Full Length Research Paper

ABSTRACT
Four mustard cultivars possessing different degrees of resistance to Sclerotinia stem rot (Sclerotinia sclerotiorum) were used to study the biochemical and enzymatic basis of resistance. These include two moderately resistant cultivars (RGN-13 and RRN-505) and two susceptible cultivars (Bio-902 and T-59). Phenol content in all the mustard cultivars tested was found to increase at all three stages of inoculation, that is, 60, 75 and 90 days after sowing in diseased plants as compared to their respective healthy plants. Reduction in the contents of total sugars and reducing sugars were observed in diseased plants of all four cultivars irrespective of their susceptibility towards S. sclerotiorum. Peroxidase (PO) activity was observed to be high in moderately resistant cultivars as compared to susceptible ones. Maximum increase in PO activity was measured in diseased tissue of moderately resistant cultivar RRN-505 followed by RGN-13 and minimum PO activity in susceptible Bio-902. Significant increase in polyphenol oxidase (PPO) activity was recorded in moderately resistant cultivars as compared to susceptible cultivars. The phenylalanine ammonia lyase (PAL) activity was maximum in moderately resistant cultivar RGN-13. The PAL activity increased with days of inoculation in healthy as well as in diseased plants. Significant decrease in catalase activity was observed in diseased plants as compared to healthy ones and this decrease was more pronounced in susceptible cultivar T-59. The pathogen related enzymes polygalacturonase trans-eliminase (PGTE), pectin trans-eliminase (PME), polygalacturonase (PG) and cellulolytic (Cx) activities were found to be lower at initial stages of enzymes activity.
Key words: Enzymes, phenol, peroxidase, phenylalanine ammonia lyase and polyphenol oxidase.
INTRODUCTION
Oilseed crops in India account for almost 5% of Gross National Product (GNP) and 10% of value of agricultural products. Rapeseed mustard (Brassica) contributes 32% of the total oilseed production in India, and it is the second largest indigenous oilseed crop (Moza, 2006). Indian mustard [Brassica juncea (L.) Czern & Coss] is a major oilseed crop grown in rabi (post rainy) season in different regions of India. Sclerotinia stem rot or stem blight or white rot caused by Sclerotinia sclerotiorum (Lib.) de Bary has become a serious problem in mustard in North India. S. sclerotium is a necrotrophic plant pathogen that is able to infect more than 400 host plant species (Boland and Hall, 1994) and causes yield losses in a broad range of agricultural crops, including oilseeds, vegetables and pulses (Bolton et al., 2006). Diseases caused by S. sclerotiorum are often sporadic in occur-rence and severity, with losses in individual crops ranging from 0-100% (Krishnia et al., 2000; McDonald and Boland, 2004; Ghasolia et al., 2004; Shukla, 2005). The explosive pathogenicity of this fungus under favorable conditions and the ability of its sclerotia to withstand adverse conditions allow it to be a successful pathogen. Peroxidase (EC 1.11.1.7) and phenylalanine ammonia-lyase (EC 4.3.1.5; PAL) are two enzymes frequently associated with infection by phytopathogens (Hammerschmidt et al., 1982; Shirashi et al., 1989; Southerton and Deverall, 1990). The stimulation of the activities of these enzymes has been correlated with resistance to infection in many of the published reports, but conclusive evidence of their role in the defense mechanism(s) of plants is not yet available. The bio-chemical processes involved in the expression of resi-stance in B. napus are rarely known. However, the accu-mulation of a phytoalexin in Brassica sp. in relation to a hypersensitive reaction towards L. maculans has been reported (Rouxel et al., 1989). S. sclerotiorum is known to produce pectinolytic and cellulolytic enzymes (Lumdsen, 1969; Favaron et al., 1988; Marciano et al., 1982). The level of these enzyme activities correlates with the development of disease symptoms (Favaron et al., 1988; Lumdsen, 1976). Aside from pectic and cellulolytic enzymes, the diversity of polysaccharidases produced by S. sclerotiorum and the mechanisms controlling expres-sion of cell wall-degrading enzymes are poorly under-stood. Pectic enzymes are one of a complex of factors involved in pathogenesis, in certain diseases, they appear to be principal factor, in another, they apparently are of relatively little or of no importance. However, information on the biochemical and enzymatic basis of resistance of mustard to Sclerotinia stem rot is limited. The objective of this study was to examine in detail the changes in biochemical factors and enzymes in different cultivars of mustard governing resistance or susceptibility against S. sclerotiorum.
MATERIALS AND METHODS
Biochemical changes in infected host plants
To study the biochemical basis of Sclerotinia stem rot in healthy and infected plants of Indian mustard, host related enzymes, that is, peroxidase, polyphenol oxidase, phenylalanine ammonia lyase and catalase, pathogen related enzymes, polygalacturonasetran-seliminase (PGTE), pectin transeliminase (PTE), polygalacturonase (PG) and biochemical constituents, soluble phenolic contents and total sugars were estimated in two moderately resistant (RGN-13 and RRN-505) and two susceptible cultivars (T-59 and Bio-902) of healthy and diseased plants (stem samples), during rabi 2008-09. Mustard plants were raised in research farm at College of Agriculture, Swami Keshwanand Rajasthan Agricultural University, Bikaner following recommended agronomic practices. The plants were artificially inoculated at 60, 75 and 90 days after sowing.
Isolation, purification and artificially inoculation of Sclerotinia sclerotiorum
Small pieces of diseased tissues together with adjoining healthy area and sclerotia found in diseased stem were surface sterilized by dipping in mercuric chloride solution (1:1000) for two minutes followed by three washings with sterile water and blot dried then plated aseptically on Potato Dextrose Agar (PDA) in Petri dishes. These were incubated in BOD incubator for growth of the fungus at 27 ± 2°C. Sub cultures from pure peripheral growth were made on PDA slants and Petri dishes. Plant materials were inoculated following the method of Reglinski et al. (1997) with some modifications. Young, healthy plants aged 60, 75 and 90 days, were selected from mustard field raised in research farm. Using a sharp needle, small injuries were made in the middle of half the stem of each cultivar. Mycelial bit (4 mm in diameter) taken from the growing edge of 3-day-old cultures on potato dextrose agar were used for inoculation. Inoculum bits were placed with the mycelium side down in the middle of each stem injury. Control stem were treated with uninfected PDA bits. The stems were then immediately sealed with two layers of parafilm to maintain high humidity. All the biochemical constituents were estimated in fresh samples of healthy and diseased plants of Indian mustard after 7 days of inoculation with the pathogen. The stem samples were collected and stored in deep freeze at -30°C till further use for biochemical analysis of phenol, total sugars and enzyme activity of the sample.
Estimation of soluble sugar content
The soluble sugar content of the leaf samples was determined by using the method of Hedge and Hofreiter (1962). The soluble sugar content of the leaf samples was determined by using the method of Hedge and Hofreiter (1962). A 100 mg of the sample was hydrolyzed by keeping it in a boiling water bath for 3 h with 5 ml of 2.5 N HCl and cooled at room temperature. After that the sample was neutralized with solid sodium carbonate until the effervescences ceased and centrifuged. The supernatant was collected and 0.5 and 1 ml aliquots was took for analysis. The standard was prepared by taking 0, 0.2, 0.4, 0.6, 0.8 and 1ml of the working standard in which ’0’ served as blank. The volume was made to 1 ml in all the tubes including the sample tubes by adding distilled water followed by addition of 4 ml anthrone reagent. The green to dark green colour was read at 630 nm. Standard graph was drawn by plotted concentration of the standard on the x-axis versus absorbance on the Y-axis and calculations were made to observe the carbohydrate in samples.
Reducing sugar content was measured by following “Nelson’s modification of “Somogyi’s method” (Somogyi, 1952) using arseno-molybdate colour forming reagent and two copper reagent “A” (sodium carbonate anhydrous 2.5 g, potassium sodium tartrate 2.5 g, sodium bi carbonate 2.0 g, sodium sulphate 20.0 g and distilled water 80.0 ml) and “B” (copper sulphate, 15 g concentrated sulphuric acid, 1 drop and distilled water, 80 ml). 1 ml of leaf sample (100 times diluted) was added with a mixture of 1 ml copper reagent prepared from 24 part of copper “B” solutions. This mixture in test tubes was heated in boiling water bath, cooled, added with the colour-forming reagent (arseno-molybdate) and absorbance was measured at 620 nm on Spectronic-20. The value was plotted against a standard curve prepared from glucose.
Estimation of soluble phenol content
The soluble phenol content was estimated by the method described by Thimmaiah (1999). One gram of fresh mustard leaf sample was macerated in pestle and mortar with 10 ml ethanol 80%. The homo-genate was centrifuged at 10,000 g for 20 min. The supernatant was pooled and evaporated to dryness in water bath. The residue was dissolved in 5 ml distilled water. An aliquot of 0.2 ml was transferred in test tube and the volume was made to 3 ml with distilled water. Folin ciocalteau reagent (0.5 ml) was added to each test tube. After three minutes, 2 ml of 20% Na2CO3 was added in each tube and mixed thoroughly. The tubes were then kept in boiling water for one minute and the absorbance was measured at 650 nm against reagent blank in spectrophotometer. The standard curve was prepared using different concentrations of catechol. The phenol content was expressed as mg phenols/g fresh tissue.
Estimation of soluble protein content
The soluble protein content of the leaf samples was determined by using the method of Lowry et al. (1951). One gram of fresh leaf was macerated in mortar with 5 ml 0.1 M sodium phosphate buffer (pH 7.0). The homogenate obtained was centrifuged at 16,000 g for 20 min. The supernatant was used for estimation of soluble protein content. For this purpose, 0.2% sodium carbonate (anhydrous) in 0.1 N NaOH (Solution A) was prepared. Similarly, 0.5% copper sulphate (CuSO4 5H2O) in 1% sodium potassium tartarate (freshly made) was prepared and was regarded as solution B. From these two reagents, solution C (alkaline copper solution) was prepared by mixing 50 ml of solution A with 1 ml of solution B just before use. An aliquot of 0.1 ml of supernatant was taken in the test tube and the volume was made to 1 ml with distilled water followed by addition of 5 ml solution C, mixed well and incubated at room temperature for ten minutes. 0.5 ml of Folin ciocalteau reagent was diluted, mixed well and incubated at room temperature in dark for 30 min. The absorbance was recorded at 660 nm against blank. The amount of protein in sample was determined from the standard curve prepared by using different concentrations of bovine serum albumin.
Host related enzymes
Estimation of peroxidase (EC 1.11.1.17) activity: Peroxidase activity was assayed by the method described by Thimmaiah (1999). One gram leaf sample was macerated in previously chilled mortar in 10 ml ice cold 0.1 M phosphate buffer at pH 6.0. The homogenate was strained through two fold of muslin cloth and centrifuged at 16,000 g for 20 min at 4°C. The supernatant was used as enzyme source. In order to assay the enzyme activity, 1 ml 0- dianisidine, 0.5 ml H2O2, 1 ml of phosphate buffer and 2.4 ml of distilled water were pipetted in test tubes. The blank was prepared by excluding H2O2 and adding additional volume of water in the place of H2O2. The reaction was initiated by adding 0.2 ml of enzyme extract (supernatant) and incubating at 30°C for five minutes. The reaction was stopped by adding 1 ml of 2 N H2SO4.
The absorbance was measured at 430 nm against reagent blank. The unit of enzyme was defined as absorbance/min/mg protein.
Estimation of polyphenol oxidase (EC1.14.18.1) activity: The polyphenol oxidase activity was determined by the method of Mayer et al. (1965). One gram leaf sample was homogenized in 2 ml 0.1 M sodium phosphate buffer at pH 6.5 and centrifuged at 16,000 g for 15 min at 4°C. The supernatant was used as enzyme source. The reaction mixture consisted of 0.2 ml enzyme extract (supernatant) and 1.5 ml of 0.1 M sodium phosphate buffer (pH 6.5). To start the reaction, 0.2 ml of 0.1 M catechol was added and the change in absorbance was recorded at 30 s intervals for up to 3 min at 495 nm. The activity of enzyme was expressed as absorbance/min/mg protein.
Estimation of phenylalanine ammonia lyase (EC 4.3.1.5) activity: Phenylalanine ammonia lyase activity was assayed by the method described by Thimmaiah (1999). Three gram leaf sample was macerated in a mortar in presence of 2.6 ml of 0.2 M sodium borate buffer (pH 8.7) containing 2-mercaptoethanol (0.8 ml/L). The homogenate was filtered through cheese cloth. Acetic acid (0.1M) was added drop by drop to bring the pH of filtrate to 5.5. Protamine sulphate solution (0.002 g + 0.008 ml of 1 M sodium acetale buffer pH 5.5 diluted 0.1 ml) was added to filtrate and stirred for 10 min followed by centrifuging at 7,000 g for 10 min. The supernatant was used as enzyme source. In order to determine the enzyme activity, an assay mixture consisting of 1 ml of 0.05 M Tris-HCl buffer (pH 8.8), 0.5 ml of 0.01 M L-phenylalanine and 0.4 ml of distilled water was incubated at 30°C for 5 min. The reaction was initiated by adding 0.1 ml enzyme extract (supernatant) and again incubated at 30°C for 60 min. The blank without L-phenylalanine was run. The reaction was stopped by adding 0.5 ml of 1 N HCl. The residue was dissolved in 3 ml of 0.05 N NaOH. The absorbance was recorded at 290 nm. The standard curve was prepared using different concentrations of cinnamic acid. The unit of enzyme was expressed as µmoles cinnamic acid produced/min/mg protein.
Estimation of catalase (EC 1.11.1.6) activity: Catalase activity was measured by adopting the procedure of Sinha (1972). The reaction mixture containing 0.4 ml of 0.2 M H2O2, 0.1 ml of enzyme extract and 0.5 ml of 0.01 M phosphate buffer (pH 7.0) was incubated at 37°C for 1 min along with continuous shaking and then 3 ml of dichromate reagent (5% potassium dichromate; glacial acetic acid, 1:3) was added. The mixture was heated for 10 min in a boiling water bath. After cooling, the intensity of colour was measured at 570 nm. The enzyme activity was expressed as m moles of H2O2 utilized min-1 mg-1 protein.
RESULTS AND DISCUSSION
In the present studies, an attempt was made to analyze the activity of host related enzymes, pathogen related enzymes, soluble phenolic compounds and total sugars in healthy and diseased plants of moderately resistant and susceptible cultivars of Indian mustard inoculated with S. clerotiorum at three stages: 60, 75 and 90 days after sowing.
Changes in biochemical constituents in healthy and infected host plants
Effect on phenolic contents
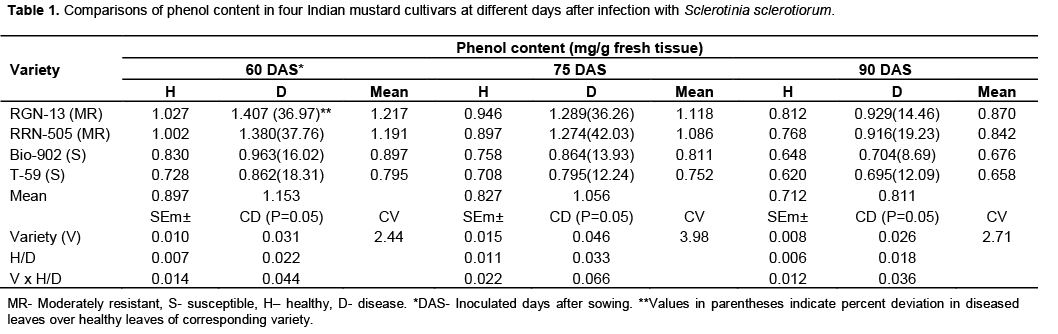
There was a significant increase in phenolic content of mustard stem due to S. sclerotiorum infection as com-pared to healthy plants (Table 1 and Figure 1). Phenol content in all the mustard cultivars tested viz., RGN-13, RRN-505, Bio-902 and T-59 was found to be increased at all three stages of inoculation: 60, 75 and 90 days after sowing in diseased plants as compared to their respective healthy plants (Table 1). Data revealed that the total phenolic contents were highest in diseased plants of moderate cultivar, RRN-505 increase of total phenolic compounds was observed in moderately resi-stant cultivar RGN-13 and RRN-505 as compared to susceptible cultivars Bio-902 and T-59 at 60,75 and 90 days after sowing. Gupta et al. (1990) also found similar trend for phenolic compounds in mustard and reported that resistant cultivar RC-781 possessed higher amount of phenols than susceptible genotypes. The higher level of total phenols in diseased leaves with A. brassicae as compared to healthy leaves in the four cultivars of Indian mustard was also reported by Gupta and Kaushik (2002). In contrast, decrease in total phenol of the leaves of Mentha arvensis with increase in initial inoculum of S. sclerotiorum was also reported by Perveen et al. (2010). Beniwal et al. (2008) also reported higher quantity of phenolic contents in resistant cultivars of wheat in response to flag smut than susceptible and highly susceptible cultivars. The level of total phenol contents indicated the involvement of phenolics in resistant mechanism through the phenomenon of oxidation to quinones which are more toxic to the pathogens (Arora and Bajaj, 1981; Bajaj et al., 1983).
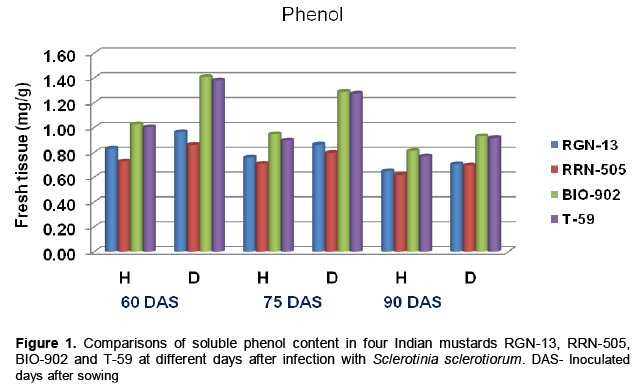
Sugars
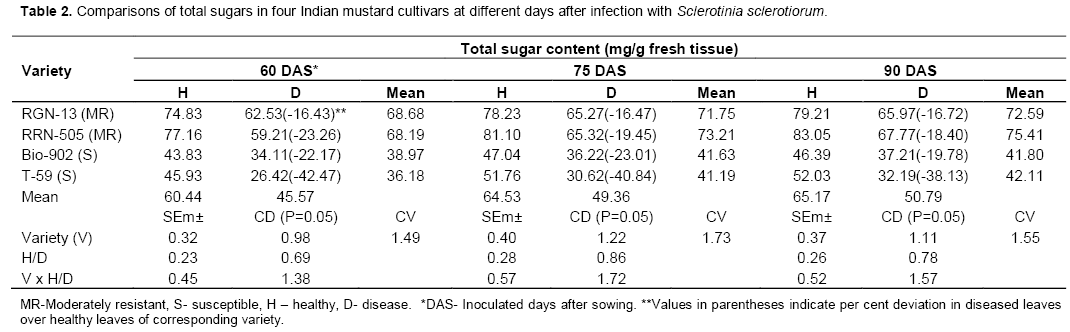
Significant decrease in total sugars was observed in S.sclerotiorum infected plant tissues of both moderately resistant cultivars as well as susceptible cultivars at all three stages (Table 2 and Figure 2). The total sugars also decreased with the days of sowing, it was higher when mustard cultivars were inoculated at 60 days of sowing as compared to 90 days of sowing in all four cultivars (Table 2). Similar trend was also found in reducing sugars (Table 3 and Figure 3). In the present studies, total, reducing and non-reducing sugars were observed to be low in Sclerotinia infected stem tissues of moderately resistant and susceptible cultivars of mustard. However, in similar study, Jobic et al. (2007) observed a decline in sugars and amino acids in the plant and fungus total content during infection. Sucrose and fructose, initially present almost exclusively in plant, were reduced by 85%. Kiran et al. (2003) reported increased amount of reducing and total sugars in calli of Brassica species subcultured on medium having different concentrations of culture filtrate of Alternaria brassicae. Guleria and Kumar (2006) studied biochemical basis of host-pathogen interaction between white stem rot and rapeseed-mustard in field experiment and found that total sugars content in the plant tissue of S. sclerotiorum resistant Brassica cultivars viz., JTC-1 and PCC-5 was found to be lower than susceptible cultivars viz., Neelam and Sheetal. Kumar et al. (1998) also reported reduction in total sugars in sunflower seeds due to infection of S. sclerotiorum.
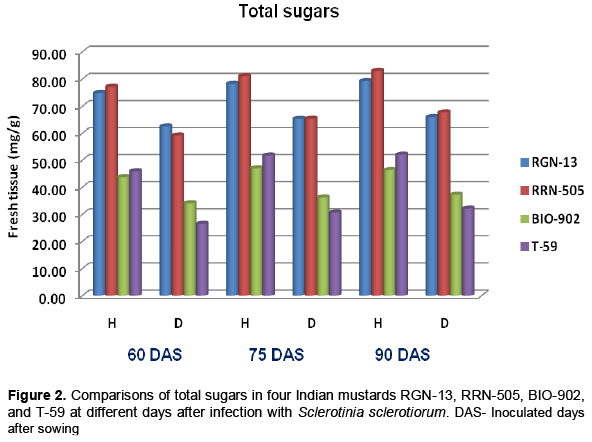
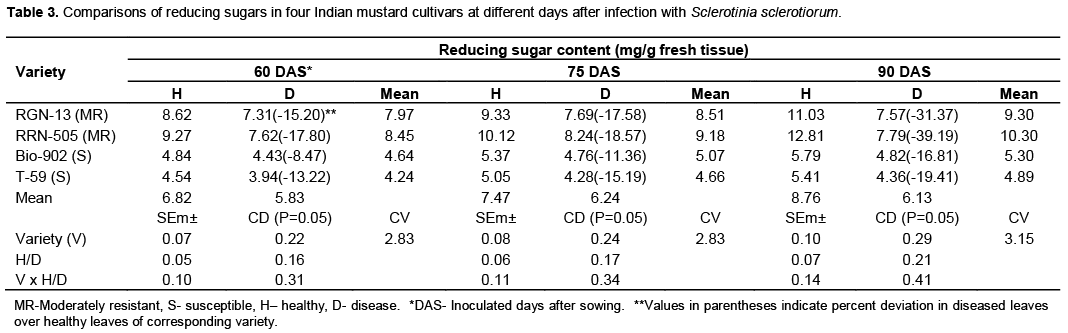
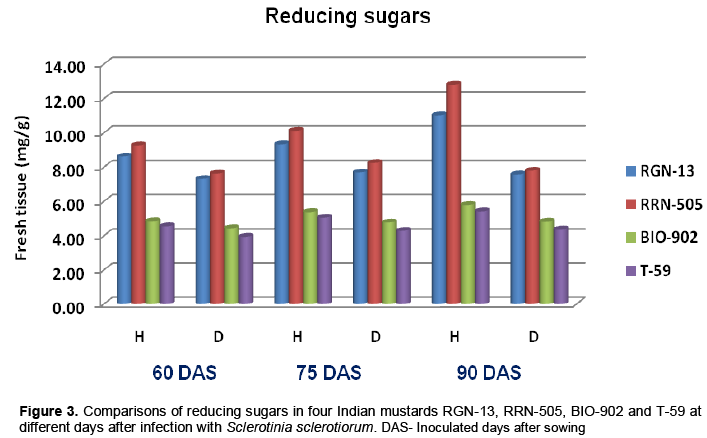
Host related enzymes
Effect on peroxidase (PO) and polyphenol oxidase (PPO)
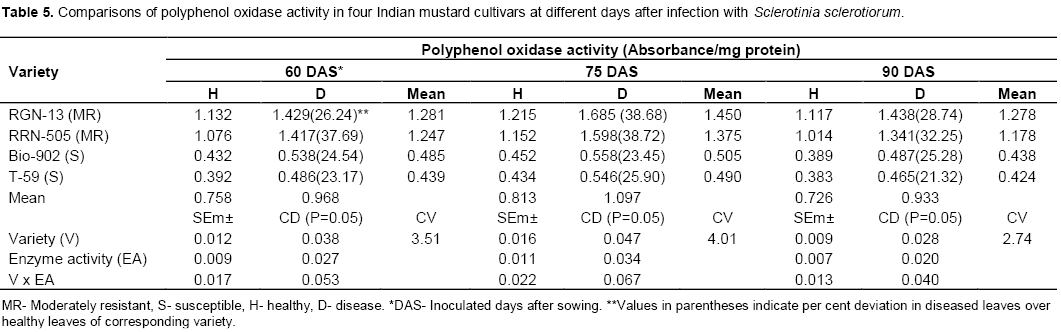
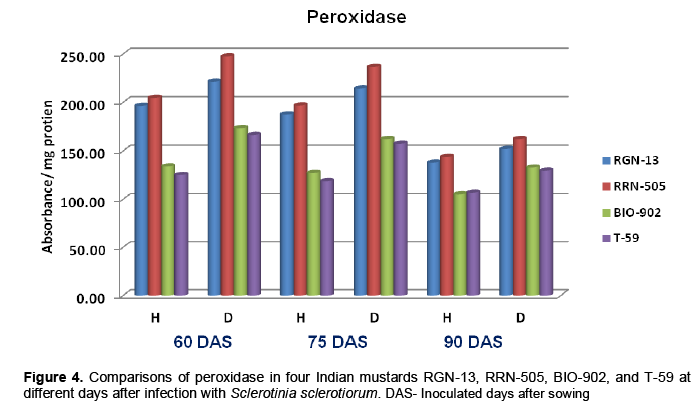
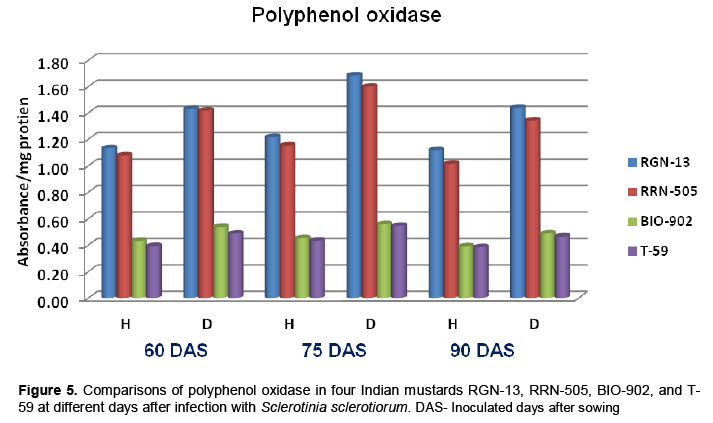
The findings of the present study revealed that the peroxidase (PO) and polyphenol oxidase (PPO) activities were higher in diseased plants as com-pared to healthy plants of mustard (Tables 4 and 5). The enzyme activity was also higher in moderately resistant genotypes in comparison with susceptible mustard genotypes. The enzyme PO and PPO activity were decreased with age of plant in all four cultivars of mustard. A sharp increase in PPO and PO activities following infection was observed in all the cultivars which appeared to be more associated with susceptibility (Table 4-5 and Figure 4-5). Similar increases in both enzymes in many other host-parasite combinations have been documented (Gangopadhyay and Lal, 1986; Gowda et al., 1989; Gupta et al., 1990). Watpade and Mehta (2012) reported activity of oxidative enzymes viz., polyphenol oxidase and peroxidase increase in tolerant calli with the increase in concentration of culture filtrate of S. sclerotiorum, whereas catalase activity increased in susceptible calli in response to increase in concentration of culture filtrate. In contrast, the lower PPO activity was observed in susceptible genotypes at both stages (pre and post infection) of determination (Mahatma et al., 2008).
.png)
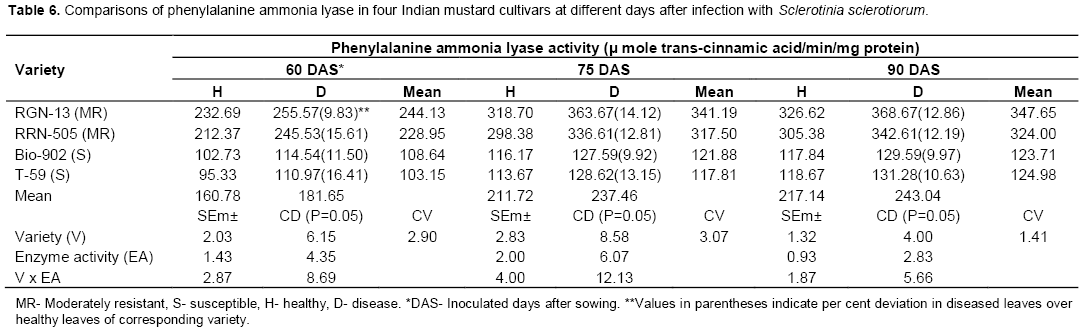
Phenylalanine ammonia lyase (PAL)
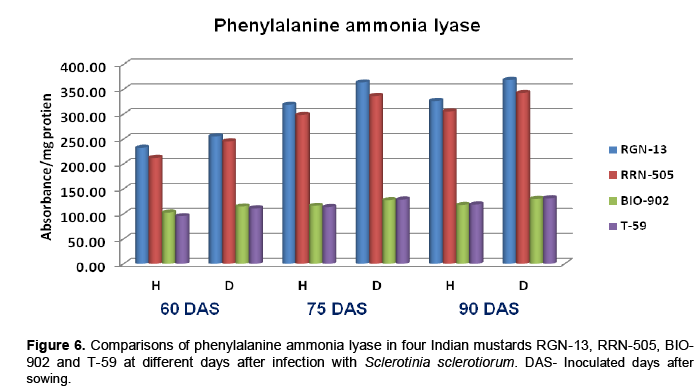
Phenylalanine ammonia lyase (PAL) activity was increased in diseased plants when compared with healthy plants tissue in all the mustard cultivars used, at all three stages of inoculation (Table 6). The PAL activity was significantly higher in mode-rately resistant cultivars as compared to susceptible cultivars. The enzyme activity increased uniformly with age of plant (Table 6 and Figure 6). Chakraborty et al. (1993) determined activities of peroxidase and phenylalanine ammonia lyase in leaves of healthy and inoculated with Leptosphaeria maculans in Brassica napus cutivars and observed increased activity of both enzymes in moderately resistant cultivars. These findings corroborate with observa-tions made by Joshi et al. (2004) and Muthukumar (2007) in Alternaria leaf spot of cluster-bean and tuberose plants, respectively. Chakraborty et al. (2004) reported the association of defense enzymes peroxidase, polyphenol oxidase and phenylalanine ammonia lyase in resistance mechanism in tea leaves against Exobasidium vexans.
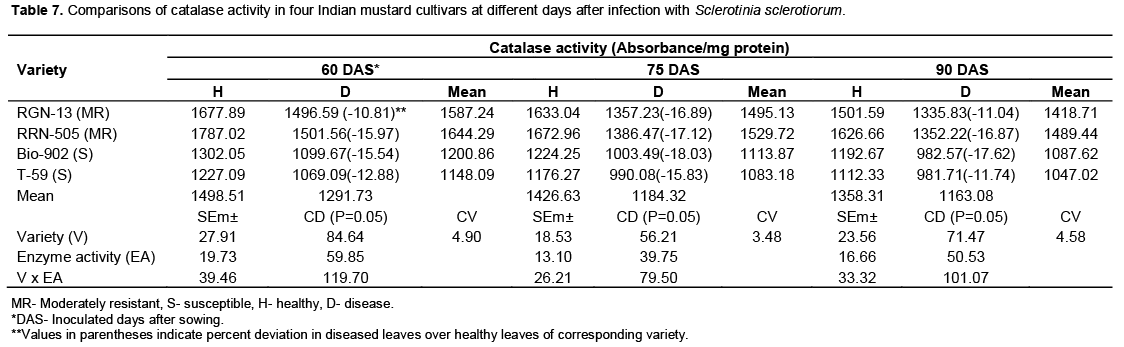
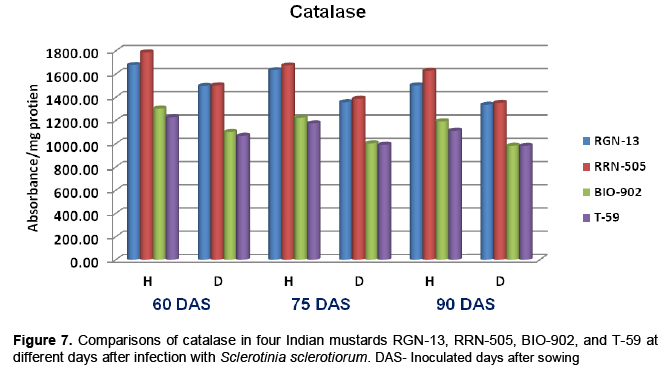
Catalase
The healthy plants of moderately resistant cultivar RGN-13 and RRN-505 had higher catalase activity in comparison with susceptible Bio-902 and T-59. When the diseased plants were analyzed for catalase activity, it was observed that there was significant decrease in catalase activity in diseased plants as compared to healthy plants in all four cultivars and this decrease was more pronounced in susceptible cultivar (Table 7 and Figure 7). Contrary to PPO and PO, the activity of catalase remained lower in diseased plants as compared to healthy ones. The present findings are in agreement with the earlier reports made by the Saharan et al. (2001) that the healthy leaves of resistant cultivars of cluster bean to Alternaria blight had higher catalase activity than susceptible cultivars both at 65 and 80 days after sowing. Gupta et al. (1995) reported that catalase activity was maximum at 40 DAS in healthy leaves, which declined after this stage in all the mustard species. The presence of this enzyme in substantial amount at initial stages of plant growth indicates its possible role in tissue. Changes in peroxidase and catalase activity in healthy and diseased fruits of mango due to black tip were also reported by Agrawala et al. (1960).
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
The authors are grateful to Dean, College of Agriculture, Bikaner for providing necessary facilities and the Vice Chancellor, S. K. Rajasthan Agricultural University, Bikaner for providing financial assistance during the course of the study
REFERENCES
|
Agrawala SC, Sharma CP, Kumar A (1960). The effect of black-tip disease on the catalase and peroxidase activity of mango fruits. Curr. Sci. 5:195. |
|
|
Arora YK, Bajaj KL (1981). Activities of enzymes of polyphenol metabolism in Phaseolus aureus seedlings germinated in the presence of 2-chloroethylphosphoric acid. Biol. Plant. 23:141-144. |
|
|
Bajaj KL, Arora YK, Mahajan R (1983). Biochemical differences in tomato cultivars resistant and susceptible to Meloidogyne incognita. Rev. Nematol. 6:143-145. |
|
|
Beniwal MS, Karwasra SS, Chhabra ML (2008). Biochemical changes in wheat plants infected with flag smut. Indian Phytopathol. 61(2): 243-246. |
|
|
Boland GJ, Hall R (1994). Index of plant hosts of Sclerotinia sclerotiorum. Can. J. Plant Pathol. 16:93-108. |
|
|
Bolton MD, Thomma BPHJ, Nelson BD (2006). Sclerotinia sclerotiorum (Lib.) de Bary: biology and molecular traits of a cosmopolitan pathogen. Mol. Plant Pathol. 7:1-16. |
|
|
Chakraborty BN, Das-Biswas R, Sharma M (2004). Multi component coordinated defense strategies in tea plants against Helopeltis theivora and Exobasidium vexans. J. Plant. Crop 32 : 289-297. |
|
|
Chakraborty U, Chakraborty BN, Kapoor M (1993). Changes in the levels of peroxidase and phenylalanine ammonia lyase in Brassica napus cultivars showing variable resistance to Leptosphaeria maculans. Folia Microbiol. 38(6): 491-496. |
|
|
Favaron F, Alghisi P, Marciano P, Magro P (1988). Polygalacturonase isoenzymes and oxalic acid produced by Sclerotinia sclerotiorum in soybean hypocotyls as elicitors of glyceollin. Physiol. Mol. Plant Pathol .33:385–395. |
|
|
Gangopadhyay S, Lal S (1986). Changes in certain biochemical constituents in maize (Zea mays L.) leaf sheath infected with Rhizoctoniasolani. Indian J. Plant Pathol. 4:9-16. |
|
|
Ghasolia RP, Shivpuri A, Bhargava AK (2004). Sclerotinia rot of Indian mustard in Rajasthan. Indian Phytopathol. 57 (1): 76-79. |
|
|
Gowda B, Bhat GS, Bhat SS (1989). Peroxidase and polyphenol oxidase activities in sorghum and Perenosclerospora sorghi interaction. Curr. Sci. 58 (18):1037-1039. |
|
|
Guleria S, Kumar A (2006). Biochemical basis of white stem rot, Sclerotinia sclerotiorum resistance in rapeseed-mustard. J. Oilseeds Res. 23(1):69-71. |
|
|
Gupta SK, Gupta PP, Kaushik CD (1995). Changes in leaf peroxidase, polyphenol oxidase, catalase and total phenol due to Alternaria blight in Brassica species. Indian J. Mycol. Plant Pathol. 25 (3): 175-180. |
|
|
Gupta SK, Gupta PP, Yadava TP, Kaushik CD (1990). Metabolic changes in mustard due to Alternaria leaf blight. Indian Phytopathol. 43:64-69. |
|
|
Gupta SK, Kaushik CD (2002). Metabolic changes in mustard leaf and siliqua well due to the infection of Alternaria blight (Alternaria brassicae). Cruciferae Newsletter 24: 85-86. |
|
|
Hammerschmidt R, Nuckles EM, Kuc J (1982). Association of enhanced peroxidase activity with induced systemic resistance of cucumber to Colletotrichum lagenarium. Physiol. Plant Pathol. 20: 73- 82. |
|
|
Hedge JE, Hofreiter BT (1962). In: Carbohydrate Chemistry, 17 (Eds. Whistler R.L. and. Be Miller, J.N.), Academic Press, New York. |
|
|
Jobic C, Boisson AM, Gout E, Rascle C, Fèvre M, Cotton P, Bligny R (2007). Metabolic processes and carbon nutrient exchanges between host and pathogen sustain the disease development during sunflower infection by Sclerotinia sclerotiorum. Planta 226 (1): 251-265. |
|
|
Joshi UN, Gupta PP, Gupta V, Kumar S (2004). Biochemical factors in clusterbean that impart Alternaria blight resistance. J. Mycol. Plant Pathol. 34 (2):581-583. |
|
|
Kiran, Dhingara HR, Mehta N, Sangwan MS (2003). Effect of culture filtrate of Alternaria brassicae on biochemical constituents of calli of Brassicas. J. Mycol. Plant Pathol. 33:51-55. |
|
|
Krishnia SK, Meena PD, Chattopadhyay C (2000). Seed-yield and yield- attributes of Indian mustard affected by Sclerotinia rot. J. Mycol. Plant Pathol. 30:265. |
|
|
Kumar B, Chahal SS, Ahuja KL (1998). Effect of Sclerotinia head rot on some bioconstituents of sunflower seed. Indian Phytopathol. 51:359-360. |
|
|
Lowry OH, Rosebrough NJ, Farn AL, Randal RJ (1951). Protein measurement with Folin-Phenol reagent. J. Biol. Chem. 193: 265-275. |
|
|
Lumdsen RD (1969). Sclerotinia sclerotiorum infection of bean and the production of cellulase. Phytopathology 59:653-657. |
|
|
Lumdsen RD (1976). Pectolytic enzymes of Sclerotinia sclerotiorum and their localization in infected bean. Can. J. Bot. 54:2630-2641. |
|
|
Mahatma MK, Bhatnagar R, Rawal P (2008). Changes in enzymes and pruline levels in leaves of downy mildew resistant and susceptible pearl millet genotypes. J. Mycol. Plant Pathol. 38 (2):277-281. |
|
|
Marciano P, Di Lenna P, Magro P (1982). Polygalacturonase isoenzymes produced by Sclerotinia sclerotiorum in vivo and in vitro. Physiol. Plant Pathol. 20:201-212. |
|
|
Mayer AM, Harel E, Shaul RB (1965). Assay of catechol oxidase, a critical comparison of methods. Phytochemistry 5 : 783-789. |
|
|
McDonald MR, Boland GJ (2004). Forecasting diseases caused by Sclerotinia spp. in eastern Canada: fact or fiction. Can. J. Plant Pathol. 26:480-488. |
|
|
Moza MK (2006). A report on the National Conference on "New developments in rapeseed mustard". Curr. Sci. 90 (9): 1174-1175. |
|
|
Muthukumar A (2007). Changes in leaf POD, PPO PAL and total phenols due to Alternaria leaf spot in tuberose plants. J. Mycol Plant Pathol. 37(2):349-351. |
|
|
Perveen K, Haseeb A, Shukla PK (2010). Effect of Sclerotinia sclerotiorum on the disease development, growth, oil yield and biochemical changes in plants of Mentha arvensis. Saudi J. Biol. Sci. 17(4): 291–294. |
|
|
Reglinski T, Poole PR, Whitaker G, Hoyte SM (1997). Induced resistance against Sclerotinia sclerotiorum in kiwifruit leaves. Plant Pathol. 46:716-721. |
|
|
Rouxel T, Sarniguet A, Kollman A, Bousquew JF (1989). Accumulation of a phytoalexin in Brassica spp. in relation to a hypersensitive reaction to Leptosphaeria maculans. Physiol. Mol. Plant Pathol. 34: 507-517. |
|
|
Saharan MS, Saharan GS, Singh JV (2001). Inheritance of resistance in clusterbean to Alternaria blight (Alternaria cucumerima var. cyamopsidis) disease. J. Mycol. Plant Pathol. 31:72-75. |
|
|
Shirashi T, Yamaoka N, Kunch H. (1989). Association between increased phenylalanine ammonia-lyase activity and cinnamic acid synthesis and the induction of temporary in accessibility caused by Erysiphe graminis primary germ tube penetration of the barley leaf. Physiol. Mol. Plant Pathol. 34:75 -83. |
|
|
Shukla AK (2005). Sclerotinia rot- its prevalence in Indian mustard at different levels of nitrogen. Indian Phytopathol. 58 (4): 493-495. |
|
|
Sinha AK (1972). Colorimetric assay of catalase. Analt. Biochem.47: 389-394. |
|
|
Somogyi M (1952). Notes on sugar determination. J. Biol. Chem. 195 : 19-23. |
|
|
Southerton SG, Deverall BJ (1990). Changes in phenylalanine ammonia-lyase and peroxidase activities in wheat cultivars expressing resistance to the leaf-rust fungus. Plant Pathol. 39:223-230. |
|
|
Thimmaiah (1999). Standard Methods of Biochemical Analysis. Kalyani Publishers, Ludhiana. 534pp. |
|
|
Watpade S, Mehta N (2012). Effect of culture filtrate of Sclerotinia sclerotiorum (Lib.) de Bary on the activities of oxidative enzymes in calli of Brassica species. Plant Dis. Res. 27 (2):182-185. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


