Full Length Research Paper

ABSTRACT
This study aimed to identify some important virulence factors, including pap, fim, sfa, aer and hly genes, typical of uropathogenic Escherichia coli (UPEC) in isolates collected from diarrheic and urinary tract infected patients in Khartoum State by multiplex polymerase chain reaction (PCR) assay. A total of 100 clinical specimens (50 urine and 50 diarrhea) were collected. Samples were cultured and identified by conventional method. Most study population were females 57/100 (57%); 42 suffering from urinary tract infections (UTIs) and 15 from diarrhea, while males were 43/100 (43%); 8 suffering from UTIs and 35 from diarrhea. Among enrolled subjects, 83 were positive for one or more uropathogenic E. coli virulent genes, while 17 isolates were negative for all genes. The results of multiplex PCR revealed that thirty two (n=32) diarrheal samples and fourteen (n=14) urine samples were aer positive. Thirty three (n=33) urine samples and eight (n=8) diarrheal samples appeared as fim positive. The genes pap and hly were found in 24 and 14 urine samples, respectively and in 9 and 3 diarrheal samples, respectively, while sfa gene was detected only in 15 urine specimens. The study concluded that fim gene was highly prevalent among UTI patients while aer gene was highly prevalent among diarrhea patients.
Key words: Uropathogenic Escherichia coli, fimH, aer, pap, sfa, hly, Sudan.
INTRODUCTION
MATERIALS AND METHODS

RESULTS
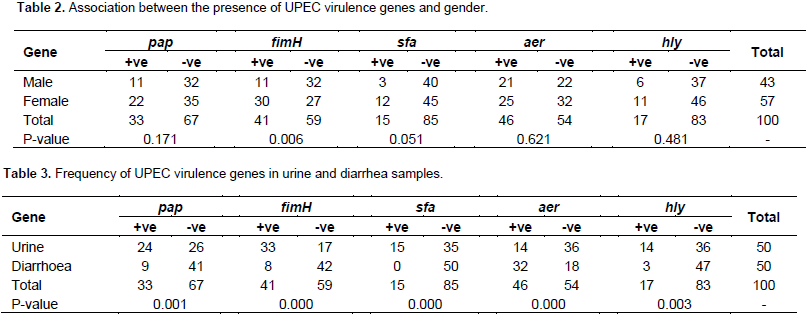
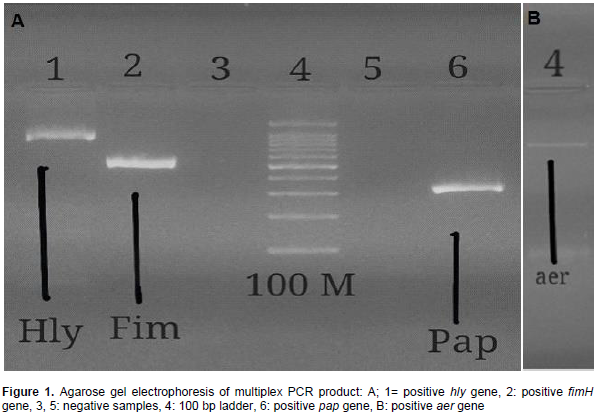
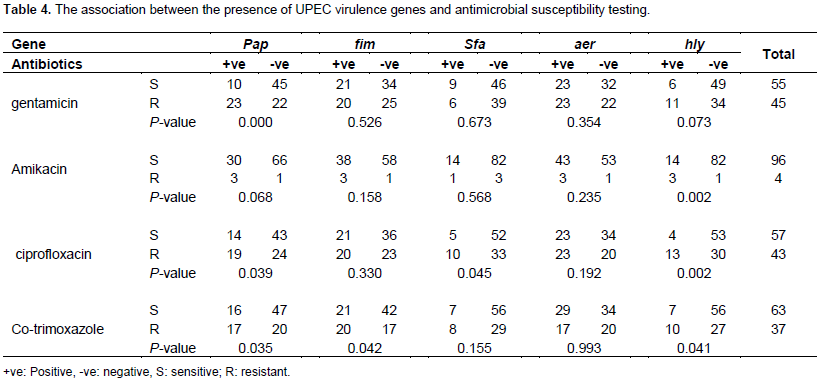
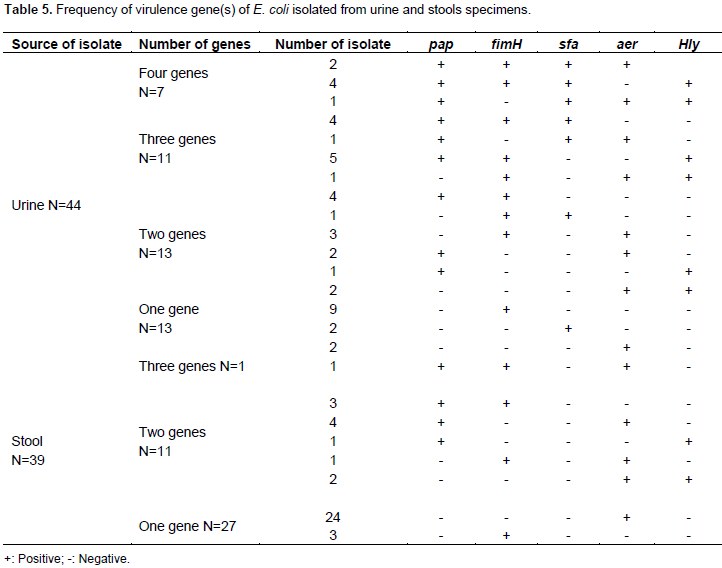
DISCUSSION
CONCLUSION
CONFLICT OF INTERESTS
REFERENCES
|
Abe CM, Salvador FA, Falsetti IN, Vieira MA, Blanco J, Blanco JE, Blanco M, Machado AM, Elias WP, Hernandes RT, Gomes TA (2008). Uropathogenic Escherichia coli (UPEC) strains may carry virulence properties of diarrhoeagenic E. coli. FEMS Immunol. Med. Microbiol. 52:397-406. |
|
|
Arisoy M, Aysev D, Ekim M, Özel D, Köse SK, Özsoy ED, Akar N (2005). Detection of virulence factors of Escherichia coli from children by multiplex PCR. Int. J. Clin. Prac. 60 (2):170-173. |
|
|
Bahalo S, Tajbakhsh E, Tajbakhsh S, Momeni M, Tajbakhsh F (2013). Detection of some virulence factors of Escherichia coli isolated from urinary tract infection isolated of children in Shahrekord Iran by Multiplex PCR. Middle-East J. Sci. Res. 14(1):29-32. |
|
|
Bahrani-Mougeot F, Gunther NW, Donnenberg MS, Mobley HLT (2002). "Uropathogenic Escherichia coli." In Escherichia coli: Virulence Mechanisms of a Versatile Pathogen. MS Donnenberg (ed). London: Academic Press. pp. 239-268. |
|
|
Blanco M, Blanco JE, Rodrıguez E, Abalia I, Alonso MP, Blanco J (1997). Detection of virulence genes in uropathogenic Escherichia coli by PCR comparison with results obtained using phenotypic methods. J. Clin. Micro. 31:37-43. |
|
|
Connell I, Agace W, Klemm P, Schembri M, Mărild S and Svanborg C (1996). Type 1 fimbrial expression enhances Escherichia coli virulence for the urinary tract. Proc. Nat. Acad. Sci. 93(18):9827-9832. |
|
|
Czaja CA, Stamm WE, Stapleton AE, Roberts PL, Hawn TR, Scholes D, Samadpour M, Hultgren SJ, Hooton TM (2009). Prospective cohort study of microbial and inflammatory events immediately preceding Escherichia coli recurrent urinary tract infection in women. J. Infect. Dis. 200:528-536. |
|
|
Derakhshandeh A1, Firouzi R, Motamedifar M, Arabshahi S, Novinrooz A, Boroojeni AM, Bahadori M, Heidari S (2015). Virulence Characteristics and Antibiotic Resistance Patterns among Various Phylogenetic Groups of Uropathogenic Escherichia coli Isolates. Jpn. J. Infect. Dis. 68(5):428-431. |
|
|
Dobrindt U, Blum-Oehler G, Nagy G, Schneider G, Johann A, Gottschalk G, Hacker J (2002). "Genetic structure and distribution of four pathogenicity islands (PAI I536 to PAI IV536) of uropathogenic Escherichia coli strain 536." Infect. Immunity 70:6365-6372. |
|
|
Eto DS, Jones TA, Sundsbak JL, Mulvey MA (2007). Integrin-mediated host cell invasion by type 1-piliated uropathogenic Escherichia coli. PLoS Pathog; 3:e100 |
|
|
Guiton PS, Cusumano CK, Kline KA, Dodson KW, Han Z, Janetka JW, Henderson JP, Caparon MG, Hultgren SJ (2012). Combinatorial small-molecule therapy prevents uropathogenic Escherichia coli catheter-associated urinary tract infections in mice. Antimicrob Agents Chemother. 56:4738-4745. |
|
|
Hannan TJ, Totsika M, Mansfield KJ, Moore KH, Schembri MA, Hultgren SJ (2012). Host–pathogen checkpoints and population bottlenecks in persistent and intracellular uropathogenic Escherichia coli bladder infection. FEMS Microbiol. Rev. 36:616-648. |
|
|
Hickling DR, Sun TT, Wu XR (2015). Anatomy and Physiology of the Urinary Tract: Relation to Host Defense and Microbial Infection. Microbiology spectrum 3(4): doi: 10.1128/microbiolspec.UTI-0016-2012. |
|
|
IBM SPSS Statistics for Windows, Version 21.0. (2012). Armonk, NY: IBM Corp, Released. www.ibm.com/software/analytics/spss/ IBM Corp. |
|
|
Jalali HR, Pourbakhsh A, Fallah F, Eslami G (2015). Genotyping of Virulence Factors of Uropathogenic Escherichia coli by PCR. Novelty in Biomed. 3(4):177-181. |
|
|
Johnson JR (2002) "Evolution of pathogenic Escherichia coli." In Escherichia coli: Virulence Mechanisms of a Versatile Pathogen. M. S. Donnenberg(ed). London: Academic Press, pp. 55-77. |
|
|
Johnson JR (1991). Virulence factors in Escherichia coli urinary tract infection. Clin Microbiol Rev. 4:80-128. |
|
|
Mabbett AN, Ulett GC, Watts RE, Tree JJ, Totsika M, Cheryl-lynn YO, Wood JM, Monaghan W, Looke DF, Nimmo GR, Svanborg C (2009). Virulence properties of asymptomatic bacteriuria Escherichia coli . Int. J. Med. Microbiol. 299:53-63. |
|
|
Meyrier A (1999). Urinary Tract Infection. In: Schrier RW, Cohen AH, Glassock RJ, Grünfeld JP. Atlas of diseases of the kidney (PDF). 2. Oxford: Blackwell Sci. ISBN 0-632-04387-3. |
|
|
Micenková L, Štaudová B, Bosák J, Mikalová L, Littnerová S, Vrba M, ŠevÄíková A, Woznicová V, Šmajs D (2014). Bacteriocin-encoding genes and ExPEC virulence determinants are associated in human fecal Escherichia coli strains. BMC Microbial. 14(1):109. |
|
|
Middendorf B, Hochhut B, Leipold K, Dobrindt U, Blum-Oehler G, Hacker J (2004) "Instability of pathogenicity islands in uropathogenic Escherichia coli 536." J. Bacteriol.186:3086-3096. |
|
|
Mobley HL, Green DM, Trifillis AL, Johnson DE, Chippendale GR, Lockatell, CV, Jones BD dan Warren JW (1990). Pyelonephritogenic Escherichia coli and killing of cultured human renal proximal tubular epithelial cells: role of hemolysin in some strains. Infect. Immunol. 58:1281. |
|
|
Morsi SS, Tash RM (2016). Virulence Determinants among Extended-Spectrum B- Lactamases producers of Uropathogenic Escherichia coli isolates In Zagagig University Hospitals, Egypt, Egyptian J. Med. Microbiol. 25(2):101-108. |
|
|
Obata-Yasuoka M, Ba-Thein W, Tsukamoto T, Yoshikawa H, Hayashi H (2002). Vaginal Escherichia coli share common virulence factor profiles, serotypes and phylogeny with other extraintestinal E. coli. Microbiology 148:2745-2752. |
|
|
Oliveira FA, Paludo KS, Arend LN, Farah SM, Pedrosa FO, Souza EM, Surek M, Picheth G, Fadel-Picheth CM (2011). Virulence characteristics and antimicrobial susceptibility of uropathogenic Escherichia coli strains. Genet. Mol. Res. 10(4):4114-25. |
|
|
Oswald E, de Rycke J, Lintermans P, van Muylem K, Mainil J, Daube G, Pohl P (1991). Virulence factors associated with cytotoxic necrotizing factor type two in bovine diarrheic and septicemic strains of Escherichia coli. J. Clin Microbiol. 29:2522-2527. |
|
|
Ribeiro TIBA M, Yano T, Da Silva LEITE D (2008). Genotypic characterization of virulence factors in Escherichia coli strains from patients with cystitis. Rev Inst. Med. Trop. S Paulo. 50(5):255-60. |
|
|
Salyers AA, Whitt DD (2002). Bacterial pathogenesis: a molecular approach. Washington DC: ASM Press. |
|
|
Schneider G, Dobrindt U, Brüggemann H, Nagy G, Janke B, Blum-Oehler G, Buchrieser C, Gottschalk G, Emödy L, Hacker J (2004) "The pathogenicity island-associated K15 capsule determinant exhibits a novel genetic structure and correlates with virulence in uropathogenic Escherichia coli strain 536. Infect. Immun. 72:5993-6001. |
|
|
Scholes D, Hooton TM, Roberts PL, Gupta K, Stapleton AE, Stamm WE (2005). Risk factors associated with acute pyelonephritis in healthy women. Ann. Int. Med.142:20-27. |
|
|
Schwartz DJ, Kalas V, Pinkner JS, Chen SL, Spaulding CN, Dodson KW, Hultgren SJ (2013). Positively selected fimH residues enhance virulence during urinary tract infection by altering fimH conformation. Proc Natl. Acad. Sci. USA.; 110:15530-15537. |
|
|
Tarchouna M, Ferjani A, Selma WB, Boukadida J (2013). Distribution of uropathogenic virulence genes in Escherichia coli isolated from patients with urinary tract infection. Int. J. Infect. Dis.17:450-3. |
|
|
Todar K (2007). Pathogenic E. coli, in Online Textbook of Bacteriology, University of Wisconsin-Department of Bacteriology.[homepage on internet] cited on -11-30. Available from View. |
|
|
Usein CR, Damian M, Tatuâ€Chitoiu D, Capusa C, Fagaras R, Tudorache D, Nica M, Bouguénec C (2001). Prevalence of virulence genes in Escherichia coli strains isolated from Romanian adult urinary tract infection cases. J. Cell. Mol. Med. 5(3):303-310. |
|
|
Welch RA, Burland V, Plunkett GI, Redford P, Roesch P, Rasko D, Buckles EL, Liou SR, Boutin A, Hackett J, Stroud D (. Extensive mosaic structure revealed by the complete genome sequence of uropathogenic Escherichia coli. Proceedings of the National Academy of Sciences. 99(26):17020-17024. |
|
|
Wullt B (2003). "The role of P fimbriae for Escherichia coli establishment and mucosal inflammation in the human urinary tract." Int. J. Antimicrob. Agents 21:605-621. |
|
|
Yamamoto S, Terai A, Yuri K, Kurazono H, Takeda Y, Yoshida O (1995). Detection of urovirulence factors in Escherichia coli by multiplex PCR. FEMS Immuno. Med. Microbiol. 12:85-90. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


