Full Length Research Paper

ABSTRACT
Microalgae can be used as both food and a source of bioactive compounds, such as oils, vitamins and biosurfactants. An important factor contributing to the production costs of bioactive compounds, such as biosurfactants, is the carbon source. One way to decrease production costs is by reducing carbon without a concomitant reduction in productivity. Biosurfactants have a wide range of industrial applications, particularly in the food industry, where they are used as emulsifiers and thickeners. We have investigated the use of cyanobacteria Arthrospira sp. LEB 18 and Synechococcus nidulans LEB 25 and of chlorophytes Chlorella minutissima LEB 108, Chlorella vulgaris LEB 106 and Chlorella homosphaera for the production of biosurfactants using autotrophic and mixotrophic cultivation. The strains were grown in Erlenmeyer photobioreactors containing appropriate media with NaHCO3 as the autotrophic carbon source and glucose or molasses for mixotrophic growth. The results obtained demonstrate the potential of organic carbon sources to stimulate both the growth of microorganisms and biosurfactant production. Furthermore, the data highlight the potential of using molasse, a low-cost byproduct, as an organic substrate for microalgae cultivation.
Key words: Bicarbonate, biosurfactants, glucose, molasse, superficial tension.
INTRODUCTION
The world production of surfactants exceeds 3 million tons per year, nearly all of which are petroleum derivatives, with 70 to 75% of this production used by industrialized countries (Banat et al., 2000). Biosur-factants are biologically produced and composed by complex molecules and encompass a wide variety of chemical structures, such as glycolipids, lipopeptides, lipoproteins, neutral lipids, fatty acids and phospholipids (Desai and Banat, 1997).
Interest in biosurfactants has increased due to their diversity and potential application in areas such as food processing, environmental protection, pharmaceuticals and the recovery of oily residues. The surfactants produced by microorganisms have the advantage of being biodegradable and possess a great deal of specificity (Sundaram and Thakur, 2015). Biossurfactant production uses microalgae because these organisms are major producers of glycolipids, phospholipids and neutral lipids (Rodolfi et al., 2009). Furthermore, these microorganisms can also be a source of biocompounds that have applications in pharmaceutical and food industries, like biopigments and essential fatty acids, direct application of biomass in animal and human feed, biofuel production (H2, biodiesel, bioethanol and biogas), and carbon dioxide biofixation (Morais et al., 2015; Pandey et al., 2014; Bellou et al., 2014 ).
A major advantage of cultivating microalgae for biosurfactant production is that many of these microorganisms fall into the Generally Recognized As Safe (GRAS) category. Such certified organisms have no risk of toxicity or pathogenicity and can be used for applications in the food and pharmaceutical industries (Soccol et al., 2013).
Commercial biosurfactant production is limited due to the high costs involved, particularly with respect to culture media. The use of cheaper substrates, such as molasses and glucose, may reduce the cost factor and make production economically viable. Mixotrophic microalgae culture can significantly enhance the growth of microalgae, resulting in cell densities three to ten times higher than those obtained in autotrophic culture (Bhatnagar et al., 2011). For Arthrospira microorganisms, the use of molasse as the carbon source is the most influential factor for maximizing biomass concentration and specific growth rate (Andrade and Costa, 2007). Furthermore, an organic substrate in the culture medium can reduce nocturnal loss of biomass because cellular energy demand can be supplied by respiration (Torzillo et al., 1991).
We investigated the cyanobacteria strains Arthrospira sp. LEB 18 and Synechococcus nidulans LEB 25 and the chlorophyte strains Chlorella minutissima LEB 108, Chlorella vulgaris LEB 106 and Chlorella homosphaera for the production of biosurfactants in autotrophic and mixotrophic cultivation.
MATERIALS AND METHODS
Mixotrophic cultivation of Arthrospira sp LEB 18
Maintenance and growth of Arthrospira.sp. strain LEB 18 (Morais et al., 2008) was performed over 10 days in Zarrouk medium (Zarrouk, 1966) and was supplemented during the dark period with a total of 1, 3, 5, 7 or 9 g.L-1 of glucose or molasses (Indumel, Brazil) added to the cultures at a rate of 10% per day. The cultures were carried out in 2 L Erlenmeyer photobioreactors maintained at 30°C and under a light intensity of 41.6 µmol m-2 s-1 generated by 40 W fluorescent lamps in a 12 h photoperiod. The cultures were stirred by injecting sterile air with a specific flow rate with volume of air per volume of medium per minute (vvm) of 0.5 (Costa et al. 2000). The initial LEB 18 concentration was 0.15 g L-1 (Radmann et al. 2007). The glucose concentration was analyzed using the glucose-oxidase enzymatic method (Laborlab, Campinas, Brazil). The culture analysis were performed in duplicate.
The organisms used in this study were the cyanobacteria Arthrospira sp. LEB 18 and S. nidulans LEB 25 and the chlorophytes C. minutissima LEB 108, C. vulgaris LEB 106 and C. homosphaera. The cyanobacteria were maintained and grown in Zarrouk medium (Zarrouk, 1966), and the chlorophytes were maintained and grown in BG-11 medium (Rippka et al., 1979). Autotrophic growth used a total of 16.8 g L-1 sodium bicarbonate as the carbon source (Chen et al., 1996). Mixotrophic growth was conducted over 20 days using glucose as total carbon source of 5 g.L-1 (selected as described in the previous section), which was added to the cultures during the dark period at a rate of 5% per day. Glucose was added incrementally because adding the total amount at the beginning of the experiment caused precipitation and, thus, reduced nutrient availability. Before glucose addition the residual glucose concentration (see below) was measured in the culture medium to determine whether the previous aliquot had been utilized during the light phase. Cultures were carried out in 2 L Erlenmeyer photobioreactors maintained at 30ºC and were exposed to a light intensity of 41.6 µmol m-2 s-1 generated by 40 W fluorescent lamps with 12 h photoperiod.
The cultures were stirred by injecting sterile air with a specific flow rate of 0.5 vvm (Costa et al., 2000). The initial biomass concentration was 0.15 g.L-1 (Radmann et al. 2007). The glucose concentration was analyzed using the glucose-oxidase enzymatic method (Laborlab, Campinas/Brazil). All tests and analysis were performed in duplicate.
The increase in biomass was monitored daily by measuring the optical density of the cultures at 670 nm using a spectrophotometer (FEMTO 700 Plus) and a previously constructed standard curve relating dry weight and optical density. The following parameters were evaluated: maximum biomass concentration (Xmax, g L-1); maximum yield, (Pmax, g L-1 d-1), obtained from P = (Xt - X0)/(t - t0), where Xt is the biomass concentration (g L-1) at time t (d) and X 0 the biomass concentration (g L-1) at time t0 (d) (Schmidell et al., 2001); and the maximum specific growth rate (μmax, d-1) by exponential regression of the logarithmic growth phase (Bailey and Ollis, 1996).
Culture samples were taken every two days and sonicated for 15 min in an ultrasonic bath to break the cell walls, and the surface tension of the sonicate was immediately evaluated using the ring method using a digital tensiometer (Kruss Processor Tensiometer K-6, Germany) and the sample in contact with air (Rodrigues et al., 2006). The results were expressed as minimum surface tension over time (TSmin, mN m-1).
Statistical analysis
All reported values represent the average value of the analysis of three replicates. An ANOVA was performed followed by Tukey's test (p<0.05) for mean comparison using the Statistica 8.0 software for Windows (Statsoft, Tulsa, OK, USA).
RESULTS AND DISCUSSION
All cultures showed cell growth over 10 days except for assays with added molasses concentrations of 7 and 9 g L-1, which showed cell death in 3 days. In experiments measuring cell growth curves with glucose, adaptation was not observed, while the cultures with molasses spent approximately three days from adapt to the conditions subjected to the microorganism. In the dark phase of photosynthesis, the microorganisms consume their own energy source, reducing growth rate and, consequently, cell concentration, and recovering again during the light period phase (Nelson and Cox, 2011). However, comparison of the spectrophotometric readings in the dark and light periods revealed that there was no decrease in cell growth in the dark phase because of the addition of organic carbon sources that maintained the growth rate (Figure 1).
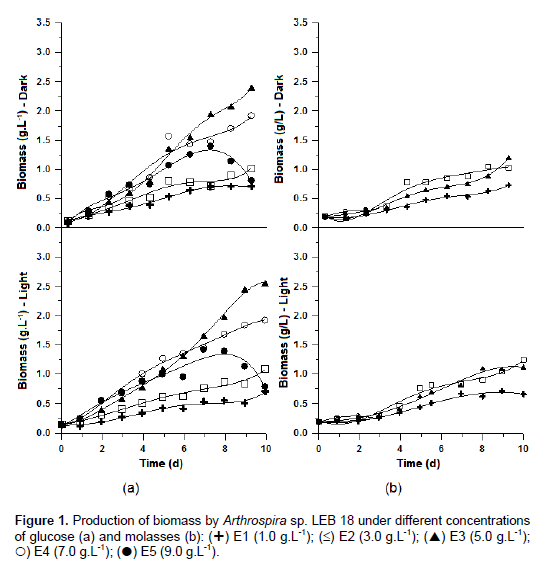
Chen and Zhang (1997) have reported that the cell growth of mixotrophic cultures is limited by low or high concentrations of organic carbon. High concentrations of carbon can induce cellular stress as a result of excess nutrients in the culture medium. At low concentrations, there was growth restriction due to the shortage of an organic carbon source. This phenomenon was observed in the tests performed with the addition of glucose and molasse.
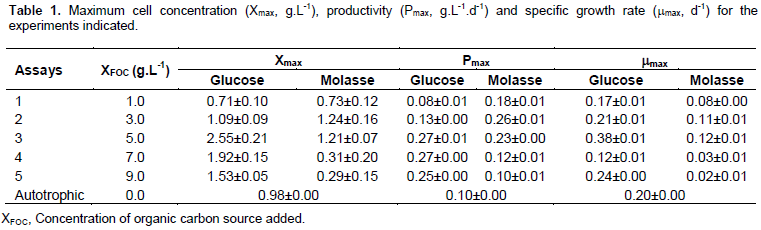
There was an increase in biomass that was directly proportional to the concentration of the carbon source to 5 g L-1, thereafter, a decrease in cell specific growth rate and productivity was observed using glucose. In cultures with molasse, the largest cell concentration achieved was 3 g L-1 of organic source. In trials where higher cell concentrations were obtained for both molasse and glucose, there was also maximum productivity (0.27 and 0.26 g L-1 d-1, respectively). Compared with autotrophic culture (0.10 g L-1d-1), it was observed that the addition of organic source to crops increased significantly (p <0.001) maximum productivity.
Glucose addition increased the maximum specific growth rate 1.9 times during mixotrophic LEB 18 cultivation compared with the autotrophic. In assays using molasses, the maximum growth rate reached in the mixotrophic assay was lower than autotrophic because, in general, this parameter was attained in the exponential growth phase. The growth curves in Figure 1 show that in cultures with molasses, the exponential cell growth phase was not achieved at the end of the experiment for any of the carbon source concentrations. The pH of crops remained between 9.5 and 10.5 which, according to Pelizer et al. (2003), is the optimal range for Arthrospira growth.
Microalgae selection for biosurfactants production
According Torzillo et al. (1991), during the dark phase, biomass is reduced because of cellular energy demand, which is supplied by the endogenous microalgal cell reserves formed during the light phase. In cultures where an organic carbon source was added nightly, biomass loss can be minimized during the dark phase, generating higher cell density compared with the autotrophic culture, as observed in the growth curves in Figure 2.
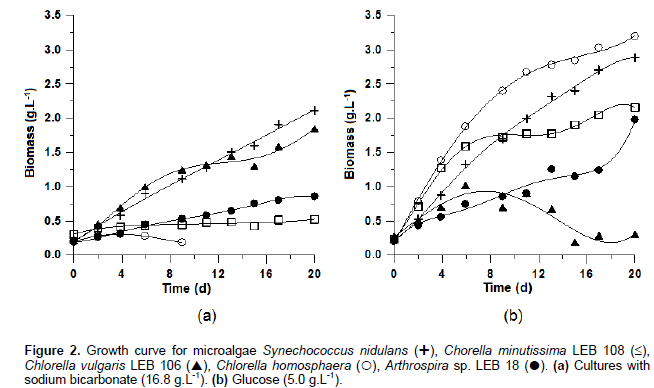
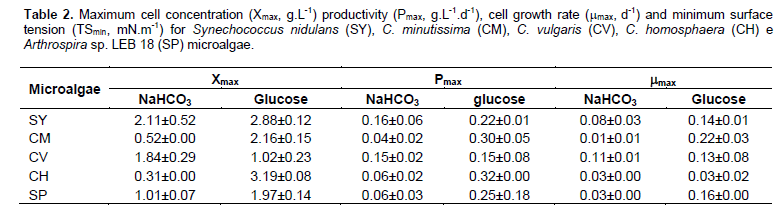
The increase in cell concentration in the mixotrophic condition was observed in the LEB 106 culture, which showed a stationary phase from 5 days of cultivation with glucose addition. In this growth step, the microalgae reached the maximum cell concentration in the culture with organic substrate (1.02 g L-1) (Table 1). The stationary phase of cell growth, which stabilizes the cell concentration, can occur because of a lack of nutrients in the culture medium or due to consumption by microorganisms during development. At this stage, the production of metabolites occurs, such as carbohydrates and lipids; these metabolites are used by the end of this phase as a form to obtain energy until the beginning of cell decline (Schimidell, 2001). In this experiment, the addition of glucose caused an increase in the maximum specific growth rate (0.13 d-1) compared with the autotrophic culture (0.11 d-1), quickly consuming the carbon provided in the culture medium and affecting the stability of cell concentration more rapidly.
The microalgae studied showed better kinetic parameters of growth in crops that were mixotrophically cultured. Chojnacka and Noworyta (2004) observed a similar pattern with higher specific growth speeds for mixotrophic cultivation compared with autotrophic. Due to the different cellular and genetic characteristics of each of the strains studied, the addition of glucose influenced growth kinetic parameters differently. The largest maximum cell concentration during glucose cultivation was obtained for C. homosphaera (3.19 g L-1); this experiment achieved the highest maximum mobile productivity (0.32 g L-1 d-1) (Table 2). However, increasing cell concentration did not have a proportional relationship to biosurfactant production. In general, the improvement of a target compound is produced by the addition of substrates that create stress conditions for the cells. This stress results in a microalgal metabolism deviation, leading to the production of specific compounds, usually energetic compounds such as lipids and carbohydrates that compose the biosurfactants.
Compared with experiments performed earlier (Figure 1), the LEB 18 exhibited lower growth with the same concentration of glucose (5.0 g L-1) because of the different conditions when substrate was added. In the previous experiment lasting 10 days, glucose was added daily at a higher concentration, while in the 20 days experiment, the daily concentration were lower for same added amount of glucose in the same experiments. The addition of substrate occurred daily because adding the total concentration (5.0 g L-1) at the beginning of the cultivation, glucose precipitation occurred, reducing nutrient availability for microalgae. Every day, before the addition of glucose to the cultures, a concentration analysis was performed in the same medium, verifying that it was totally consumed by the microalgae.
Biosurfactant quality is inverse to its surface tension: the smaller the surfactant, the more efficient the surfactant. Carbon source plays an important role in biosurfactant production, influencing the synthesis of induction or repression. According to Cameotra and Makkar (1998), the carbon sources used for the production of biosurfactants can be divided into carbohydrates, hydrocarbons, and vegetable oils and may determine the specificity of the product. According to Cavalero and Cooper (2003) and Hommel et al. (1994), water soluble substrates such as glucose are primarily used in cell metabolism for synthesis of the hydrophilic portion of the biosurfactant molecule, while the lipophilic substrates are used for the production of the lipophilic portion.
Surface tension reduction was found for all microalgae studied, occurring during biosurfactant production in mixotrophic and autotrophic experiment, with an increasing concentration of this product during cultivation time. Among the cultures with added glucose, LEB 18 had the highest reduction in surface tension: 38% from the initial surface tension, reaching 43 mN.m-1 (Figure 3). S. nidulans presented an end surface tension value that was similar to Arthrospira (44.0 mN.m-1), with a reduction of 35% compared to baseline.
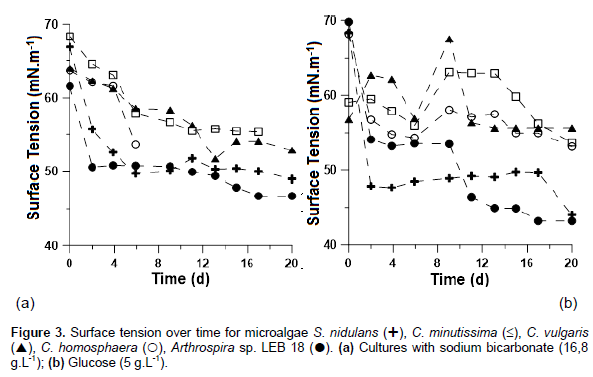
CONCLUSION
In the study of organic carbon sources for the mixotrophic cultivation of Arthrospira sp. LEB 18, the best growth was observed in culture using glucose as an organic carbon source at a concentration of 5 g L-1. Under these conditions, the maximum cell concentration was 2.6 times higher compared with autotrophic cultivation. When evaluating the culture of different microalgae species for biosurfactant production, a reduction in surface tension was observed in all microalgae studied, with an increased concentration of this product during cultivation time. LEB 18 had the highest reduction in surface tension: 38% from the initial, reaching 43 mN.m-1. These results demonstrate the potential of organic carbon sources to stimulate both the growth of microorganisms and biosurfactant production. Furthermore, the data highlight the potential of molasses, a low-cost byproduct, as an organic substrate for microalgae cultivation.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
The authors would like to thank the Brazilian Electric Power Company (ELETROBRAS), the Thermal Generation of Electric Energy Company (CGTEE), Publication Support Program of Academic Production (PROPESP/FURG -2015), and Ministry of Science, Technology and Innovation (MCTI) for their financial support of this study.
REFERENCES
|
Andrade MR, Costa JAV (2007). Mixotrophic cultivation of Microalga Spirulina platensis using molasses as organic substrate. Aquaculture 264:130-134. Crossref |
||||
| Bailey JE, Ollis DF (1996). Biochemical Engineering Fundamentals, 2ª ed. Singapore, McGraw-Hill. | ||||
|
Banat IN, Makkar RS, Cameotra SS (2000). Potential commercial applications of microbial surfactants. Appl. Microbiol. Biotechnol. 53:495-508. Crossref |
||||
|
Bellou S, Baeshen MN, Elazzazy AM, Aggeli D, Sayegh F, Aggelis G (2014). Microalgal lipids biochemistry and biotechnological perspectives. Biotechnol. Adv. 32:1476-1493. Crossref |
||||
|
Bhatnagar A, Chinnasamy S, Singh M, Das KC (2011). Renewable biomass production by mixotrophic algae in the presence of various carbon sources and wastewaters. Appl. Energy. 88:3425-3431. Crossref |
||||
|
Cameotra SS, Makkar RS (1998). Synthesis of biosurfactants in extreme conditions. Appl. Microbiol. Biotechnol. 50:520-529. Crossref |
||||
|
Cavalero DA, Cooper DG (2003). The effect of medium composition on the structure and physical state of sophorolipids produced by Candida bombicola ATCC22214. J. Biotechnol. 103:31-41. Crossref |
||||
|
Chen F, Zhang Y, Guo S (1996). Growth and phycocyanin formation of Spirulina platensis in photoheterotrophic culture. Biotchnol. Letters 18:603-608. Crossref |
||||
|
Chojnacka K, Noworyta A (2004). Evaluation of Spirulina sp. Growth in photoautotrophic, heterotrophic and mixotrophic cultures. Enzyme Microb. Technol. 34:461-465. Crossref |
||||
|
Costa JAV, Linde GA, Atala DIP, Mibielli GM, Arrieche L, Krüger RT (2000). Modelling of growth conditions for cyanobacterium Spirulina platensis in microcosms. World J. Microbiol. Biotechnol. 16:15-18. Crossref |
||||
| Desai JD, Banat IM (1997). Microbial production of surfactants and their commercial potential. Microbiol. Mol. Ver. 61:47-64. | ||||
|
Hommel RK, Weber L, Wiss A, Haferburg D, Kleber HP (1994). Production of sophorose lipids by Candida (Torulopsis) apicola grown on glucose. J. Biotechnol. 33:147-155. Crossref |
||||
|
Morais MG, Reichert CC, Dalcanton F, Durante AJ, Marins LFF, COSTA JAV (2008). Isolation and characterization of a new Arthrospira strain. Z. Naturforsch. 63:144-150. Crossref |
||||
|
Morais MG, Vaz BS, Morais EG, Costa JAV (2015). Biologically Active Metabolites Synthesized by Microalgae. BioMed Res. Int. 2015: 1-15. Crossref |
||||
| Nelson DL, Cox MM (2011). Princípios de Bioquimica de Lehninger, 5 ed, Sarvier. | ||||
| Pandey A, Lee DJ, Chisti Y, Soccol CR (2014). Biofuels from algae, United States, Elsevier. | ||||
|
Pelizer LH, Danesi ED, Rangel CO, Sassano CEN, Carvalho JCM, Sato S, Moraes IO (2003). Influence of inoculum age and concentration in Spirulina platensis cultivation. J. Food Eng. 56: 371-375. Crossref |
||||
|
Radmann EM, Reinehr CO, Costa JAV (2007). Otimization of the repeated batch cultivation of microalga Spirulina platensis in open raceway ponds. Aquaculture. 265:118-126. Crossref |
||||
| Rippka R, Deruelles J, Waterbury JW, Herdman M, Stanier RG (1979). Genetic assignments, strain histories and properties of pure cultures of Cyanobacteria. J. Gen. Microbiol. 111:1-61. | ||||
|
Rodolfi L, Zittelli GC, Bassi N, Padovani G, Biondi N, Bonini G, Tredici MR (2009). Microalgae for Oil: Strain Selection, Induction of Lipid Synthesis and Outdoor Mass Cultivation in a Low-Cost Photobioreactor. Biotechnol. Bioeng. 102:100-112. Crossref |
||||
|
Rodrigues L, Moldes A, Teixeira J, Oliveira R (2006). Kinetic study of fermentative biosurfactant production by Lactobacillus strains. Bioch. Eng. J. 28:109-116. Crossref |
||||
| Schimidell W, Lima AU, Aquarone E, Borzani W (2001). Biotecnologia Industrial. v. 2, São Paulo, Edgard Blücher LTDA. | ||||
|
Soccol CR, Pandey A, Larroche C (2013). Fermentation Processes Engineering in the Food Industry, CRC Press. Crossref |
||||
|
Sundaram S, Thakur IS (2015). Biosurfactant production by a CO2 sequestering Bacillus sp. strain ISTS2. Bioresour. Technol. 188:247-250. Crossref |
||||
|
Torzillo G, Sacchi A, Materasi R (1991). Temperature as an important factor affecting productivity and night biomass loss in Spirulina platensis grown outdoors in tubular photobioreactors. Bioresour. Technol. 38:95-100. Crossref |
||||
| Zarrouk C (1966). Contribuition a Letude Dune Cyanophycee, Influence de Divers Facteurs physiques et Chimiques sur la Croissance et photosynthese de Spirulina maxima geitler. Ph.D. Thesis, University of Paris. | ||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


