanoDrop 3300 fluorospectrophotometer (Thermo Scientific, Wilimington, DE).
A subset of (20) isolates was carried through for identification using the almost full length 16S rRNA gene (approximately 1500 bp) that was PCR amplified using bacteria specific primers (27F, 5’-AGA GTT TGA TCC TGG CTC AG--3’ and 1452R, 5’-AAG GAG GTG ATC CAG CCG CA—3). The gene was amplified in a 50 µl PCR reaction mix containing 1´ PCR buffer, 75 mmol MgCl2, 4 nmoldeoxynucleoside triphosphates, 100 pmol of forward and reverse primers, and 50-100 ng of template DNA. The thermal cycle protocol began with an initial denaturation at 95°C for 5 min, then thirty cycles of 94°C for 30 s, 55°C for 30 s, and 72°C for 30 s, followed by 10 min at 72°C. The PCR products were directly sequenced using the Sanger Sequencing method at the Purdue University Genomics facilities. Best matches to nucleotide sequences in the GenBank database (NCBI) were determined using the BLASTn program (Altschul et al., 1997). The best matches to bacterial type strains were determined using SeqMatch and the RDP database (Cole et al., 2014).
To illustrate the relatedness of the identified isolates to type strains of officially classified taxa, a phylogenetic tree was generated. Clustal Omega (Sievers et al., 2011) was used for multiple sequence alignment and the neighbour-joining method to build the tree using 1000 reiterations. Only isolates for which the full 16S rRNA gene sequence was obtained were used for the phylogenetic tree along with the best matching type strains.
Biochemical characterization using the acid phosphatase assay method
Selected isolates were grown in nutrient brothand also in MM with tricalcium phosphate (TCP) as the sole P source instead of rock phosphate for the acid phosphatase assay. The second medium was used to test enzyme activity in the absence of any organic phosphate source. Growth in both media was observed and recorded. Acid phosphatase activity for each isolate was determined by the method of Eivazi and Tabatabai (1977), however rather than using 1 g of soil, 1 mL of broth culture was used. In brief, one 1 mL of broth culture was mixed with 4 mL of pH 6.5 modified universal buffer (MUB) and 0.2 mL of toluene to inhibit cellular metabolism, then 1 µL of 0.05 M disodium p-nitrophenyl phosphate (PNP) tetrahydrate substrate solution was added. The mixture was incubated for 1 hour at 37°C. The reaction was stopped by adding 1 mL of 0.5 M CaCl2 and 4 mL of 0.5 M NaOH. The resulting solution was filtered using Whatman #2 filter paper, the solution’s yellow colour intensity was measured at 410 nm on a spectrophotometer and compared to a standard curve to determine the concentration of released substrate. Acid phosphatase activity was then normalized to µg of PNP released/108 cells. Cell density was estimated by measuring the optical density at 600 nm. This value was then converted to cells/mL using the E. coli cell density conversion of an OD600 of 1.0 being equivalent to 8 x 108 cells/mL.
RESULTS
Soil characteristics
Selected soil chemical characteristics are shown in Table 1. In order to set up subsequent isolation experiments, it was important that the selected soil samples were low in available phosphorous. The results indicated that only Pinney 1 had medium levels (28 mg kg-1) of P while the others had either low or very low P concentrations (Table 1). The pH ranged from 6.4 to 7.0 with an average of 6.6. The levels of potassium ranged from medium to high (103 to 200 mg kg-1) with an average of 128.7 mg kg-1. Magnesium levels were all very high and ranged from 465 to 575 mg kg-1. Calcium levels were low in one of the samples (Pinney 3) but the rest were in the medium range (1350 to 1800 mg kg-1) with an average of 1575 mg kg-1.
.png)
Effect of carbon source and rock phosphate on microbial growth
Growth in all carbon source- rock phosphate combinations was observed, except in the cellulose + Sinda (C+S-RP) combination. Growth of glucose cultures was faster than in both the succinate and cellulose cultures (Table 2).
Growth was also observed in some of the no-soil (control) cultures (Figure 2; Table 2). No- soil control cultures with the all carbon sources tested and Chilembwe rock phosphate combinations (G+C-RP, S+C-RP, and C+C-RP) as well as G+S-RP and S+S-RP, showed growth of microorganisms encapsulating the rock phosphate. The growth observed in these control enrichment cultures differed and depended on the carbon-rock phosphate combinations. The G+S-RP combination resulted in three different forms of growth around the rock phosphate (Figure 2D-F). The cultures with C-RP showed more growth when compared to the S-RP cultures when cellulose was used as the carbon source, which exhibited no growth at all (Figure 3). The lack of growth was confirmed by failure of colonies to develop on corresponding C+S- RP plates when 100 µl of culture was plated.
.png)
.png)
.png)
Morphological characterization of isolates
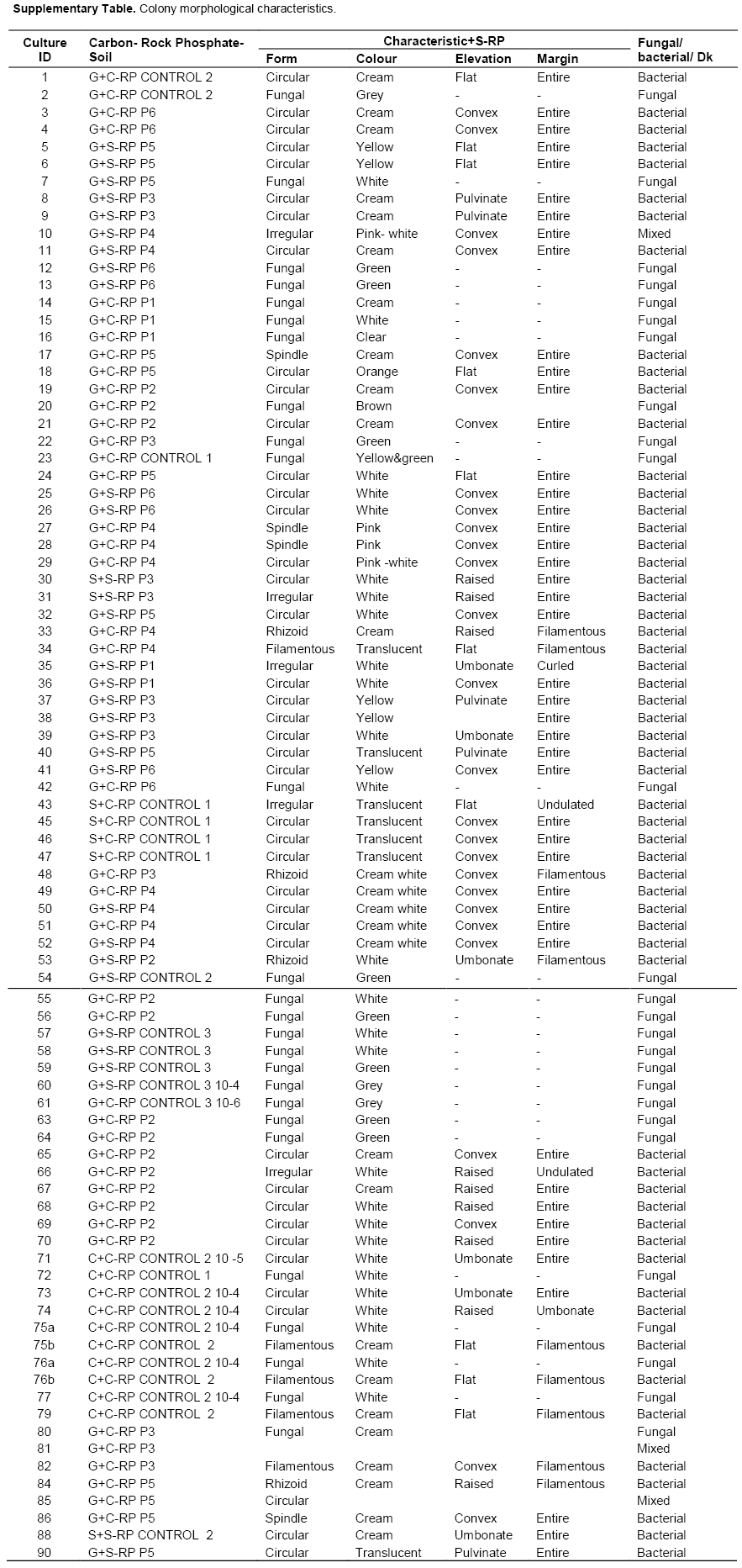
Once individual colonies were isolated, they were assigned a unique identity and their morphological characteristics recorded. Over 80 colonies were isolated and characterized (Supplemental Table 1). In summary, 41, 28, 8, 4 and 3 isolates were obtained from G + C-RP, G+SRP, C+C-RP, S+CRP and S+S-RP carbon source -rock phosphate combinations, respectively, giving a total of 86 (Table 3). Based on their characteristics, isolates were assumed to be either bacterial or fungal (Figures 4 and 5). The bacterial isolates had circular, irregular, rhizoid and spindle forms, with white, cream, pink, yellow and translucent colouration. All the isolates from the S+C-RP combination were translucent (Figure 4; Supplementary Table 1). The types of bacterial colony elevations observed included flat, convex, pulvinate and umbonate. Colony edges were filamentous, entire and undulate. The fungal isolates had mycelia with grey, green, white or yellow colouration. Of the 86 isolates, twenty representative bacterial isolates were chosen for further characterization.
.png)
.png)
.png)
Molecular characterization and tentative identification of isolates
Based on their 16S rRNA gene sequences, 20 isolates were identified as belonging to a total of seven different phyla. Five different bacterial genera were isolated from the C-RP enrichments, while four were isolated from the S- RP enrichments. Chilembwe and Sinda RP isolates had two genera in common, Burkholderia and Enterobacter. Isolates were most commonly identified asEnterobacter, representing 9 out of 20 sequenced isolates. The closest genetic matches for the remainingisolates were to type strains in the genera Bacillus,Burkholderia, Arthrobacter, Dyella, Curtobacterium andBeijerinckia (Table 4) and illustrated in a phylogenetic tree (Figure 6).
.png)

Acid phosphatase activity of selected isolates
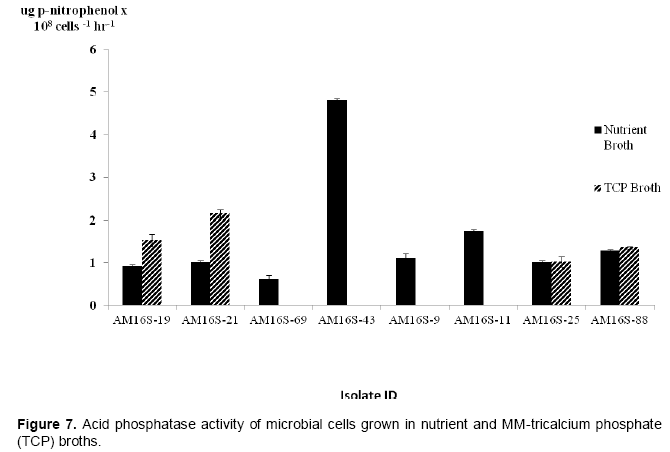
Only a subset of six isolates identified by 16S rRNA gene sequencing and two others were tested for growth in MM-TCP both and for phosphatase activity. All isolates grew in nutrient broth but three isolates could not use TCP as a phosphate source when grown in minimal medium (Table 5). The isolates that grew in both the nutrient and TCP broths were from the genus Enterobacter whereas Bacillus, Curtobacterium and two isolates without 16S rRNA gene sequences (Table 5, Figure 6) grew only in nutrient broth. Apart from AM16S-17, all isolates tested showed acid phosphatase activityranging from 0.62 to 4.80, and 1.02 to 2.15 µg PNP 108 cells-1 hr-1 in nutrient and MM-TCP broths, respectively (Figure 7). The highest enzyme activity (4.80 PNP x108 cells-1 h-1) was observed in an unidentified isolate (AM16S-43) from the control cultures of S+C-RP. Comparing isolates that grew in both media, the enzyme activity was higher in MM-TCP broth than NB for isolates from Chilembwe rock phosphate (AM16S-19 and AM16S-21) whereas it was about the same for isolates from Sinda rock phosphate (AM16S-25 and AM16S-88).

DISCUSSION
A total of 86 bacterial and fungal isolates growing on Chilembwe and Sinda rock phosphates were isolated from the soils and control cultures. Most (69) isolates were isolated from the cultures with glucose as the carbon source. This was attributed to more growth in glucose cultures resulting in more colonies on plates which were isolated compared to succinate and cellulose (Table 2). Glucose is more readily metabolised than cellulose and succinate by soil microorganisms. Out of the total, 24 were isolated from control plates or enrichment cultures without soil (Table 3). The observed growth in the control cultures was attributed to the possibility that rock phosphate solubilizing fungi and bacteria had colonized both the Chilembwe and Sinda rock phosphates, and that these organisms must have been in a form that was able to withstand the moist heat and pressure of the autoclaving process. Some rod-shaped, endospore forming bacteria, such as those of the genus Bacillus have been shown to possess this characteristic (Wolska et al., 2007). The isolate AM16S-88, tentatively identified as Enterobacterasburiae, from a control culture of succinate + Sinda-RP was among this group; species from this genus are known to be rod-shaped, non-spore forming Gram negative bacilli (Khunthongpan et al., 2013). This observation prompts further analysis of these isolates.
The type of rock phosphate appeared to influence microbial growth. The cultures with Chilembwe-RP showed more growth of microorganisms when compared to the Sinda-RP cultures (Table 3 and Figure 3). The observed differences were attributed to the possible differences in the mineralogy and subsequent solubility between the two rock phosphates. The Chilembwe-RP deposits has been characterized as being composed mainly of apatite, quartz, feldspar, biotite, hornblend, magnetite-haematite-goethite and ilmenite, with an average of 12 % P2O5 and a solubility of 1-1.7% P2O5 in neutral ammonium citrate (Appleton, 2002). In the absence of information on the mineralogical characteristics of the Sinda-RP, it can be inferred that Sinda-RP potentially has an even lower solubility than Chilembwe-RP, and therefore microorganisms metabolizing RP as a source of P would prefer the latter RP. Otherwise, othermineralogical attributes may be considered to be responsible for the observed differences.
Based on the morphological and molecular characterisation of the isolates, results show a wide range of characteristics and genera among the isolates so far identified. Previously, Bacillus spp., Rhizobia spp.,
Azotobacterspp., Arthrobacter spp. and Burkholderia spp. have been isolated and reported to have rock phosphate solubilizing capacities (Halder et al., 1991; Illmer and Schinner, 1992; Kumar et al., 2001). Fungal species such as Trichodermaspp., Aspergillusspp., Rhizoctonia spp. and Penicillium spp. have also been isolated and characterized for this capability (Fankem et al., 2014; Babana et al., 2013; Jacobs et al., 2002; Altomare et al., 1999; Motsara, et al., 1995). We report here the identification, based on the 16S rRNA gene sequences, of 20 isolates. The results indicate a large diversity of organisms capable of solubilizing rock phos-phate across the two rock phosphates, including two genera that have not previously been associated with P-solubilization, Dyella and Curtobacterium(Table 4; Figure 6). However, because Curtobacteriumpusillum (AM16S-9) could not grow in TCP broth, its capabilities to effectively solubilize RP remain questionable. The results show five different bacterial genera from the enrichments with Chilembwe-RP and four from the Sinda-RP.
The results of the phosphatase enzyme activity of the 10 isolates were comparable to previous research employing this method to measure acid phosphatase activity in bacterial isolates (Traoré, et al., 2013). Some of the microorganisms that had acid phosphatase activity failed to grow in TCP broth suggesting that they are incapable of utilizing both inorganic and organic forms of P. Previous studies have, however, shown that some microorganisms capable of solubilizing RP by producing low molecular weight organic acids also have acid phosphatase activity and that 70-80% of the microbial population in soils produce phosphatases (Plante, 2007). In this study, that the tested organisms were incapable of solubilizing tricalcuim phosphate after being isolated with RP as the only source of phosphorus may be explained by the composition of the RP. In addition, this observation does not agree with earlier findings that show that isolates preferred tricalcium phosphate to rock phosphate as a source of inorganic P in culture media (Asuming-Brempong and Keri, 2014).
In conclusion, over 80 isolates were isolated that were able to grow on Chilembwe and Sinda rock phosphates with varying morphological characteristics. Of these isolates, 20 were tentatively identified based on their 16S rRNA sequences, representing a large diversity of organisms capable of solubilizing rock phosphate. Biochemically, almost all the isolates tested had acid phosphatase enzyme activity. The ability to grow in culture, the genera represented as well as the enzyme activity of the isolates, all depended on the type of rock phosphate. The findings from this study provide a basis for developing appropriate seed inocula that can enhance availability of P from Chilembwe and Sinda RP resources for crop production.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Ademiluyi BO, Omotoso SO (2007). Comparative evaluation of TithoniadiversifoliaandNPKfertilizerforsoilimprovementinmaize(Zeamays) production in Ado Ekiti, South Western Nigeria. Res. J. Agron. 2:8-11. |
|
|
Altomare C, Norvell WA, Bjorkman T, Harman GE (1999). Solubilisation of phosphates and micronutrients by the plant growth promoting and biocontrol fungus TrichodermaharzianumRijai 1295-22. Appl. Environ. Microbiol. 65(7):2926-2933. |
|
|
Altschul SF, Madden TL, Schäffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ (1997). Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 25:3389-3402. |
|
|
Appleton JD (2002).Local rock phosphate resources for sustainable development in sub-Saharan Africa.British Geological Survey, Report CR/02/121/N, Nottingham.Retrieved. |
|
|
Asuming-Brempong S, Keri NK (2014). Isolation of phosphate solubilizing bacteria from tropical soil.Glob. Adv. Res. J. Agric. Sci. 3(1):8-15. |
|
|
Babana AH, Dicko AH, Maiga K, Traoré D (2013). Characterization of rock phosphate-solubilizing microorganisms isolated from wheat (Triticumaestivum L.) rhizosphere in Mali. J. Microbiol. Microb. Res. 1(1):1-6. |
|
|
Brempong AS (2013).Phosphate solubilising microorganisms and their ability to influence field of Rice. Agric. Sci. Res. J. 3(12):379-386. |
|
|
Cole JR, Wang Q, Fish JA, Chai B, McGarrell D M, Sun Y, Brown CT, Porras-Alfaro A, Kuske CR, Tiedje JM (2014). Ribosomal Database Project: data and tools for high throughput rRNA analysis Nucl. Acids Res.41(Database issue):D633-D642. |
|
|
Dick WA, Juma NG, Tabatabai MA (1983). Effects of soils on acid phosphatase and inorganic pyrophosphatase of corn roots. Soil Sci.136:19-25. |
|
|
Eivazi F, Tabatabai MA (1977). Phosphatases in soils. Soil Biol. Biochem. 9:167-172. |
|
|
Fankem H, Ngo NL, Nguesseu NG, Tchuisseu TGV, Tchiaze IAV, Nwaga D (2014). Rock phosphate solubilization by strains of Penicilliumspp. Isolated from farm and forest soils of three agro ecological zones of Cameroon. Am. J. Agric. For.2(2):25-32. |
|
|
Halder AK, Mishra AK, Chakrabartty PK (1991).Solubilization of inorganic phosphates by Bradyrhizobium. Ind. J. Exp. Biol.29:28-31. |
|
|
Illmer P, Shinner F (1992). Solubilization of inorganic phosphates by microorganisms isolated from forest soils. Soil Biol. Biochem. 24:389-395. |
|
|
Jacobs H, Boswell GP, Ritz K, Davidson FA, Gadd GM (2002). Solubilization of calcium phosphate as a consequence of carbon translocation by Rhizoctoniasolani.FEMS Microbiol. Ecol. 40:65-71. |
|
|
Jama BC, Niang A, Cachenco C, Nziguheha G, Amadalo B (2000). Tithoniadiversifolia as green manure for soil fertility improvement in western Kenya, A review. Agroforestry Syst. 49:204-212. |
|
|
Juma NG, Tabatabai MA (1988a). Comparison of kinetic and thermodynamic parameters of phosphomonoesterases of soils and of corn and soybean roots. Soil Biol. Biochem. 20:533-539. |
|
|
Juma NG, Tabatabai MA (1988b). Hydrolysis of organic phosphates by corn and soybean roots. Plant Soil 107:31-38. |
|
|
Juma NG, Tabatabai MA (1988c). Phosphatase activity in corn and soybean roots: Conditions for assay and effects of metals. Plant Soil107:39-47. |
|
|
Kang SC, Ha CG, Lee TG, Maheswari DK (2002).Solubilization of insoluble inorganic phosphates by a soil-inhabiting fungus Fomistopsis sp. PS 102.Curr. Sci. 82:439-442. |
|
|
Khan MS, Zaida A, Wani PA (2007). Role of phosphate-solubilising microorganism in sustainable agriculture- A review.Agron. Sustain. Dev. 27:29-43. |
|
|
Khunthongpan S, Bourneow C, H-Kittikun A, Tanasupawat S, Benjakul S, Sumpavapol P (2013). Enterobactersiamensissp.nov., a transglutaminase-producing bacterium isolated from seafood processing wastewater in Thailand. J. Gen. Appl. Microbiol. 59 (2):135-140. |
|
|
Kumar V, Behl RK, Narula N (2001). Establishment of phosphate solubilizing strains of Azotobacterchroococcum in the rhizosphere and their effect on wheat cultivars under greenhouse conditions.Microbiol. Res. 156: 87-93. |
|
|
Motsara MR, Bhattacharyya PB, Srivastava B (1995). Biofertilizers- their description and characteristics. In: A.B. Corner (Ed.), Biofertilizer technology, marketing and usage, a sourcebook-cum glossary, Fertilizer development and consultation organization, Pamposh Enclave, New Delhi. pp. 871-879. |
|
|
Nannipieri P, Giagnoni L, Landi L, Renella G (2011). Rock phosphatase enzymes in soil.In E.K. Bunemann, A. Oberson. E. Frossard (Eds.), Phosphorus in action, Soil Biol. 26: 215-243. |
|
|
Palm CA, Nziguheba G, Gachengo C, Gacheru E, Rao MR (1999). Organic materials as sources of phosphorous. Agroforestry Forum 9:30-33. |
|
|
Plante AF (2007). Soil biogeochemical cycling of inorganic nutrients and metals. In: E. A. Paul (Ed.), Soil microbiology, ecology and biochemistry (3rd ed.), Amsterdam. Academic Press |
|
|
Sharma SB, Sayyed RZ, Trivedi MH, Gobi TA (2013). Phosphate solubilizing microbes: sustainable approach for managing phosphorus deficiency in agricultural soils. SpringerPlus 2:587. |
|
|
Sievers F, Wilm A, Dineen DG, Gibson TJ, Karplus K, Li W, Lopez R, McWilliam H, Remmert M, Söding J, Thompson JD, Higgins DG (2011). Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 7:539. |
|
|
Sikazwe O, de Waele B (2004). Assessment of the quality and reserves of Bat Guano at Chiponwe and Kapongo caves near Lusaka as fertilizer material. UNZA J. Sci. Technol. Special Edition 32-42. |
|
|
Syer JK, Johnston AE, Curtin D (2008).Efficiency of soil and fertilizer phosphorus use.Reconciling changing concepts of soil phosphorus.Food and Agriculture Organisation of the UN Rome. |
|
|
Traoré L, Nakatsu CH, DeLeon A, Stott DE (2013). Characterization of six phosphate-dissolving bacteria isolated from rhizospheric soils in Mali. Afr. J. Microbiol. Res. 7(28):3641-3650. |
|
|
vanStraaten P (2002). Rocks for crops.Agro minerals of Sub - Sahara Africa.ICRAF, Nairobi, Kenya. |
|
|
vanStraaten P (2007). Agrogeology.The Use of rocks for crops. Cambridge, Ontario. Enviroquest Limited. |
|
|
Whalem JK, Sampedro L (2010). Soil ecology and management. Wallingford, Oxfordshire. CABI International |
|
|
Wolska KI, Grudniak AM, Kraczkiewicz-Dowjat A (2007). Genetic and physiological regulation of bacterial endospore development. Pol. J. Microbiol. 1:11-17. |
|
|
Yerokun OA (2008). Chemical characteristics of Phosphorus in some representative bench mark soils of Zambia. Geoderma147(1-2):63- 68. |
|
APPENDIX
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


