Full Length Research Paper

ABSTRACT
The combination of microorganisms in consortia enhances high bioflocculant yields. The study aimed at producing and characterising bioflocculant from Bacillus pumilus JX860616 and Alcaligenes faecalis HCB2. Bioflocculant TPT-1 was obtained through ethanol extraction after optimum conditions were established. The characteristics of the bioflocculant TPT-1 were obtained by scanning electron microscope equipped with elemental detector, Zetasizer nano, ultraviolet-visible spectrophotometry. Fourier transform infrared (IR) spectrophotometry and the thermal decomposition was used to conduct the thermal gravimetric analysis. The flocculation mechanism of TPT-1 on Kaolin suspension was obtained by Zetasizer Nano. Glucose and yeast extract were the best energy sources, yielding 3.0 g/L of TPT-1 in optimum conditions (30°C, 165 rpm, initial pH 6 and 72h). TPT-1 revealed to be an anionic, heat stable glycoprotein, with the total carbohydrate content of 83.1% (w/w) and the total proteins content of 9.7% (w/w). The elemental analysis demonstrated the presence of N (1.3), C (15.0), O (44.8), P (0.8), Ca (9.0), Cl (2.8), Mg (0.4), S (12.1), K (11.4) and Na (1.9) in mass proportion (% w/w), while the IR spectrum showed the presence of hydroxyl, carbonyl and amine groups. Ba2+ mediated bridge flocculation mechanism between the bioflocculant TPT-1 and Kaolin particles. The high flocculating capability (90%) and characteristics of TPT-1 suggested its potentiality in industrial applications.
Key words: Bioflocculant TPT-1, flocculating activity, flocculation mechanism and bacterial consortium.
INTRODUCTION
Flocculation is the natural process whereby flocculants are used to flocculate, settle and remove particles, suspended solids and colour in solutions (Cong-Liang et al., 2012). Flocculating agents are widely used in industrial fields such as dredging, textile dyeing, mining, pharmacology, cosmetology, wastewater treatment, food and fermentation processes (Zhang et al., 1999; Salehizadeh and Shojaosadati, 2001). Flocculants are grouped as inorganic (aluminium salts), synthetic (polyacrylamide) and natural occurring flocculants (bioflocculants and chitosan) (Okaiyeto et al., 2015). Inorganic and synthetic organic flocculants are extensively used in vast biotechnological applications due to their cost effectiveness. However, they are both reported to impose health risks and environmental hazards (Salehizadeh and Shojaosadati, 2001). Synthetic flocculants have shown adverse health effects such as neurotoxicity, carcinogenicity and cause Alzheimer’s diseases (Serdar et al., 2011). Moreover, chemical flocculants are non-degradable and their by-products are often toxic (Okaiyeto et al., 2015). Owing to the short-comings, the need for eco-friendly flocculants that impose less health threats is of high demand. Bioflocculants are macromolecules secreted by microorganisms due to substrate metabolism, microbial growth, cell lysis and degradation of microorganisms or microbial components (Carlos et al., 2011). Bioflocculants lack secondary pollution and thus considered as environmental friendly substances. This is due to the special bioflocculant components (carbohydrates, proteins, nucleic acid and lipid), microbial flocculant matrix that show biodegradability adsorption abilities and hydrophilicity or hydrophobicity (More et al., 2014). However, production costs and low flocculant yields have been restrictive factors in the industrial applicability of bioflocculants. Moreover, the lack of knowledge of the active ingredients of most bioflocculants and the mechanism involved during flocculation also decreases their market potential (Barredo, 2005). In nature, microorganisms do not live in isolation; they coexist with many other microorganisms forming synergism and symbiosis relationships (Chapelle, 2001). Zhu et al. (2004) and Zhang et al. (2007) have reported that the combination of strains of microorganisms in consortia produce bioflocculants that have better flocculating activity and higher bioflocculant yields than pure strains. Bacillus pumilus JX860616 and Alcaligenes faecalis HCB2 produced biofloccculants (2.4 and 2.7 g/L, respectively), with high flocculating activities of 93.3 and 92.1%, respectively. Thus, in attempt to improve the quantity and quality of the bioflocculants, a consortium of these bacterial strains was constructed and the characteristics and flocculation mechanism of the produced bioflocculant was determined.
MATERIALS AND METHODS
Bacterial strains
Bacterial strains were previously isolated from Sodwana Bay sediment in the north of KwaZulu-Natal, (RSA). The bacterial strains were identified by 16S rRNA nucleotide sequencing genes with subsequent BLAST analyses. Nucleotide sequences were placed in GenBank and the repository accession numbers were Bacillus pumilus JX860616 and Alcaligenes faecalis HCB2. The bacterial strains were then preserved in 20% glycerol stock at −80°C in the Department of Biochemistry and Microbiology, University of Zululand, (RSA).
Production media
The standard fermentation medium by Zhang et al. (2007) was used. The medium composed of glucose (20.0 g), KH2PO4 (2.0 g), K2HPO4 (5.0 g), (NH4)2SO4 (0.2 g), NaCl (0.1 g), CH4N2O (0.5 g), MgSO4 (0.2 g) and yeast extract (0.5 g) in a litre of filtered seawater and was autoclaved at 121°C for 15min.
Bacterial consortium
B. pumilus JX860616 and A. faecalis HCB2 were used in combination to construct bacterial consortium. A loopful of each strain colony was inoculated separately into 50 ml of the fermentation medium and incubated for 72 h at 30°C with the shaking speed of 165 rpm. The preculture was then used as the standard inoculum. For construction of the bacterial consortium, 1ml of the standard inoculums of bacterial strains were both inoculated into the same 50ml of the fresh fermentation medium and incubated at 30°C at the shaking speed of 165 rpm. After 72h, the culture broth was centrifuged at 8000 × g for 30 minutes at 4°C and the supernatant was used for determination of flocculating activity (FA) (Zhang et al., 2007).
Determination of flocculating activity
The method by Kurane et al. (1986) was adapted to access the flocculating activity. 100 ml of Kaolin clay solution (4 g/1000 ml) was mixed with 3 ml of 1% w/v CaCl2 and 2 ml of obtained supernatant. The mixture was vigorously agitated, poured into measuring cylinder and left to stand at room temperature. After 5min, the optical density (OD550) was read and the flocculating activity determined using the formula:
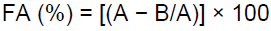
Where A was the optical density of Kaolin suspension at 550nm and B was optical density of a sample at 550 nm.
Optimisation of flocculation conditions
The effect of the different inoculum sizes (1 to 5%) of the constructed consortium on flocculating activity were assessed (Zhang et al., 2007). Bacterial species were cultivated in various organic carbon sources (glucose, fructose, sucrose, lactose, maltose, xylose, starch and molasses) and nitrogen sources (casein, beef extract, yeast extract, ammonium sulphate urea and peptone) for determination of suitable carbon and nitrogen sources for bioflocculant production (Okaiyeto et al., 2016). The concentration of 20 g/L of culture medium was used for carbon sources and 1.2 g/L for nitrogen sources. The effect of initial pH on flocculating activity was determined using a pH meter (Eutech instruments-pH 700) (Liu et al., 2010). The pH of the culture medium was adjusted by 0.1 M HCl and 0.1 M KOH in the range of 3 to 12. The effect of cultivating temperatures (20 to 45°C) on the flocculating activity were assessed and various metal cations (NaCl, KCl, LiCl, MnCl2, BaCl2 and FeCl3) were also used to evaluate their effect on flocculating activity by replacing the CaCl2 used in an original medium (Zhao et al., 2013).
Time course assay
The effect of fermentation time on flocculating activity was conducted in accordance with a previous study by Cosa et al. (2013). Bacterial cultures in consortium were incubated under obtained optimal growth conditions. From the fermentation broth, samples were drawn every 12h for 120h and the final pH and optical density (OD550) of the broth were determined. The fermentation broth was then centrifuged (8000 × g, 30 min) including the supernatant used for evaluation of FA.
Extraction and purification bioflocculant TPT-1
The extraction and purification of bioflocculant TPT-1 was done in accordance with the standard method by Chang et al. (1998). After 72h of incubation, the culture broth was centrifuged (8000 × g, 4°C, 30min). The distilled water (1000 ml) was added to the supernatant phase and centrifuged (4000 × g, 30 min at 4°C) to remove the insoluble substances. Ethanol (2000 ml) was added to the supernatant, agitated and then left at 4°C to precipitate overnight. The precipitate was vacuum-dried and the crude product dissolved in 100ml of distilled water. Hundred millilitre of the mixture of chloroform and butanol (5:2 v/v) was then added, agitated and left to settle overnight at room temperature. The supernatant was again centrifuged (4000 × g, 4°C, 30 min) and vacuumed-dried.
Physicochemical characterisation of bioflocculant TPT-1
The morphological structure of the bioflocculant, Kaolin floc and Kaolin particles were determined by scanning electron microscope (SEM) (SEM-Sipma-VP-03-67) and the electrical charge of the bioflocculant was analysed by Malvern Zetasizer Nano. The total sugar content was evaluated by phenol-sulphuric acid method (Chaplin and Kennedy, 1994). Ultraviolet-visible spectrophotometry was used to qualitatively determine the protein content (Harrington and Raper, 1968), and the total protein concentration was determined by Bradford, (1976) method. Ninhydrin method was quantitatively used to determine the presence of amino acid (Kay et al., 1956). The elemental analysis was carried out with SEM equipped with elemental analyser. Prior to SEM analysis, 5mg of bioflocculant, was added on slides coated with silicon and fixed by spin coater at rpm, 60s. The functional groups of the produced TPT-1 were assessed by a Fourier transform infrared (IR) spectroscopy (Okaiyeto et al., 2013). The solubility test was done in different solvents (water, benzene, acetone, ethyl acetate, chloroform, dichloromethane, ethanol, hexane, methanol and butanol) (Zaki et al., 2011). Determination of optimum dosage of the produced bioflocculant was done in different dosages (0.2 to 1.0 mg/ml) (Okoh and Ugbenyen, 2014)
Effect of cations, salinity, pH and thermal on the flocculating activity of TPT-1
The synergistic effect of cations (KCl, NaCl, MnCl2, CaCl2, BaCl2 and FeCl3) on the flocculating activity of the bioflocculant was evaluated (Okaiyeto et al., 1013). The effect of salinity on bioflocculant activity was determined by varying NaCl concentrations (5 to 35 g/L) in Kaolin solution (4g/L). The effect of pH on the flocculating activity of the purified bioflocculant was assayed according to Okaiyeto et al. (2013), whereby the pH of Kaolin solutions in separate flasks were adjusted using KOH and HCl in a range of 3 to 12 prior to the assessment of the flocculating activity of the bioflocculant. Thermal stability of the purified bioflocculant was determined at different temperatures (50 to 100°C) (Wang et al., 2011). The pyrolysis analysis of the purified bioflocculant was assessed at the range of 22 to 900°C at a rate constant of 10°C min-1 under constant flow of nitrogen gas using thermo-gravimetric instrument (Okaiyeto et al., 2013).
Zeta potential
The zeta potential of the bioflocculant TPT-1, Kaolin solution, Kaolin solution with BaCl2, Kaolin solution with both BaCl2 and bioflocculant TPT-1 were measured by a ZetaSizer Nano (Malvern, UK) to determine and propose the flocculation mechanism involved during flocculating of Kaolin particles in suspension by of bioflocculant TPT-1 (Aljuboori et al., 2015). The pH of the suspensions were adjusted to pH 3 using HCl or KOH. The zeta potential of each sample was determined using a Malvern Zetasizer Nano with clear disposable cuvettes at 25°C. The cuvettes were thoroughly washed and 2.5 ml of samples were injected carefully whilst avoiding any air bubbles in the cuvette.
Statistical analysis
The data was obtained in triplicate experimentation and subjected to one-way analysis of variance (ANOVA) using the MINITAB Student Release 12 statistical package. A significance level of p Ë‚ 0.05 was used.
RESULTS
Inoculum size
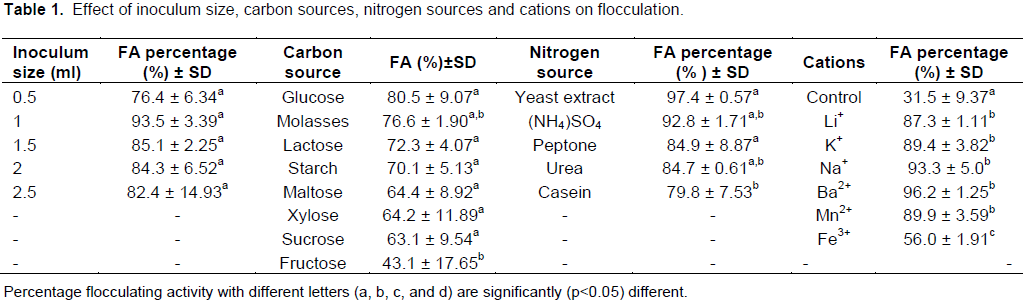
Table 1 displayed the effect of inoculum size of mixed bacterial isolates on bioflocculant TPT-1 production. The inoculum size of 2% (v/v) gave the highest flocculating activity (93%) compared to the other four sizes. The increase or decrease in inoculum size led to a slight decrease in the flocculating activities. Thus, an inoculum size of 2% was used in all experiments.
Effect of carbon and nitrogen sources on flocculating activity
Bacteria in consortium utilized various carbon and nitrogen sources in the culture medium. Glucose was the most preferred carbon source with the highest flocculating activity of 80.5%, followed by lactose with 70.5% (Table 1). Fructose, sucrose, maltose, xylose, starch and molasses were poorly utilised by the bacteria for bioflocculant production as shown by the low flocculating activities which were generally less than 70%. The effect of different nitrogen sources on bioflocculant production was studied and the results were illustrated in Table 1. All the nitrogen sources showed a potential to be used in bioflocculant production as they resulted in flocculating activities above 80%. Ammonia sulphate was the most preferred nitrogen source with the highest flocculating activity of 97% while casein had the least flocculating activity (80%).
Cations effect on flocculating activity
Table 1 demonstrated the effect of metal cations on flocculating activity. All the cations used, including the control (without cation), stimulated flocculation process and resulted in over 70% of flocculating activities. Ba2+ was the most active and preferred metal cation, resulting with the highest flocculating activity of 96%. Fe3+ showed the lowest flocculating activity of 77%.
Effect of temperature on flocculating activity
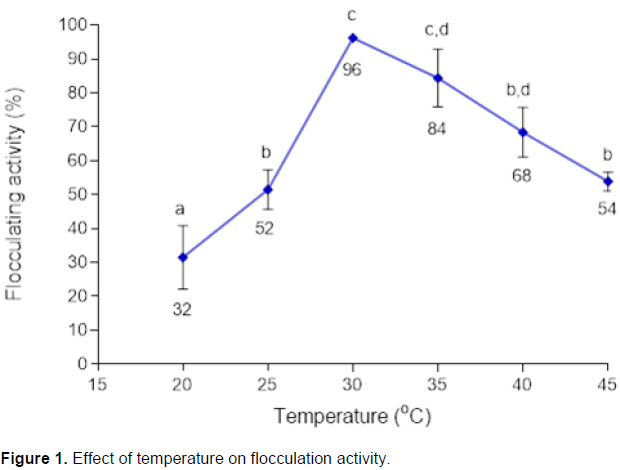
Fermentation temperature has an impact on bioflocculant production. Figure 1 illustrated the effect of fermentation temperatures on flocculation activity. When the fermentation temperature was 30°C, the flocculating activity reached the maximum of 96%. The increase or decrease in fermentation temperature above or below 30°C resulted in the decrease in flocculating activity. Thus, 30°C was used as the optimal temperature for the bioflocculant production.
Effect of Initial pH on flocculating activity
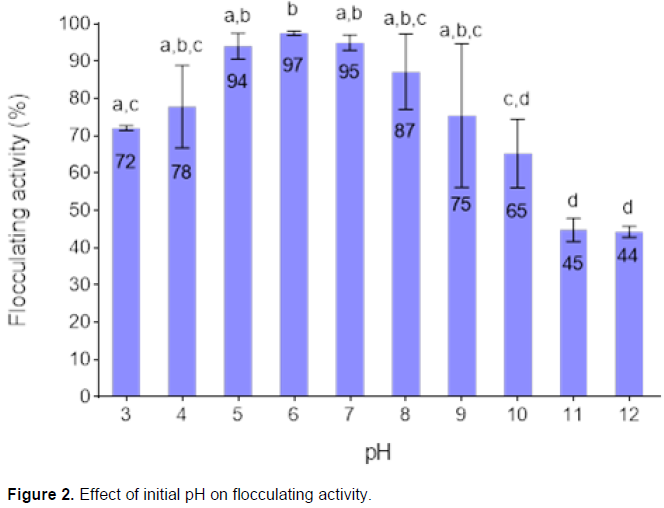
Figure 2 shows the effect of initial pH of the fermentation medium on the flocculating activity. The bacteria in consortium maintained over 80% of flocculating activity over the weak acidic (pH 5 to 6) and alkaline (pH 8), with the highest flocculating activity (97%) at pH of 6. It was observed that highly alkaline pH 10 to 12 did not favour bioflocculant production and resulted in poor flocculating activity of less than 70%; pH 12 gave the least flocculating activity of 44%.
Time course
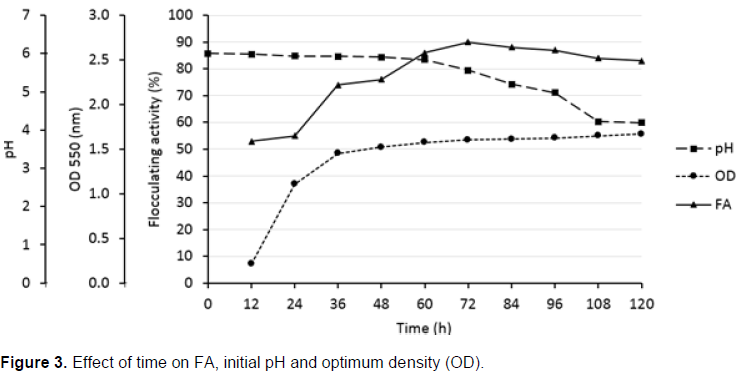
The effect of time course on flocculating activity (FA), bacterial growth (shown as optical density (OD)) and pH are shown in Figure 3. The flocculating activity increased relatively to the bacterial growth until 72 h of fermentation. The highest flocculating activity (90%) was obtained after 72 h. A slight decrease in flocculating activity was observed after 72 h. The initial pH of the medium dropped constantly from the initial pH of 6.0 to the final pH of 4.3.
Bioflocculant yield and solubility
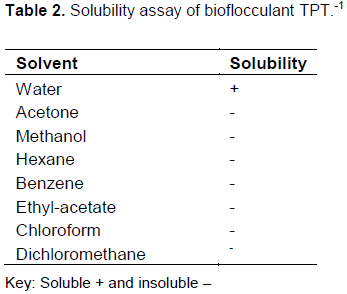
The purified bioflocculant TPT-1 of 3.0 g was obtained from 1 L of fermentation broth of consortium of B. pumilus JX860616 and A. faecalis HCB2. The extracted and purified bioflocculant was insoluble in all solvents, with the exception of water as a solvent (Table 2).
Effect of bioflocculant dosage size on flocculating activity
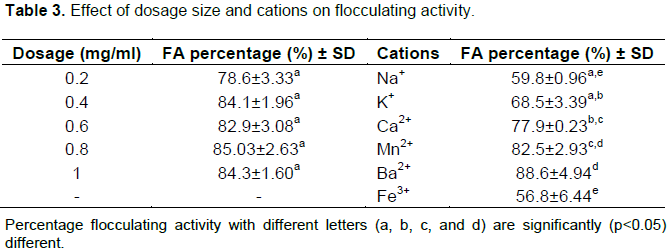
Table 3 shows the results obtained during dosage size assay. The high flocculating activity of 84% was obtained at the low bioflocculant concentration of 0.2 mg/ml. Although 0.2 mg/ml is half of 0.8 mg/ml in concentration, it gave significantly high flocculation activity (84%) and was thus the more preferred over 0.8 mg/ml that had 86% of flocculating activity.
Effect of metal cations on flocculating activity of the bioflocculant
Cations enhance flocculating rate by neutralizing and stabilizing the residual negative surface charge of the functional groups on the bioflocculant. The flocculating activity of bioflocculant was greatly stimulated by the addition of Li+, Mn2+ and Ba2+ at concentrations of about 1% (Table 1). These cations showed the flocculating activity above 80%. Ba2+ was the most preferred metal cation with the flocculating activity of 89%, while Fe3+ was the least cation with the flocculating activity of 56%.
Physicochemical composition of the bioflocculant TPT-1
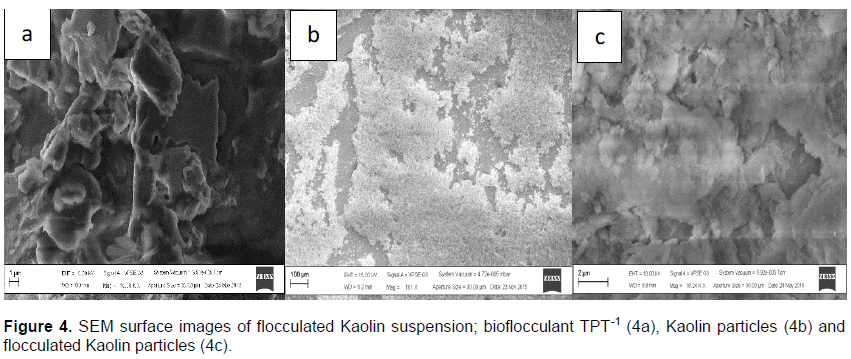
Figure 4 shows SEM surface images. TPT-1 had a porous and crystal-like morphology (Figure 4a). Kaolin particles revealed a fine and smooth structure (Figure 4b) and the clump like structure was observed in flocculated Kaolin particles (Figure 4c).
Electrical charge of the bioflocculant
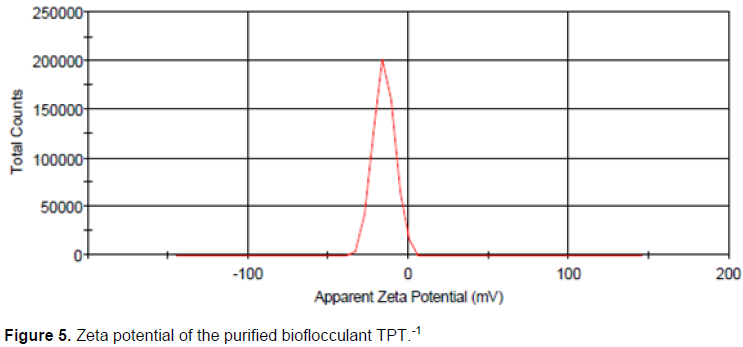
The surface charge of TPT-1 was determined and the results were illustrated in Figure 5. The bioflocculant had a surface negative charge of -15.1 mV, as illustrated by the ZetaSizer Nano. The zeta potential of TPT-1 revealed the bioflocculant as an anionic biomolecule.
Chemical composition of bioflocculant TPT-1
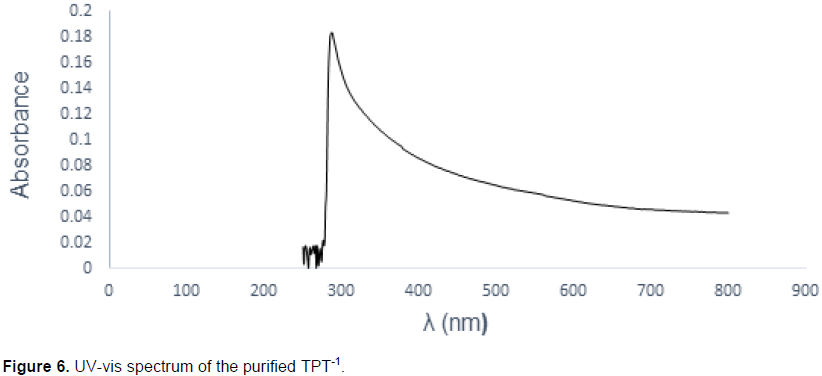
TPT-1 was Ninhydrin positive. The UV-vis spectrum of the bioflocculant demonstrated a sharp absorption peak at 289 nm, which was a characteristic of proteins (Figure 6). Thus, the results suggested that the bioflocculant had protein content.
Chemical composition of bioflocculant TPT-1
Table 4 shows the quantitative chemical composition of TPT-1. The bioflocculant was predominantly composed of the total carbohydrates (83.1%) and trace protein content (9.7%).

Elemental and functional group analysis
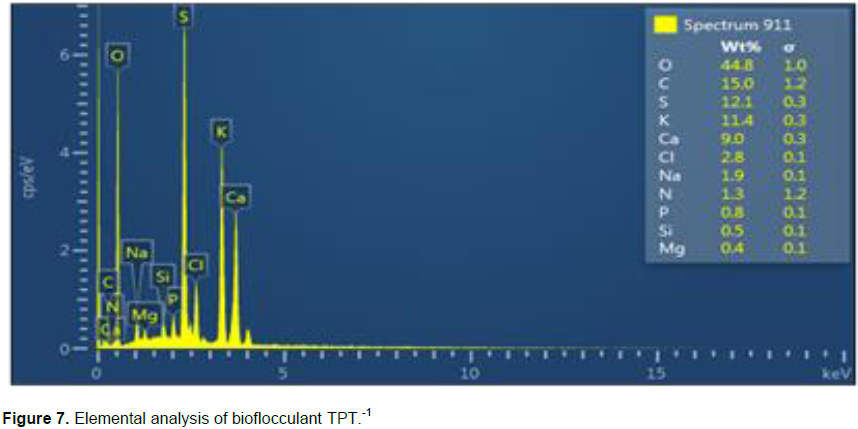
Figure 7 illustrates the elemental analysis of the bioflocculant. The bioflocculant composed of; N (1.3), C (15.0), O (44.8), P (0.8), Ca (9.0), Cl (2.8), Mg (0.4), S (12.1), K (11.4) and Na (1.9), in mass proportion (% w/t).
IR spectrophotometry analysis
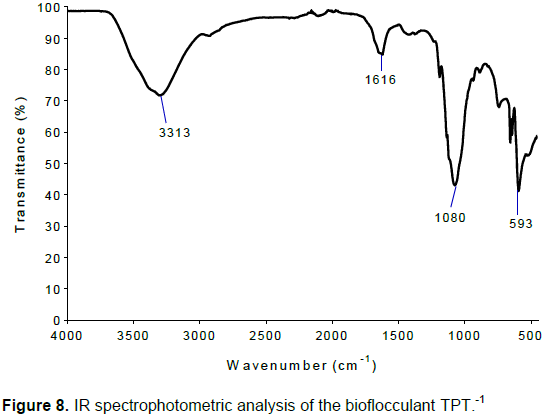
The functional groups of TPT-1 were determined and the results shown in Figure 8. The IR spectrum revealed the presence of different functional groups of the bioflocculant. Hydroxyl (3313 cm-1), carbonyl (1616 cm-1) and amino groups (1080 cm-1) were the main functional groups observed.
Thermal stability of bioflocculant TPT-1
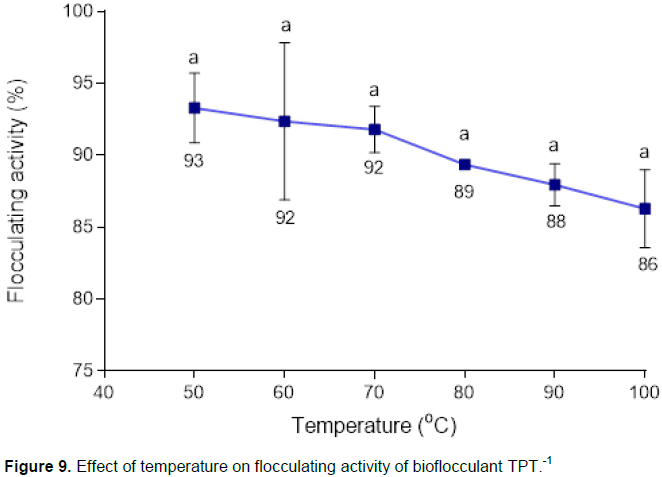
The effect of temperature on flocculating activity of bioflocculant TPT-1 is shown in Figure 9. There was a slight, insignificant drop in flocculating activity with the increase in temperature. TPT-1 also showed high flocculating activity of 86% even at 100°C after an hour of heat exposure.
Pyrolysis property of TPT-1
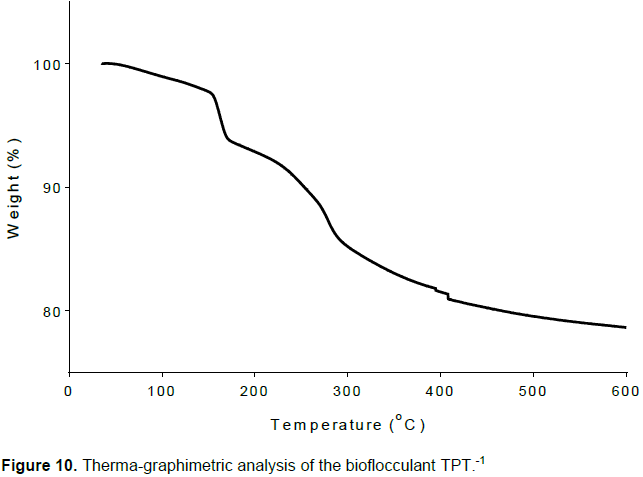
Figure 10 shows the pyrolysis properties of bioflocculant TPT-1. The first degradation was observed at 150°C. The two other degradation temperatures were observed at 270.03 and 402.58°C.
pH stability of bioflocculant TPT-1
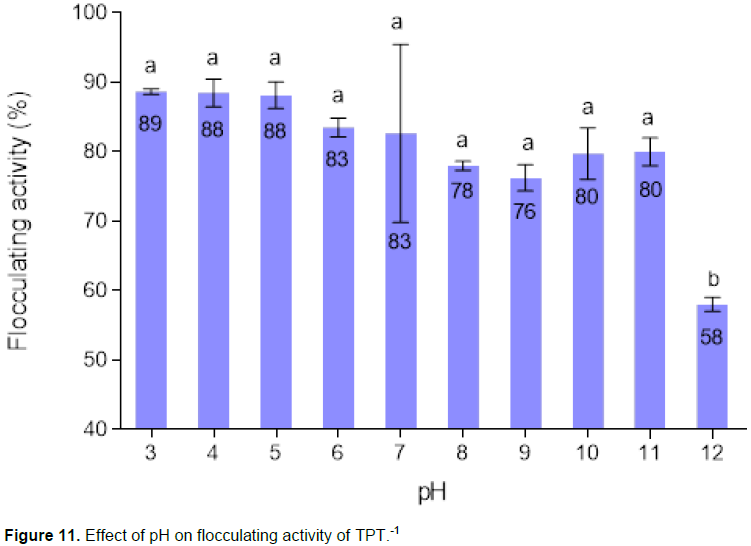
Figure 11 presented the effect of pH on flocculating activity of the bioflocculant. The flocculating activity of TPT-1 was more than 80% in the acidic conditions (pH 3 to 6) and the maximum flocculating activity (89%) was at pH 3.0. pH 12 demonstrated the least flocculating activity of 58%.
Salinity stability of bioflocculant TPT-1
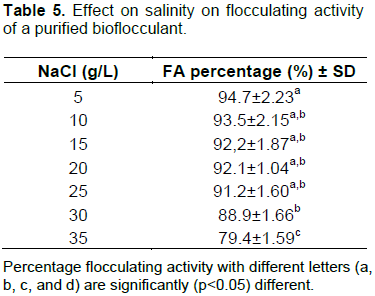
The effect of Na+ concentration on the flocculating activity of bioflocculant TPT-1 is illustrated in Table 5. The flocculating activity of the bioflocculant decreased proportionally with the increase in Na+ concentration. However, TPT-1 maintained high flocculation activity (79.4%) even at the high salinity (35 g/L).
Proposed flocculation mechanism of TPT-1
Table 6 illustrates the zeta potential values of the bioflocculant, Kaolin suspension, Kaolin plus Ba2+ suspension and Kaolin suspension flocculated with Ba2+ and TPT-1. There was an increase in the zeta potential with the addition of Ba2+. Thus, it was suggested that Ba2+ mediated bridge flocculation mechanism between the bioflocculant and Kaolin particles in solution.

DISCUSSION
Biologically, synthesised active bioflocculants are produced in extremely low quantities naturally. This is due to the fact that they are none essential for microbial survival or are present in sufficient amounts to satisfy their primary needs. Low productivity often translates economical unfriendliness especially at industrial level (Barredo, 2005). Thus, to increase bioflocculant production and improve flocculating activity, optimization of cultivation conditions are done.
Naturally, microorganisms coexist in ecological niches and have relationships that have all positive effect on biological adequacy of all interacting species. Microbial species interact with one another and with physic-chemical surroundings in diverse ways, forming convoluted relationships which include symbiosis and synergism (Manahan, 2005). Thus, it have been reported that mixed microbial strains in consortia produce bioflocculants that have better flocculating activity and higher yields than pure strains (Zhu et al., 2004).
The growth kinetics of cultured bacteria and the production of bioflocculant synthesis greatly depended on inoculum size. Small inoculum sizes lengthen the stagnant phase during bioflocculant synthesis while large inoculum sizes cause a niche overlap and thus tend to hinder bioflocculant production (Aljuboori et al., 2014). The optimum inoculum size was 2% (1 ml (v/v)) and gave the highest flocculating activity (93%) (Table1). The desirable inoculum sizes range from 1 to 5% (v/v) (Okoh et al., 2012), and 2% inoculum size was within the preferred range.
Bacterial strains acquire their energy for metabolic processes and reproduction by mediating chemical reactions. The bulk of net energy–yielding or energy consuming metabolic reactions in microorganisms involves change in the oxidation state of carbon or nitrogen elements in substrates (Mahan, 2005). Table 1 showed the effects of various carbon and nitrogen sources on bioflocculant production. Glucose was the most preferred carbon source, yielding flocculation activity of 80.5±9.07%.
Okoh et al, (2012), found identical observations whereby glucose enhanced production of bioflocculant by mixed culture of Streptomyces sp. Gansen and Cellulomonas sp. Okoh. Yeast extract revealed to be the most preferred nitrogen source, resulting in flocculating activity of 97.4±0.57%. Cations enhance flocculation by accelerating bridge formation between colloids and bioflocculants (Lu et al., 2005). Cations increase the initial adsorption of bioflocculants onto colloids by decreasing the negative charges on functional groups of bioflocculants and colloids. Table 1 showed that all divalent cations used, significantly improved flocculation giving the flocculation activities above 80%. Ba2+ was the most profound cation with the highest flocculation activity of 96.2±1.25%.
The cultivating temperature of microorganisms has direct relationship with their metabolism. Maximum enzymatic reactions for bioflocculant production are obtained at the optimum temperature. A lower culture temperature may cause bacteria to hibernate and delay activation of bioflocculant production while the high temperature can denature enzymes and thus limit bioflocculant production. Bacteria in consortium had the maximum flocculating activity (96%) at 30°C (Figure 1). The results resembled those by Zhang et al. (2007), whereby strain BAFRT4, HXCS2, HXTD2, CYGS1 and CYGS4 in consortia had the highest flocculating activity
at at 30°C.
The initial pH of the culture medium affects bioflocculant production. Initial pH determines the electric charge of the microbial cells and the oxidation-reduction potential, which in turn can affect biochemical reactions and adsorption of nutrients. Summarily, initial pH affects the biosynthesis of bioflocculants and bacterial growth (Zhang et al., 2007). The initial pH of 6 was the optimum pH for the fermentation medium, yielding 97% of the flocculating activity (Figure 2). B. pumilus JX860616 and A. faecalis HCB2B might have had symbiosis or synergism relationship, which enabled them to survive and produce bioflocculant within a wide range of pH (Manahan, 2005). The findings were similar to those reported by Zhang et al. (2007), whereby the initial pH of 6 was the best pH for bioflocculant production by strain BAFRT4, HXCS2, HXTD2, CYGS1 and CYGS4 in consortia.
Figure 3 shows that the flocculating activity increased in parallel with cell growth for the first 72h. This suggested that the bioflocculant accretion in the medium during exponential growth phase was produced by biosynthesis and not cell autolysis (Okoh et al., 2012). The flocculating activity decreased slowly after 72h, and it was assumed to be due to cell autolysis and partial decrease in enzymatic reactions. Okoh et al. (2013) also found that the flocculant production by consortium of Methylobacterium sp. and Actinobacterium sp. Mayor was at its peak after 72 h.
Moreover, the initial pH of the medium dropped constantly from the initial pH of 6.0 after 12h to the final pH of 4.3 (Figure 3). The decrease in the initial pH value might be due to the acidic components of the produced bioflocculan (Gomaa, 2012).
Three grams of the purified bioflocculant TPT-1 was obtained from 1 L of fermentation broth of consortium of B. pumilus JX860616 and A. faecalis HCB2 B. Zhang et al. (2007) reported the highest bioflocculant production (15 g/L) ever, from multiple-microorganisms consortium in a litter. The obtained yield was five times lower in quantity. Nevertheless, it was higher than the yields mostly produced by single bacterial isolates (Lin and Harrichurd, 2011).
Solubility is as the result of solvation of polar and charged groups on the surface of bioflocculant molecule (Walker and Wilson, 2005). TPT-1 was soluble in water but insoluble in all other solvents used (Table 2). The presence of hydroxyl groups might have built up strong forces of attraction between bioflocculant TPT-1 make-ups and resulted in relatively rigid crystalline solids – whereby hydrogen bonding could occur. Since these strong forces were not dissociable by organic solvents, the bioflocculant TPT-1 was insoluble in all organic solvents (Patil et al., 2011). The solubility of TPT-1 in aqueous solution was due to the principle that “like dissolve like “(Boyd, 2015). Thus, polar and charged functional groups of TPT-1 were solvated by aqueous molecules. The hydroxyl groups of the bioflocculant have the probability of forming hydrogen bonds with water, thus making TPT-1 soluble and hydrophilic (Patil et al., 2011).
Low dosage or over-dosage of bioflocculant often turns to lower flocculation activity (Cosa et al., 2013). Table 3 shows that the highest flocculating activity (85.03 ± 2.63%) was at the high concentration of 0.8 mg/ml. However, there was no significant difference between 0.2 mg/ml and 0.8 mg/l in flocculating activity. Thus, 0.2 mg/ml have the most preferred dosage size with the flocculating activity of 78.6 ± 3.33%. At a very low concentration of 0.2 mg/ml, the bridging flocculation mechanism of the bioflocculant was effectively enhanced as most Kaolin particles in solution which have enough bioflocculant molecules to bind to. The low concentration implied that the bioflocculant TPT-1 is potentially economical (Okaiyeto et al., 2016). Lu et al. (2005) reported that cations have significant effect on flocculation. Cations are used as coagulant aids in achieving high flocculation activities by neutralizing the negatively charged functional groups of bioflocculants and suspended particles, thereby increasing the adsorption of bioflocculants to the suspended particles (He et al., 2010). The stimulating effect of cations in flocculation is dependent on the concentration of the cations as well as the valence ions (Nwodo et al., 2016). The obtained results affirmed the hypothesis (Table 3). All the divalent cations used (Ca2+, Mn2+ and Ba2+) significantly enhanced flocculation activity, with Ba2+ being the best cation with the flocculation activity of 88.6±4.94%. The results are in agreement with those of Okoh and Ugbenyen. (2014), whereby the flocculating activity of the bioflocculant from the consortium of Cobetia sp. and Bacillus sp. was significantly stimulated by all divalent cations used (Mg2+, Mn2+ and Ca2+) but reduced slightly by monovalent cations (K+ and Li+). The chemical functionalities and structures of bioflocculants affect the flocculating efficiency and flocculating mechanisms (Badireddy et al., 2010; Li-Fan and Cheng, 2010). The SEM images in Figure 4, revealed bioflocculant TPT-1 to be crystal-like in structure (Figure 4a).
The configuration of TPT-1 might be accountable for its flocculation activity. The Kaolin clay particles (Figure 4b) appeared to be fine and evenly scattered and the floc seemed to be clustered together (Figure 4c). The observations were in consistent with the findings of Zhang et al. (2007), whereby the images of the flocculated Kaolin particles appeared aggregated.TPT-1 was Ninhydrin positive, implying that amino acids were one of the components of the bioflocculant (Gomaa, 2012). The zeta potential spectrum showed bioflocculant TPT-1 to be negatively charged (−15.1 mV) (Figure 5).This implied that the nature of bioflocculant TPT-1 is anionic. The net negative charge of the bioflocculant TPT-1 was concluded to be from the carbohydrates and proteins contents, which normally have the negatively charged functional groups (carboxyl groups) (Liu et al., 2015). Proteins usually show absorption peaks in the range of 275 to 280 nm and between 265 to 295 nm, the absorbance often originate from exposure of the tryptophan residue (Lucas et al., 2006).
The UV-vis spectrum in Figure 6, displayed a clear absorption peak at 289 nm, which have distinctive characteristic of protein. There was no characteristic absorption peak observed at 260 nm, implying that bioflocculant TPT-1 had no nucleic acid composition. The total carbohydrates concentration of 83.1% w/w of the TPT-1 was much higher than the total protein concentration (9.7% w/w) (Table 4). The results confirmed the hypothesis that the predominant components of bioflocculants are carbohydrates (More et al., 2014). Thus, it was assumed that carbohydrates were most active components in flocculation process. The elemental state of biomolecules, often consist of carbon, hydrogen, oxygen, nitrogen, sulphur, phosphorus, iron, sodium, calcium and magnesium (Singh and Kapoor, 2010). Figure 7 shows the results obtained from quantitative analysis of different elements in TPT-1. The presence of carbon, oxygen and nitrogen elements further confirmed TPT-1 as a glycoprotein bioflocculant (Devi et al., 2015). The non-sugar components were little, although they may be responsible for flexibility and stability of TPT-1 (Cosa et al., 2013).
The functional moieties of bioflocculant TPT-1 revealed by IR spectrum in Figure 8, showed the presence of strong broad O-H stretch (3313 cm-1), which indicated the presence of alcohol (Cosa et al., 2013). The carbonyl stretching absorption was observed between 1616 cm-1 and the C-N stretch (1080 cm-1), representing aliphatic amine. There was also a strong absorption peak at 593 cm-1, representing presence of halo compound. The revealed functional groups were the major adsorptive forces of TPT-1 and were perceived to have been involved in flocculating activity of the bioflocculant (Wang et al., 2011). The results also suggest that the functional groups were the binding sites for Ba2+, permitting an enhanced flocculation process (Zheng et al., 2008).
Bioflocculant TPT-1 was heat stable and retained high flocculating activity of 86% at 100°C (Figure 9). The thermal stability of TPT-1 was in consistent with the fact that bioflocculants rich in polysaccharides are more thermally stable than those that are mainly proteinous in nature, which are thermally labile (Zaki et al., 2013). The carbonyl and hydroxyl groups of TPT-1, as shown by IR spectrum (Figure 8), might have permitted the formation of hydrogen bonds which might be responsible for thermal stability of TPT-1 (Okoh and Ugbenyen, 2014). Figure 10 illustrated the pyrolysis property of bioflocculant TPT-1.
The first degradation temperature (Td) was observed at 150°C. The higher the carboxyl content the greater the affinity of the bioflocculant to interact with water molecules. The observed initial weight loss between 35 and 150°C was due to loss of moisture content as a result of the levels of carboxyl groups present in bioflocculant TPT-1. The decrease in weight after 150°C was credited to the first degradation of the bioflocculant TPT-1 and the onset decomposition further occurred at 270.03 and 402.58°C, which led to the drastic weight loss. However, the results suggested that bioflocculant TPT-1 is thermostable.
The change in pH may affect the charge status of the functional groups of bioflocculants and the stability of colloidal materials and formation of bigger flocs, consequently affecting the flocculating activity (Zhang and Lin, 1999). TPT-1 was found to be effective at a wide range of pH (pH 3 to 9), with the highest flocculation activity (89%) at pH 3 (Figure 11). However, the decrease in flocculating activity at pH 12, suggested the alkaline degradation of the bioflocculant TPT-1, which could have resulted from changes that include molecular rearrange-ments of the polysaccharide chain fragmentation of the bioflocculant (Patil et al., 2011). It was, thus assumed that the slight decrease in flocculation activity was due to OH- ions which might have inhibited the formation of flocs of TPT-1 and Kaolin particles in solution. At acidic pH, TPT-1 and the Kaolin particles were able to bind together, forming settleable flocs. The pH stability of the bioflocculant TPT-1 at low pH might be due to its chemical composition that is mainly polysaccharides (Li et al., 2008).
The flocculating activity of TPT-1 decreased slightly in proportion to the increase in NaCl concentration (Figure 12). This maybe due to the fact that high concentrations of Na+ did interfere with the establishment of the flocs of bioflocculant TPT-1 and Kaolin particles. Moreover, high concentrations of Na+ may also have denatured some functional moieties of TPT-1 that might be responsible for flocculation process (Aljuboori et al., 2015). However, TPT-1 maintained high flocculation activity (79.4%), even at high Na+ concentration (35 g/L). The saline stability of TPT-1 maybe due to the fact that the bioflocculant was produced by marine bacterial isolates, which survive in marine environment and is characterised by high NaCl concentrations (35 g/L), of which 61.2% is the salt and sodium chloride. Thus, TPT-1 has potential effectiveness in high salinity water bodies. The results were similar to the findings by Li et al. (2008), whereby EPS SM9913 showed high flocculating performance at high salinity of 5 to 100% (Li et al., 2008).
Zeta potential shows the strength of the repulsion force between colloidal particles and the distance which must be overlapped to allow particles to agglomerate together and is measured in millivolts (mV) (Hadgson et al., 2004). The zeta potential values of TPT-1, Kaolin suspension, Kaolin particles plus BaCl2 and flocculated Kaolin particles with BaCl2 and TPT-1 were all negative (Table 6). In Kaolin suspension, the net zeta potential significantly increased with an addition of BaCl2 and bioflocculant. The flocculation process of the negatively charged Kaolin particles by the anionic TPT-1 might also be activated by cationic bridge formation between Kaolin particles and the functional groups of the bioflocculant molecule. Based on these results, it was concluded that Ba2+ stimulated flocculation process by neutralization and stabilization of residual functional groups of TPT-1, forming bridges that aggregated Kaolin particles together. Ba2+ compacted the double layer of Kaolin particles, deteriorated the static repulsive force between Kaolin particles and reduced the distance between Kaolin particles and the functional groups of TPT-1 (He et al., 2010). The observations were similar to those of Song et al. (2014), whereby the bridging mechanism was concluded as the main flocculation mechanism of bioflocculant ETH-2.
CONCLUSION
Bioflocculant TPT-1 is an anionic, heat stable glycoprotein molecule which is best produced when glucose and yeast extract are used as energy sources in optimum fermentation conditions (30°C, 165 rpm, initial pH 6 and 72 h). The produced bioflocculant TPT-1 (3.0 g/L) had good flocculating activity at low concentration (0.2 mg/ml) and performed effectively in a wide range of pH and salinity. The bridging mechanism was involved during flocculation of Kaolin particles. Ba2+ effectively mediated flocculation by neutralizing and stabilizing the TPT-1 residual functional groups. The revealed properties of TPT-1 suggested its potential in industrial applicability. For further studies, the bioflocculant TPT-1 will be applied on wastewater treatment and the molecular methods will also be done so to increase the bioflocculant yield.
CONFLICTS OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
This work was part of a PhD program. The authors acknowledge the research and innovation committee of the University of Zululand for funding this work.
REFERENCES
|
Aljuboori AH, Idris A, Al-joubory HH, Uemura Y, Ibn Abubakar BS (2015). Flocculation behavior and mechanism of bioflocculant produced by Aspergillusflavus. J. Environ. Manage. 150:466-471. |
|
|
Badireddy AR, Chellam S, Gassman PL, Engelhard MH, Lea AS, Rosso KM (2010). Role of extracellular polymeric substances in bioflocculation of activated sludge microorganisms under glucose- controlled conditions. Water Res. 44(15):4505-4516. |
|
|
Barredo JL (2005). Microbial process and products-methods in Biotechnology 18. Humana Press pp. 1-18. |
|
|
Boyd CE (2015). Water quality-An Introduction. 2nd ed. Springer International Publishing. pp. 41-55. |
|
|
Bradford MM (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of proteindye binding. Analys. Biochem. 72:248-254. |
|
|
Carlos DMF, Nancy GL, Glen TD, Lerslie GCP (2011). Biological wastewater treatment. 3rd ed. CRC Press. New York. pp. 38-44, 60-62, 655. |
|
|
Chang WC, Soon AY, In HO, Sang HP (1998). Characterization of an extracellular flocculating substance produced by a planktonic cyanobacterium, Anabaena sp. Biotechnol Lett. 20(12):643-646. |
|
|
Chapelle FH (2001). Ground water Microbiology and Geochemistry. 2nd ed. Willey. Canada. pp. 176-177. |
|
|
Chaplin MF, Kennedy JF (1994). Carbohydrate Analysis, seconded. Oxford University Press, New York. pp. 138-141. |
|
|
Cong-Liang Z, Ya-Nan C, Yan W (2012). Bioflocculant produced from bacteria for decolorization, Cr removal and swine wastewater application. Sustain. Environ. Res. 22(2):129-134. |
|
|
Cosa S, Ugbenyen MA, Mabinya LV, Okoh IA (2013). Characterization of a thermostable polysaccharide bioflocculant produced by Virgibacillus species isolated from Algoa bay. Afr. J. Microbiol. Res. 7(23):2925-2938. |
|
|
Gomaa EZ (2012). Production and characteristics of a heavy metals removing bioflocculant produced by Pseudomonas aeruginosa. Pol. J. Microbiol. 61(4):281-289. |
|
|
Hadgson KG, Freese SD, Nozaic DJ, Borain G (2004) Quantification of factors affecting coagulation of water with cation polymers and laboratory methods for determining these effects.WRC Report. No 1225/1/04. pp. 5-25. |
|
|
Harrington BJ, Raper KB. (1968). Use of a fluorescent brightener to demonstrate cellulose in the cellular slime molds. Appl. Microbiol. 16:106-113. |
|
|
He J, Zou J, Shao Z, Zhang J, Liu Z, Yu Z (2010). Characteristics and flocculating mechanism of a novel bioflocculant HBF-3 produced by deep-sea bacterium mutant Halomonas sp. V3a'. World J. Microbiol Biotechnol. 26:1135-1141. |
|
|
Kay RE, Harris DC, Entenman C (1956). Quantification of the Ninhydrin color reaction as applied to paper chromatography. Arch. Biochem. Biophys. 63:14-25. |
|
|
Kurane R, Takeda K, Suzuki T (1986). Screening for and characteristics of microbial flocculants. Agric. Biol. Chem. 50(9):2301-2307. |
|
|
Li WW, Zhou WZ, Zhang YZ, Wang J, Zhu XB (2008). Flocculation behavior and mechanism of an exopolysaccharide from the deep sea psychrophilic bacterium Pseudoalteromonas sp. SM9913. Bioresour. Technol. 99:6893-6899. |
|
|
Li-Fan L, Cheng W (2010). Characteristics and Culture Conditions of a Bioflocculant Produced by Penicillium sp. Biomed Environ Sci. 23: Liu W, Wang K, Li B, Yuan H, Yang J (2010). Production and characterization of an intracellular bioflocculant by Chryseobacterium daeguense W6 cultured in low nutrition medium. Bioresour. Technol. 101:1044. |
|
|
Liu W, Cong L, Yuan H, Yang J (2015). The mechanism of kaolin clay flocculation by a cation-independent bioflocculant produced by Chryseobacterium daeguense W6. AIMS Environ. Sci. 2(2):169-179. |
|
|
Lu W, Zhang T, Zhang D, Li C, Wen J, Du L (2005). A bioflocculant produced by Enterobacter aerogenes and its use in defecating the trona suspension. Biochem. Eng. J. 27(1):1-7. |
|
|
Lucas LH, Ersoy BA, Kueltzo LA, Joshi SB, Brandau DT, Thyagarajapuram T, Peek LJ, Middaugh R (2006). Probing protein structure and dynamics by second-derivative ultraviolet absorption analysis of cation–π interactions. Protein Sci. 15(10):2228-2243. |
|
|
Manahan S (2005). Environmental Chemistry. 8th ed. CRC Press. pp. 135-165. |
|
|
More TT, Yadav JSS, Yan S, Tyagi RD, Surampalli RY (2014). Extracellular polymeric substances of bacteria and their potential environmental applications. J. Environ. Manage. 144:1-25. |
|
|
Nwodo UU, Makapela B, Okaiyeto K, Ntozonke N, Green E, Mabinya LV, Okoh AI (2016). Assessment of Bacillus pumilus isolated from fresh water milieu for bioflocculant production. Appl. Sci. 6(8):211 |
|
|
Okaiyeto K, Nwodo UU, Mabinya LV, Okoh AI (2013). Characterization of a bioflocculant produced by a consortium of Halomonas sp. Okoh and Micrococcus sp. Leo. Environ. Res. Public Health. 10(10):5097-5110. |
|
|
Okaiyeto K, Nwodo UU, Mabinya LV, Okoli AS, Okoh AI (2015). Characterization of a bioflocculant (MBF-UFH) produced by Bacillus sp. AEMREG7. Int. J. Mol. Sci. 16:12986-13003. |
|
|
Okaiyeto K, Nwodo UU, Okoli SA, Mabinya LV, Okoh AI (2016). Implications for public health demands alternatives to inorganic and synthetic flocculants: bioflocculants as important candidates. Microbiologyopen. 5(2):177-211. |
|
|
Okaiyeto K, Nwodo UU, Okoli AS, Mabinya LV, Okoh AI (2016). Studies on bioflocculant production by Bacillus sp. AEMREG7. Pol. J. Environ. Stud. 25(1):241-250. |
|
|
Okoh A, Ugbenyen A, Cosa S, Mabinya L, Babalola OO, Aghdasi F (2012). Thermostable bacterial bioflocculant produced by Cobetia Spp. isolated from Algoa Bay (South Africa). Int. J. Environ. Res. Public Health 9(6):2108-2120. |
|
|
Okoh AI, Luvuyo N, Nwodo UU, Mabinya LV (2013). Studies on bioflocculant production by a mixed culture of Methylobacterium sp. Obi and Actinobacterium sp. Mayor. BMC Biotechnol. 13(1):1-62. |
|
|
Okoh AI, Ugbenyen AM (2014). Characteristics of a bioflocculant produced by a consortium of Cobetia and Bacillus species and its application in the treatment of wastewaters. Water SA. 40(1):139-144. |
|
|
Patil SV, Pati CD, Salunke BK, Salunkhe RB, Bathe GA, Patil DM (2011). Studies on characterization of bioflocculant exopolysaccharide of Azotobacter indicus and its potential for wastewater treatment. Appl. Biochem. Biotechnol. 163(4):463-472. |
|
|
Salehizadeh H, Shojaosadati SA (2001). Extracellular biopolymeric flocculants Recent trends and biotechnological importance. Biotechnol. Adv. 19:371-385. |
|
|
Serdar S, Faruk K, Orhan Y, Barbara N, Ebru TO, Mehmet AY (2011). Flocculating performances of exopolysaccharides produced by a halophilic bacterial strain cultivated on agro-industrial waste. Bioresour. Technol. 102:1788-1794. |
|
|
Singh US, Kapoor K (2010). Microbial biotechnology. Oxford Book Company. India. pp. 232-233. |
|
|
Song L, Tang W, Li D, Qiao J, Zhao T, Zhao H (2014). Production, characterization, and flocculation mechanism of cation independent, pH tolerant, and thermally stable bioflocculant from Enterobacter sp.ETH-2. PLoS ONE 9(12):e114591. |
|
|
Walker J, Wilson K (2005). Principles and Techniques of Practical Biochemistry. 6th ed. Cambridge University Press. pp. 349-404. |
|
|
Wang L, Ma F, Qu Y, Sun D, Li A, Guo J, Yu B (2011). Characterization of a compound bioflocculant produced by mixed culture of Rhizobium radiobacter F2 and Bacillus sphaeicus F6. World J. Microbiol. Biotechnol. 27:2559-2565. |
|
|
Zaki S, Farag S, Abu Elreesh G, Elkady M, Nosier M, Abd-El D (2011). Characterization of bioflocculants produced by bacteria isolated from crude petroleum oil. Int. J. Environ. Sci. Technol. 8(4):831-840. |
|
|
Zaki SA, Elkady MF, Farag S, Abd-El-Haleem D (2013). Characterization and flocculation properties of a carbohydrate bioflocculant from a newly isolated Bacillus velezensis 40B. J. Environ. Biol. 34:51-58. |
|
|
Zhang H, Lin Z (1999) Microbial flocculant and its application in environmental protection. J. Environ. Sci. 11:1-12. |
|
|
Zhang ZQ, Lin B, Xia SQ, Wang XJ, Yang AM (2007). Production and application of a novel bioflocculant by multiple-microorganism consortia using brewery wastewater as carbon source. J. Environ. Sci.-China. 19(6):667-673. |
|
|
Zheng Y, Ye ZL, Fang XL, Li YH, Cai WM (2008). Production and characteristics of a bioflocculant produced by Bacillus sp. F19. Bioresour. Technol. 99:7686-7691. |
|
|
Zhu YB, Feng M, YangA JX (2004). Screening of complex biofloc-culant producing bacterium and their flocculating mechanism. J. Harbin Inst. Technol. 36(6):759-762. |
|
|
Zhao HJ, Liu HT, Zhou JG (2013). Characterization of a biofiocculant MBF-5 by Klebsiella pneumoniae and its application in Acanthamoeba cysts removal. Bioresour. Technol. 137:226-232. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


