ABSTRACT
Solanum lycocarpum St. Hil. is an endemic plant of the Brazilian tropical savannah (cerrado) that is capable of growing on acidic and nutrient-poor land, an ability which attracts attention to its rhizospheric microbiota, including plant growth-promoting rhizobacteria (PGPR). In this work, 131 bacterial strains were isolated from rhizosphere samples of S. lycocarpum and were tested in vitro for direct mechanisms of plant growth promotion (biological nitrogen fixation, phosphate solubilization and indolic compounds production) and enzyme activities. The 26 most promising isolates selected from the previous tests were used to continue the screening. Ten of these isolates showed antifungal activity against fourteen phytopathogenic fungi and twelve isolates showed antimicrobial activity against at least one of the three clinical pathogens evaluated. Seven of the 26 isolates were identified at random as belonging to the genera Bacillus, Burkholderia and Microbacterium, are PGPR and have potential to participate in more in-depth research aimed at the development of bio-inputs, especially the Bacillus strains.
Key words: Biological control, biotechnological potential, cerrado, plant growth-promoting rhizobacteria (PGPR), phytopathogenic fungi, Rhizosphere, Solanum lycocarpum St. Hil.
The Brazilian tropical savannah (cerrado) biome is a mosaic of savannah, forests and grasslands with high species richness. This biome comprises approximately 2 million hectares or 24% of Brazil’s land surface and is surpassed only by the Amazonian forest. The cerrado hosts approximately 6,000 vascular plant species, featuring the highest floral biodiversity among the world's savannas. The cerrado also has a high diversity of microorganisms. Much of the biodiversity of this microbiome is not yet known, which makes it one of the 25 most important terrestrial biodiversity hotspots on the planet (Oliveira-Filho and Ratter, 2002; Sousa et al., 2017).
Native plants are interesting for the study of bioactive compounds since they are adapted to the adverse conditions of their biome, such as aluminum-toxic acidic soil with few nutrients, water stress, exposure to UV radiation and attacks by herbivores and phytopathogens (Bailão et al., 2015). Among the plants in the cerrado, Solanum lycocarpum St. Hil is an important species described in the literature and is popularly known as “lobeira”. The fruit of S. lycocarpum is the basic food of the endangered maned wolf (Chrysocyon brachyurus) (Aragona and Setz, 2001). This plant also attracts interest for its medicinal properties and is used to treat hepatitis, asthma, flu and colds (Araújo et al., 2010).
Plants live in association with soil microbes (bacteria and fungi) during their growth. Free-living soil microorganisms that inhabit the rhizosphere of many plant species have diverse beneficial effects on the host plants through different mechanisms; these microorganisms are generally referred to as plant growth-promoting rhizobacteria (PGPR). In return, the plant's roots release exudates containing nutrients and compounds that can be used by PGPR for their development (Kloepper and Schroth, 1978). PGPR and their interactions with plants have been documented and used as important tools to sustainably manage agriculture. PGPR boost plant growth through various mechanisms, including phosphate solubilization (PS), biological nitrogen fixation (BNF) and production of indole-3-acetic acid (IAA) (Glick, 2012). In addition to their environmental benefits, biofertilizers formulated with PGPR have increasing economic potential. Global projections for the biological market point to an annual growth of 12.7% by 2022, estimated at US $ 11.35 billion. In Brazil, the estimates from the Brazilian Association of Control Companies (ABCBio) appear promising, with an expected annual growth ranging between 15% and 20% for the coming years (Calvo et al., 2014). PGPR may also act in the biological control of phytopathogens by producing metabolites. For example, fungi belonging to the genera Phytophthora and Fusarium, regarded as the most calamitous crop pathogens in the world, are inhibited by PGPR (Ramadan et al., 2016).
PGPR are interesting subjects for the study of new enzymes and antibiotics due to the survival strategies they develop to compete with other microorganisms in the rhizosphere, such as the production of bioactive secondary metabolites (Raaijmakers and Mazzola, 2012) and lytic enzymes. In enzymatic production processes, microorganisms have advantages over other sources due to their low cost and susceptibility to genetic manipulation (Castro et al., 2014). There is strong interest in microbial enzymes such as proteases, pectinases, cellulases, lipases, esterases and amylases in industry sectors. These sectors include food processing, detergent and textile manufacturing, medical therapy and molecular biology, agricultural and pharmaceutical research (Carrim et al., 2006; Ferreira-Filho et al., 2012; Quecine et al., 2014).
Studies on plant-microbe interactions are necessary to preserve the plant richness and bioprospecting of microorganisms associated with plants in the cerrado. Although this topic is currently underexplored, it can generate beneficial bioproducts for humanity. Therefore, this study aimed to isolate, characterize and select cultivable rhizobacteria associated with Solanum lycocarpum St. Hil. with potential applications in agriculture, medicine and industry.
Bacterial isolation
Three rhizospheric soil samples of approximately 50 g each were collected in March 2018 from three S. lycocarpum St. Hil. plants (n = 9) located in a fragment of cerrado vegetation (S 21° 58’/WO 47° 52’) on the campus of the Federal University of São Carlos, São Paulo state, Brazil. For soil sample collection, 5 cm of topsoil was removed and 20 to 25 cm of the soil layer where roots were concentrated was collected. Collected samples were stored in sterilized plastic bags and transported to the Laboratory of Microbiology and Biomolecules (LaMiB®/UFSCar) for bacterial isolation.
The roots were agitated to remove excess soil, the three samples from each plant were mixed and approximately 50 g of rhizospheric soil were obtained per plant. Each mix was added to 90 ml of phosphate buffered saline (PBS) and shaken for 1 h at 150 rpm. The soil-PBS solution obtained was serially diluted and tryptic soy agar (TSA) (Kasvi, Sao Jose dos Pinhais, Brazil) medium supplemented with benomyl (50 μg/ml) to inhibit fungal growth was used for isolation with 10−4 to 10−5 serial dilutions. The plates were incubated at 28°C for 72 h and colonies were collected after 24 h and 72 h to obtain a bacteria pool with greater diversity of metabolism and growth rates and these colonies were purified by repeated plating at least three times on TSA (Schmidt and Belser, 1983). A total of 131 bacterial strains were isolated and inoculated into liquid tryptic soy broth (TSB) (Kasvi, Sao Jose dos Pinhais, Brazil) medium supplemented with glycerol (15% final concentration) and stored at −80°C for future experiments. The strains received codes beginning with the letters "A" or "T" which meant, respectively, picked up after 24 or 72 h of isolation; a number from 1 to 3, related to S. lycocarpum specimen from which it was isolated; the letters "LRZ", indicating that it was isolated from the rhizosphere; finally, sequential numbers for identification.
Growth capacity of isolates in nitrogen-free culture medium
The ability to fix atmospheric nitrogen by the bacteria was evaluated by growing the strains on the semisolid nitrogen-free medium (NFb), prepared according to Dobereiner et al. (1995) and Cattelan et al. (1999). Glass tubes were filled with 3 ml of the NFb medium and after solidification the microorganisms were inoculated in the tube. The test was performed
by triplicate. After 96 h of incubation at 28°C in the dark, the formation of a white growth film near the surface of the tubes indicated a positive result.
Burkholderia ambifaria strain RZ2MS16, was used as a positive control (Batista et al., 2018; Fukuda et al., 2021). The strains that showed positive results in this stage were re-inoculated to confirm the result.
Indolic compound production
Indolic compound production was evaluated using the colorimetric method (Bric et al., 1991) with modifications to make it quantitative. The cultures were resuspended in sterile distilled water at an optical density of 0.1 at 550 nm (OD550 = 1.0 ± 0.05). Then, 100 μl of each solution was inoculated in separate tubes containing 5 mL of liquid 10% TSB (w/v) with 50 μg/ml L-tryptophan. The tubes were incubated for 72 h at 28 ± 2°C and were then centrifuged at 12,000 rpm for 5 min. Subsequently, the IAA concentration in each culture’s supernatant was estimated using Salkowski reagent. The measurements were estimated with a standard curve, which was calculated based on known doses of the hormone (Sigma) at the following concentrations: 1, 5, 25, 50, 75, 100, 125, 150, 175 and 200 μg/ml. The reddish-pink color of the samples indicated the production of indolic compounds. The test was performed by triplicate and the measurements were averaged.
Phosphate solubilization
Qualitative and quantitative assays for phosphate solubilization were performed and measured in insoluble phosphate-containing solid medium (Berraquero et al., 1976). The presence of a clear halo around a colony indicated phosphate solubilization. The solubilization index (SI) values of the isolates that showed this capacity were then calculated according to the ratio of the diameter of the solubilization halo to the diameter of the colony halo and they were classified as having low solubilization potential (SI less than 2), medium solubilization potential (SI between 2 and 3) or high solubilization potential (SI greater than 3). Experiments were conducted in a completely randomized design with two replications per isolate in the quantitative test and four replicates per isolate in the qualitative test, in which the SI was calculated by averaging the values from these replications.
Enzymatic activity
Amylolytic activity was measured in solid agar (Hankin and Anagnostakis, 1975). Isolates were inoculated in 5% TSA medium containing 1% soluble starch. After bacterial growth, 5 ml of a 1% iodine solution was added to each plate, allowing visualization of clear halos around colonies.
A specific medium was used to evaluate lipolytic activity (Sierra, 1957). Previously sterilized Tween 80 or Tween 20 was added to the sterilized culture medium to a final concentration of 1% (w/v) to evaluate bacterial esterase and lipase activities, respectively. The presence of a halo was considered indicative of enzymatic activity.
Pectinolytic activity was determined after growing bacterial isolates on M9 medium, substituting glucose for 1% (w/v) pectin and adding 0.5% yeast extract (Miller, 1972). After bacterial growth, 10 mL of Lugol’s solution was added, followed by a wash with water. The presence of a halo around a colony indicated pectinolytic activity. For pectate lyase activity, final pH of the M9 medium was adjusted to pH 8.0 with 5 M NaOH and for polygalacturonase was adjusted to pH 5.0 using 1 M HCl.
We used a culture medium containing skim milk to determine proteolytic activity (Zarnowski et al., 2007). The formation of a halo around a colony was considered indicative of proteolytic activity. The cellulolytic activity of the isolates was also evaluated (Teather and Wood, 1982). The isolates were grown on M9 medium containing 0.5% yeast extract and 1% carboxymethylcellulose (CMC). After bacterial growth, 10 ml of Congo red dye (1%) was added and the plates were washed with NaCl (5 M). The presence of a colorless halo around a colony indicated enzymatic activity.
Selection of promising PGPR
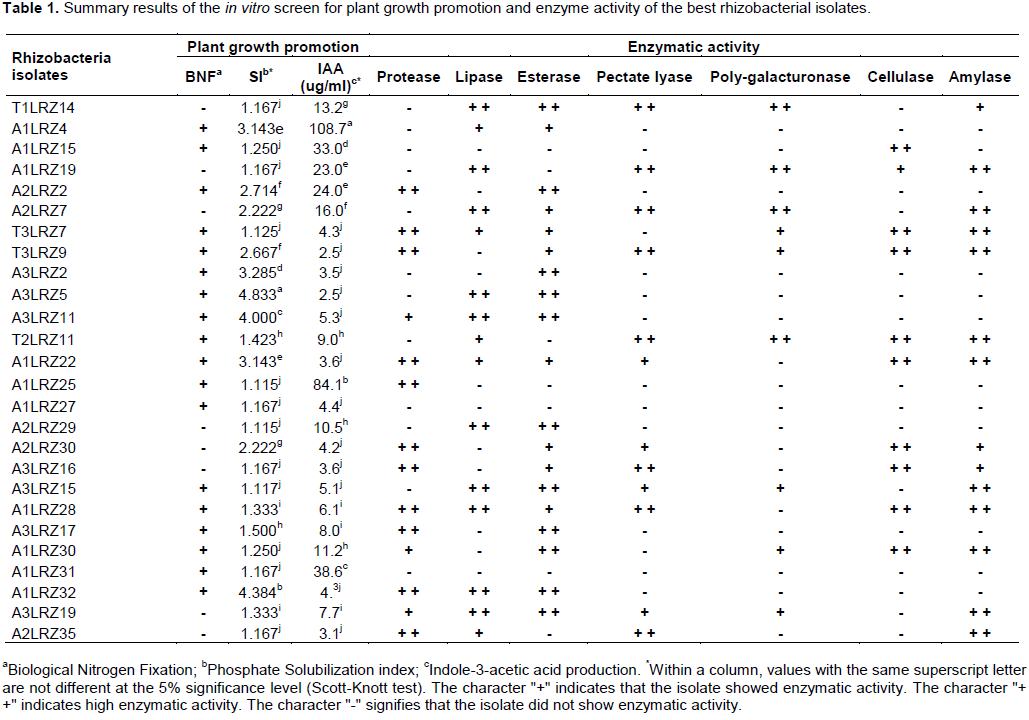
To narrow the screening and select the most promising PGPR from a pool, following assays to determine the direct mechanisms and enzymatic activity, isolates were selected that: presented positive results by more than one direct mechanism, showed SI higher than 1.1, produced indolic compounds and showed more than one type of enzymatic activity. Following this selection, 26 isolates were selected for evaluation in the following tests.
Assessment of antifungal activity
Antifungal activity was evaluated in two parts, visual agar plate and plate pairing assays (Chang et al., 1999). The potato dextrose agar (PDA) plates were divided into quadrants and the 26 selected isolates were inoculated punctually, one isolate per quadrant, 3 cm from the center of the plate. A fungal phytopathogen was point inoculated at the center of each plate. The control plate was only point inoculated with the fungal phytopathogen. The plates were sealed with parafilm and incubated at 28 ± 2°C until the mycelium reached the edge of the control plate (visual agar plate assay). The test was conducted by duplicate.
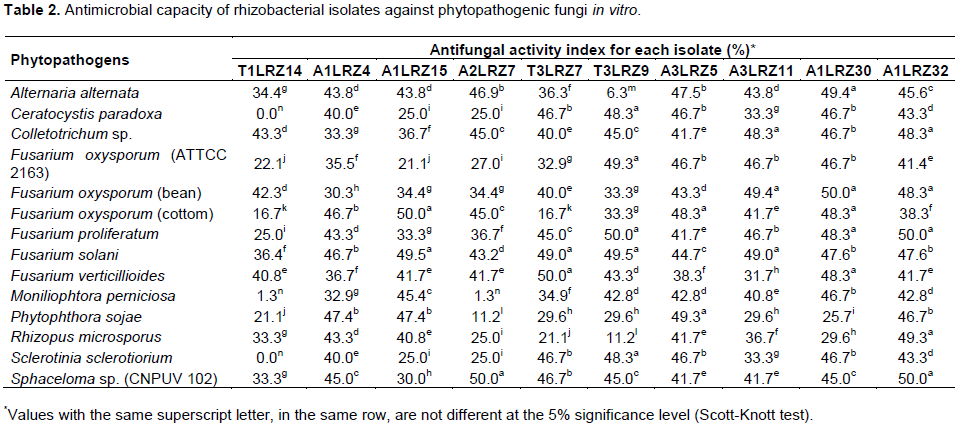
The ten isolates that showed antifungal activity in the visual assay were streaked on PDA medium and the fungal phytopathogen was inoculated punctually at 1 cm from the opposite end from which the bacterial inoculum was streaked (plate pairing assay). The control plate was point inoculated with the fungal phytopathogen. The plates were sealed with parafilm and incubated at 28 ± 2°C until the mycelium reached the edge of the control plate. Antifungal activity was calculated in terms of the inhibition index by measuring the zone of fungal growth inhibition divided by mycelium diameter of the positive control. The test was performed by triplicate.
The phytopathogenic fungi used in this study were chosen for their economic importance as phytopathogens of ornamentals, seeds, fruits and vegetables. The following species from the microbiological collection of the Laboratory of Microbiology and Biomolecules (LaMiB®/UFSCar) were used in antagonism tests: Alternaria alternata, Ceratocystis paradoxa, Colletotrichum sp., three strains of Fusarium oxysporum (ATTCC 2163, an isolate from bean and an isolate from cotton), F. proliferatum, F. solani, F. verticillioides, Moniliophthora perniciosa, Phytophthora sojae, Rhizopus microsporus, Sclerotinia sclerotiorium and Sphaceloma sp. (CNPUV 102).
Antagonistic activity against clinical pathogens
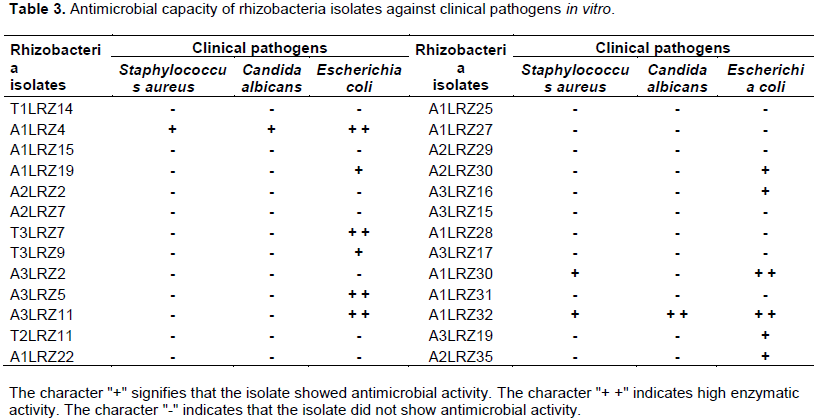
The overlay technique used in inhibition assay was adapted from Pugsley and Oudega (1987). Each isolate was inoculated in tryptic soy broth (TSB) and incubated for 72 h at 28°C. Then the inoculum was spread with sterile swabs on TSA plate and after incubation for 72 h at 28°C, an agar block was cut out in the form of a 1 cm diameter cylindrical disk. The clinical pathogens Candida albicans ATCC 10231, Escherichia coli ATCC 25923 and Staphylococcus aureus ATCC 25922 were used in this test after being reactivated and spread with swabs on the surfaces of plates containing TSA medium. After the formation of a layer of pathogenic microbial cells, the previously prepared agar blocks of twenty-six isolates selected for their performance in the direct plant-growth promotion tests (BNF, PS and IAA production) and enzymatic activity assays were added to plates, one agar block per quadrant. After incubation at 37°C for 48 h, the absence or presence of an antibiosis halo around the blocks was recorded. The tests were performed by triplicate.
Bacterial molecular identification
After all the evaluations, seven of the twenty-six best isolates were selected randomly and were identified by partial sequencing of 16S rDNA. The DNA extraction was performed using Bacteria DNA Preparation Kit (Cellco Biotec do Brasil Ltda., Sao Paulo, Brazil) according to the manufacturer's specifications. The concentration and quality of the extracted nucleic acids were analyzed on an agarose gel (1.0%) (3 volts/cm) with the GelRed nucleic acid gel stain (1.0 mg/ml) together with a 1-kb DNA Ladder RTU weight marker (Kasvi, Sao Jose dos Pinhais, Brazil). The 16S rDNA was amplified using the primers V3F (5’- CCA GAC TCC TAC GGG AGG CAG - 3’, forward sequence) and V6R (5’-ACA TTT CAC AAC ACG AGC TGA CGA - 3’, reverse sequence). Polymerase chain reaction (PCR) was performed in a 15 μl volume containing 3.75 mM MgCl2, 0.2 μM dNTPs, 0.2 μM of each primer, 2.5 U Taq DNA polymerase (Jena Bioscience, Jena, Germany) and 1 μl template DNA. The reaction conditions for the thermocycler (Eppendorf AG, Hamburg, Germany) were as follows: initial denaturation for 10 min at 95°C; 35 cycles at 95°C for 20 s, annealing for 30 s at 52°C and primer extension for 30 s at 72°C; followed by final extension for 10 min at 72°C. The reaction products were analyzed on an agarose gel (1.0%) (3 volts/cm) with the GelRed nucleic acid gel stain (1.0 mg/mL) together with a 1-kb DNA Ladder RTU weight marker (Kasvi, Sao Jose dos Pinhais, Brazil). Then the agarose gel was analyzed on Gel Doc™ EZ System/Image Lab™ software (Bio-Rad Laboratories, CA, USA).
The 16S rDNA PCR products (approximately 1400 bp in size) were purified using the QIAquick® PCR purification kit (Qiagen, Hilden, Germany) and sequenced at the Department of Biological Sciences, School of Pharmaceutical Sciences, São Paulo State University (UNESP), Araraquara campus, SP, Brazil. Sequencing was performed using the primer V6R. Bacterial identification was performed by comparison with sequences deposited in GenBank (National Center for Biotechnology Information, NCBI, www.ncbi.nlm.nih.gov) with more than 97% homology , using the BLASTn tool for this purpose.
Statistical analyses
Data obtained from each semiquantitative assay using a completely randomized design with three (IAA production and the antifungal activity index) or four (phosphorus solubilization index) replicates were subjected to one-way ANOVA. The means were compared and clustered using the Scott-Knott test (p < 0.05). Analyses were conducted using R Statistical Software (version 2.14.0; R Foundation for Statistical Computing, Vienna, Austria).
Bacterial isolation and selection, evaluation of the direct mechanisms of plant growth promotion and enzymatic activity assays
One hundred thirty-one bacterial isolates were successfully isolated from the rhizosphere of S. lycocarpum St. Hil. soil samples. The results of the 26 most promising isolates for biological nitrogen fixation, phosphate solubilization, IAA production and enzyme activity tests are shown in Table 1. Among the 131 isolates evaluated, 32.1% were able to form a typical subsuperficial whitish ‘veil-like’ pellicle in nitrogen-free semisolid medium after incubation and were considered to be positive. All the isolates were able to produce IAA, which varied in concentration from 0.07 to 134.1 μg/ml. The calcium phosphate solubilizing capacity trials showed that 45.8% of the isolates exhibited a positive reaction. The T1LRZ1, T1LRZ6, T3LRZ4, A1LRZ4, A1LRZ7 (Microbacterium sp.), A1LRZ14, A1LRZ22 (Bacillus sp.), A1LRZ32, A2LRZ8, A3LRZ2, A3LRZ5 and A3LRZ11 (Burkholderia sp.) isolates showed high solubilization potential. The T3LRZ29, A1LRZ1, A1LRZ3, A1LRZ11, A2LRZ2 (Bacillus sp.) and A2LRZ7 isolates showed medium solubilization potential. The remaining isolates had low potential or were considered negative in the test. Eighty-eight percent of the isolates were able to produce at least 1 of the 7 evaluated enzymes. Regarding pectinolytic activity, 22.3% of the evaluated isolates showed enzymatic activity of polygalacturonase and 40.2% of the evaluated isolates showed pectate lyase activity. Amylolytic activity was observed in 42.9% of the evaluated isolates, esterase activity in 47.3%, lipolytic activity in 27.7%, proteolytic activity in 60.7% and cellulolytic activity in 42.9%. Some isolates exhibited 4 or more enzymatic activities: T1LRZ14, A1LRZ19, A2LRZ7, T3LRZ7 (Bacillus sp.), T3LRZ9, T2LRZ11, A1LRZ22 (Bacillus sp.), A2LRZ30 (Bacillus sp.), A3LRZ16, A3LRZ15, A1LRZ28, A3LRZ19, A1LRZ30 and A2LRZ35 (Bacillus sp.).
Antagonistic activity against phytopathogenic fungi and clinical pathogens
Twenty-six bacterial isolates were evaluated in visual agar plate assays against phytopathogenic fungi. A total of 50% of the isolates showed antagonist activity against Sclerotinia sclerotiorium, 58% against Moniliophthora perniciosa, 38% against Fusarium solani, 40% against Fusarium oxysporum (ATTCC 2163), 52% against Sphaceloma sp., 61% against Ceratocystis paradoxa, 23% against Alternaria alternata, 46% against Fusarium proliferatum, 50% against Colletotrichum sp., 31% against Fusarium verticillioides, 33% against Fusarium oxysporum (bean), 47% against Fusarium oxysporum (cotton), 32% against Phytophthora sojae and 46% against Rhizopus microsporus. Ten isolates were selected for plate pairing assays and the mycelium growth inhibition percentages are shown in Table 2. Among the 26 bacteria evaluated, 7.7% exhibited antagonism to Candida albicans, 46% to Escherichia coli and 11.5% to Staphylococcus aureus (Table 3).
Molecular identification of bacterial isolates
The seven first sequenced isolates were chosen randomly from the twenty-six best isolates and have presented genera described in the literature as plant growth promoters (Kumar et al. 2017; Kashyap et al. 2019). The identified strains belonged to the genera Bacillus (strains A1LRZ22, A2LRZ2, A2LRZ30, A2LRZ35 and T3LRZ7), Burkholderia (A3LRZ11) and Microbacterium (A1LRZ27). The nucleotide sequences were deposited in GenBank with the accession numbers MT415822, MT415823, MT415824, MT415825, MT415826, MT415827 and MT415828.
The Brazilian tropical savannah (cerrado) has a high diversity of microorganisms. However, studies focusing on the isolation and characterization of rhizobacteria from native plant specimens from the Brazilian savannah biome are rare. To our knowledge, the present work is the first report on the characterization, identification and isolation of rhizospheric bacteria from S. lycocarpum in Brazil. The cerrado soil is acidic, had high concentrations of aluminum, low level of exchangeable cations and low availability of nutrients, mainly carbon, nitrogen and phosphorus, which come from the slow mineralization of organic matter. Physical-chemical parameters of the soil are common in agriculture, but it is necessary to understand the soil as a dynamic living system and its biological indicators, such as the resident microbiota, are directly related to the balance of this system. PGPR have a fundamental role in soil fertility, health and quality, in addition to being involved in the biogeochemical cycles of nitrogen, phosphorus, carbon and other chemical elements (Procopio and Barreto, 2021).
The flow of a nitrogen in the cerrado soil involves FBN, process responsible for approximately 65% of the total N introduced into soil. In agriculture, the need to maintain soil fertility leads to anthropogenic application of nitrogen fertilizers, which leads to a disturbance of the biogeochemical cycle. The efficient associations between diazotrophic microorganisms and several economically and environmentally important plant species can supply most of the nutritional needs related to nitrogen, thus reducing the use of nitrogen fertilizers. In this study, 32.1% of the isolates were able to grow in NFb and this result corroborates with previous studies (Grobelak et al., 2015; Habibi et al., 2019; Fukuda et al., 2021), emphasizing the BNF potential of rhizobacteria associated with plants.
Phosphorus is the second most required element by plants and is fundamental for structural development and metabolism, it is found in greater quantities in the shallower layers of the soil. Bacillus sp., Burkholderia sp. and Microbacterium sp. are described in the literature as having the ability to solubilize inorganic phosphate (Midekssa et al., 2015). In the present study, 45.8% of the evaluated isolates exhibited this ability, including isolates belonging to the aforementioned genera. Twelve isolates (9.2%) featured high solubilization potential (SI greater than 3.0). Phosphate-solubilizing bacteria (PSB) are important not only for the growth of plants but also for reducing the quantity and increasing the quality (in terms of crop yield) of manufactured fertilizers (Quecine et al., 2014). The beneficial in vivo effects of PSB have been evidenced in studies conducted with Triticum aestivum L. (wheat). The addition of an inoculum containing strains of Bacillus sp., Burkholderia sp. and Microbacterium sp. to a conventional phosphate fertilizer induced better plant development than the use of the fertilizer alone (Chauhan et al., 2013).
In the present work, rhizobacteria isolates produced indolic compounds, which plants can use as growth regulators. A previous study recorded a significant increase in the number of stem branches after inoculation with indolic compound-producing PGPR in Brassica napus (Asghar e al., 2004). The ranges of the beneficial and toxic quantities of IAA common to all plants are not yet known and the effects of these amounts may vary. Still, these isolates deserve attention since they have shown other mechanisms for promoting plant growth that can be combined with the indolic compounds production (Taiz and Zeiger, 2013).
Over 70% of diseases that affect crops of high economic importance are caused by fungi (Agrios, 2005). Fusarium spp., for example, causes diseases in soy (Glycine max), tobacco (Nicotiana tabacum) and beans (Phaseolus vulgaris), reducing both the quality and quantity of crops, resulting in losses (Matarese et al., 2012). A study reported strains of Burkholderia sp. isolated from sugarcane inhibiting growth of Fusarium verticillioides (Mendes et al., 2007). Bacillus sp. has also shown an antagonistic capability against the phytopathogen Alternaria alternate (Abdalla et al., 2014). Isolates belonging to the genera Bacillus and Burkholderia demonstrated excellent results for mycelial inhibition of phytopathogenic fungi and in antibiosis tests against clinical pathogens in this study.
Rhizospheric and endophytic bacteria isolated from medicinal plants of the cerrado represent a large reservoir of secondary metabolites; several species producing bioactive compounds can be found within this niche (Piza et al., 2016). By analyzing the results of this study, we highlight isolates (A1LRZ32 and A1LRZ4) that exhibited antibiosis against the three evaluated pathogens. Several studies have reported that bacteria of different genera, including Bacillus and Burkholderia, exhibit enzymatic activities of amylases, cellulases, esterases, lipases, pectinases and proteases (Khan et al., 2017; Liu and Kokare, 2017). Enzymes from cerrado soil microorganisms have already been characterized structurally and functionally and even successfully applied to the manufacture of products (Silva et al., 2010; Istvan et al., 2018).
Our study presented several isolates with 5 or more enzymatic activities, confirming the usefulness of rhizobacteria isolated from a cerrado plant as a source for bioprospecting enzymes of industrial interest. This finding also reinforces the need to protect this biome, which has great economic notoriety in addition to its ecological importance. Some PGPR strains are already being used commercially in the composition of bioproducts for nutrition and control of plant diseases, such as Nemix (Bacillus spp.) (AgriLife/Chr Hansen, Brazil) and Life® (PGPR consortia) (Biomax, India). Several species of Bacillus sp. have great advantages over other bacterial genera due to their rapid growth and their ability to form resistant spores against variations in pH, temperature, agrochemicals, fertilizers and storage time, allowing its use in the formulation of stable bioproducts and in biological control programs (Fantinel et al., 2018).
In conclusion, bacteria belonging to the genera Bacillus, Burkholderia and Microbacterium isolated from the rhizosphere of S. lycocarpum St. Hil. have in vitro potential for promoting plant growth, both directly and indirectly (antagonisms). The rhizobacteria associated with Solanum lycocarpum have enzymatic and antibiosis activities against pathogens, showing their importance and usefulness as sources of bioprospecting of enzymes and secondary metabolites, although further evaluation is necessary. This also shows the importance of the cerrado as a reservoir of biodiversity.
The authors have not declared any conflict of interests.
This study was financed in part by the Coordenação de Aperfeicoamento de Pessoal de Nivel Superior (CAPES), Brazil, Finance Code 001. We are grateful for FAPESP 2016/13423-5 awarded to CPS.
REFERENCES
|
Abdalla SA, Algam SAA, Ibrahim EA, Naim AME (2014). In vitro screening of Bacillus isolates for biological control of early blight disease of tomato in Shambat soil. World Journal of Agriculture Research 2(2):47-50.
Crossref
|
|
|
|
Agrios GN (2005). Plant Pathology, 5th edition. Elsevier Academic Press.
|
|
|
|
|
Aragona M, Setz EZF (2001). Diet of the maned wolf, Chrysocyon brachyurus (Mammalia: Canidae), during wet and dry seasons at Ibitipoca State Park, Brazil. Journal of Zoology 254(1):131-136.
Crossref
|
|
|
|
|
Araújo ND, Coelho VPDM, Agra MDF (2010). The pharmacobotanical comparative study of leaves of Solanum crinitum Lam., Solanum gomphodes Dunal and Solanum lycocarpum A. St-Hil (Solanaceae). Revista Brasileira de Farmacognosia 20(5):666-674.
Crossref
|
|
|
|
|
Asghar HN, Zahir ZA, Arshad M (2004). Screening rhizobacteria for improving the growth, yield and oil content of canola (Brassica napus L.). Australian Journal of Agriculture Research 55(2):187-194.
Crossref
|
|
|
|
|
Bailão EFLC, Devilla IA, Conceição EC, Borges LL (2015). Bioactive compounds found in Brazilian Cerrado fruits. Internacional Journal of Molecular Sciences 16:23760-23783.
Crossref
|
|
|
|
|
Batista BD, Lacava PT, Ferrari A, Teixeira-Silva NS, Bonatelli ML, Tsui S, Mondim M, Kitajima EW, Pereira JO, Azevedo JL, Quecine MC (2018). Screening of tropically derived, multi-trait plant growthpromoting rhizobacteria and evaluation of corn and soybean colonization ability. Microbiolical Research 206:33-42.
Crossref
|
|
|
|
|
Berraquero FR, Baya AM, Cormenzana AR (1976). Establecimiento de índices para el estudio de la solubilizacion de fosfatos por bacterias del suelo. Ars Pharmaceutica 17:399-406.
|
|
|
|
|
Bric JM, Bostock RM, Silverstone SE (1991). Rapid in situ assay for indoleacetic acid production by bacteria immobilized on a nitrocellulose membrane. Applied Environmental Microbiology 57(2):535-538.
Crossref
|
|
|
|
|
Calvo P, Nelson L, Kloepper JW (2014). Agricultural uses of plant biostimulants. Plant Soil 383:3-41.
Crossref
|
|
|
|
|
Carrim AJI, Barbosa EC, Vieira JDG (2006). Enzymatic activity of endophytic bacterial isolates of Jacaranda decurrens Cham. (Carobinha-do-campo). Brazilian Archives of Biology and Technology 49:353-359.
Crossref
|
|
|
|
|
Castro RA, Quecine CQ, Lacava PT, Batista BD, Luvizotto DM, Marcon J, Ferreira A, Melo IS, Azevedo JL (2014). Isolation and enzyme bioprospection of endophytic bacteria associated with plants of Brazilian mangrove ecosystem. Springerplus 3(1):1-9.
Crossref
|
|
|
|
|
Cattelan AJ, Nepomuceno AL, Moscardi F, Liberatti IA, Kaster M, Neumaier N, Saraiva OF (1999). Métodos qualitativos para determinação de características bioquímicas e fisiológicas associadas com bactérias promotoras do crescimento vegetal. Embrapa Soja 139:36.
|
|
|
|
|
Chang ST, Wang SY, Wu CL, Su YC, Kuo YH (1999). Antifungal compounds in the ethyl acetate soluble fraction of theextractives of Taiwania (Taiwania cryptomerioides Hayata) heartwood. Holzforschung 53:487-490.
Crossref
|
|
|
|
|
Chauhan H, Bagyaraj DJ, Sharma A (2013). Plant growth-promoting bacterial endophytes from sugarcane and their potential in promoting growth of the host under field conditions. Experimental Agriculture 49(1):43-52.
Crossref
|
|
|
|
|
Dobereiner J, Urquiaga S, Boddey RM (1995). Alternatives for nitrogen nutrition of crops in tropical agriculture. In: Ahmad N (ed) Nitrogen economy in tropical soils. Developments in plant and soil sciences 69:338-346.
Crossref
|
|
|
|
|
Fantinel VS, Muniz MFB, Poletto T, Dutra AF, Krahn JT, Favaretto RF, Sarzi JS (2018). In vitro biocontrol of Colletotrichum siamense using Trichoderma spp. and Bacillus thuringiensis var. kurstaki. Revista Ciência Agrícola 16(3):43-50.
Crossref
|
|
|
|
|
Ferreira-Filho AS, Quecine MC, Bogas AC, Rossetto PB, Lima AOS, Lacava PT, Azevedo JL, Araújo WL (2012). Endophytic Methylobacterium extorquens expresses a heterologous b-1,4-endoglucanase A (EglA) in Catharanthus roseus seedlings, a model host plant for Xylella fastidiosa. World Journal of Microbiology and Biotechnology 28:1475-148.
Crossref
|
|
|
|
|
Fukuda TTH, Pereira CF, Melo WGP, Menegatti C, Andrade PHM, Groppo M, Lacava PT, Currie CR, Pupo MT (2021). Insights into the ecological role of Pseudomonas spp. in an ant-plant symbiosis. Frontiers in Microbiology 12:621274.
Crossref
|
|
|
|
|
Glick BR (2012). Plant growth-promoting bacteria: mechanisms and applications. Hindawi Publishing Corporation.
Crossref
|
|
|
|
|
Grobelak A, Napora A, Kacprzak M (2015). Using plant growth-promoting rhizobacteria (PGPR) to improve plant growth. Ecological Engineering 84:22-28.
Crossref
|
|
|
|
|
Habibi S, Djedidi S, Ohkama-Ohtsu N, Sarhadi WA, Kojima K, Rallos RV, Ramirez M, Yamaya H, Sekimoto H, Yokoyama T (2019). Isolation and screening of indigenous plant growth-promoting rhizobacteria from different rice cultivars in Afghanistan Soils. Microbes and Environments 34(4):347-355.
Crossref
|
|
|
|
|
Hankin L, Anagnostakis SL (1975). The use of solid media for detection of enzyme production by fungi. Mycologia 67:597-607.
Crossref
|
|
|
|
|
Istvan P, Souza AA, Garay AV, Dos Santos DFK, de Oliveira GM, Santana RH, Lopes FAC, de Freitas SM, Barbosa JARG, Krüger RH (2018). Structural and functional characterization of a novel lipolytic enzyme from a Brazilian Cerrado soil metagenomic library. Biotechnology letters, 40(9):1395-1406.
Crossref
|
|
|
|
|
Kashyap BK, Solanki MK, Pandey AK, Prabha S, Kumar P, Kumari B (2019). Bacillus as plant growth promoting rhizobacteria (PGPR): A promising green agriculture technology. In: Ansari R., Mahmood I. (eds) Plant Health Under Biotic Stress. Springer.
Crossref
|
|
|
|
|
Khan AL, Shahzad R, Al-Harrasi A, Lee IJ (2017). Endophytic microbes: a resource for producing extracellular enzymes. In: Maheshwari D, Annapurna K (eds) Endophytes: crop productivity and protection. Springer 16:95-110.
Crossref
|
|
|
|
|
Kloepper JW, Schroth MN (1978). Plant growth-promoting rhizobacteria on radishes. In: Station de pathologie végétale et phyto-bactériologie (ed), Proceedings of the 4th International Conference on Plant Pathogenic Bacteria 2:879-882.
|
|
|
|
|
Kumar H, Dubey RC, Maheshwari DK (2017). Seed-coating fenugreek with Burkholderia rhizobacteria enhances yield in field trials and can combat Fusarium wilt. Rhizosphere 3:92-99.
Crossref
|
|
|
|
|
Liu X, Kokare C (2017). Microbial enzymes of use in industry. In: Brahmachari G (ed) Biotechnology of microbial enzymes. Elsevier Academic Press 267-298.
Crossref
|
|
|
|
|
Matarese F, Sarrocco S, Gruber S, Seidl-Seiboth V, Vannacci G (2012). Biocontrol of Fusarium head blight: interactions between Trichoderma and mycotoxigenic Fusarium. Microbiology 158:98-106.
Crossref
|
|
|
|
|
Mendes R, Pizzirani-Kleiner AA, Araújo WL, Raaijmakers JM (2007). Diversity of cultivated endophytic bacteria from sugarcane: genetic and biochemical characterization of Burkholderia cepacia complex isolates. Applied Environmental Microbiology 73:7259-7267.
Crossref
|
|
|
|
|
Midekssa M, Loscher C, Schmitz R, Assefa F (2015). Characterization of phosphate solubilizing rhizobacteria isolated from lentil growing areas of Ethiopia. African Journal of Microbiology Research 9:1637-1648.
Crossref
|
|
|
|
|
Miller J (1972). Experiment in molecular genetics. Cold Spring Harbor Laboratory Press 82-85.
|
|
|
|
|
Oliveira-Filho AT, Ratter JA (2002). Vegetation physiognomies and woody flora of the Cerrado biome. In: Oliveira PS, Marquis RJ (eds) The Cerrados of Brazil: ecology and natural history of a neotropical savanna. Columbia University Press pp. 91-120.
Crossref
|
|
|
|
|
Piza ACMT, Lima LCPS, Hokka CO, Sousa CP (2016). Endophytic Nocardiopsis dassonvillei and Amycolatopsis orientalis isolated from Brazilian tropical savannah presented antibiosis against pathogens. In: Méndez-Vilas A (ed) Microbes in the spotlight: recent progress in the understanding of beneficial and harmful microorganisms, 1st edition. BrownWalker Press 1:264-266.
|
|
|
|
|
Procópio L, Barreto C (2021). The soil microbiomes of the Brazilian Cerrado. Journal of Soils and Sediments pp. 1-16.
Crossref
|
|
|
|
|
Pugsley AP, Oudega B (1987). Methods for studying colicins and their plasmids. In: Hardy KG (ed) Plasmids, a practical approach. Press pp. 105-161.
|
|
|
|
|
Quecine MC, Batista BD, Lacava PT (2014). Diversity and biotechnological potential of plant-associated endophytic bacteria. In: Kumar PA, Govil JN (eds) Biotechnology: Plant Biotechnology. Studium Press LLC 2:377-423.
|
|
|
|
|
Raaijmakers JM, Mazzola M (2012). Diversity and natural functions of antibiotics produced by beneficial and plant pathogenic bacteria. Annual Review of Phytopathology 50:403-424.
Crossref
|
|
|
|
|
Ramadan EM, AbdelHafez AA, Hassan EA, Saber FM (2016). Plant growth promoting rhizobacteria and their potential for biocontrol of phytopathogens. African Journal of Microbiology Research 10(15):486-504.
Crossref
|
|
|
|
|
Schmidt EL, Belser LW (1983). Nitrifying bacteria. In: Page, AL (ed) Methods of soil analysis: part 2 chemical and microbiological properties. Madison pp. 1027-1042.
Crossref
|
|
|
|
|
Sierra GA (1957). A simple method for the detection of lypoytic activity of microorganisms and some observations on the influence of the contact between cells and fatty substracts. Antonine van Leeuwenhoeck 28:15-22.
Crossref
|
|
|
|
|
Silva RDN, Quintino FP, Monteiro VN, Asquieri ER (2010). Production of glucose and fructose syrups from cassava (Manihot esculenta Crantz) starch using enzymes produced by microorganisms isolated from Brazilian Cerrado soil. Food Science and Technology 30(1):213-217.
Crossref
|
|
|
|
|
Sousa CP, Serrano NFG, Lacava PT (2017). Endophytic Microorganisms of the Tropical Savannah: A Promising Source of Bioactive Molecules. In: de Azevedo J, Quecine M (eds) Diversity and Benefits of Microorganisms from the Tropics. Springer pp. 57-70.
Crossref
|
|
|
|
|
Taiz L, Zeiger E (2013). Fisiologia Vegetal. 5th Edition, Artmed, Porto Alegre 918 p.
|
|
|
|
|
Teather RM, Wood PJ (1982). Use of Congo red-polysaccharide interactions in enumeration and characterization of cellulolytic bacteria from bovine rumen. Applied Environmental Microbiology 43(4):777-780.
Crossref
|
|
|
|
|
Zarnowski R, Connolly PA, Wheat LJ, Woods JP (2007). Production of extracellular proteolytic activity by Histoplasma capsulatum grown in Histoplasma-macrophage medium is limited to restriction fragment length polymorphism class 1 isolates. Diagnostic Microbiology and Infectious Disease 59:3947.
Crossref
|
|