Review

ABSTRACT
Aflatoxin contamination of food and feed commodities, caused by Aspergillus section Flavi group of fungi, is a serious problem worldwide. Exposure through consumption of contaminated food and feed has deleterious effects on human and animal health. Therefore, aflatoxin contaminated products are a barrier to international trade of agricultural commodities. Not all fungi from Aspergillus section Flavi produce aflatoxins. Hence it is important to differentiate Aspergillus spp. into toxigenic and atoxigenic species to better understand their population structure in a specific environment. A range of methods are available today, including cultural, analytical and molecular methods, to identify the toxin producing ability of isolates from section Flavi. A comprehensive review of these methods would be of great use for researchers in developing nations where fully equipped aflatoxin detection laboratories are lacking. In this paper we critically reviewed the cultural and molecular methods of detecting aflatoxigenic Aspergillus species and their precision.
Key words: Toxigenic Aspergillus, atoxigenic Aspergillus, cultural methods, PCR based methods.
INTRODUCTION
Aflatoxins are secondary metabolites produced by Aspergillus section Flavi group of fungi. These aflatoxins are potent carcinogenic, teratogenic, mutagenic, hepatotoxic and immunosuppressive agents that cause significant damage to human and animal health (Moss, 2002; Saleemullah et al., 2006). The Aspergillus section Flavi comprises two groups of species; first group include A. flavus, A. parasiticus, A. nomius, A. pseudotamarii and A. bombycis. Of these, the A. flavus and A. parasiticus are the predominant aflatoxin producers (Cary and Ehrlich, 2006; Ehrlich et al., 2007). Other Aspergillus spp. reported to be aflatoxigenic in this section are A. pseudocaelatus (B1, B2 and G1, G2 aflatoxins), A. pseudonomius (B1 and not G-type aflatoxins) (Varga et al., 2011) and A. parvisclerotigenus (Godet and Munaut, 2010). The second group comprises atoxigenic species such as A. oryzae, A. sojae and A. tamarii (Kumeda and Asao, 2001). The atoxigenic species such as A. sojae and A. oryzae are generally used as starters in fermen-tation of foods (Chang et al., 2007). Particularly, A. sojae and A. oryzae are perceived as atoxigenic variants of A. flavus and A. parasiticus (Klich and Pitt, 1988). Other species of Aspergillus not included in section Flavi are also reported to be aflatoxigenic (Chang et al., 2007). For example, A. ochraceoroseus (from section Ochraceroesi); the ascomycete fungi Emericella astellata and E. venezuelensis (Aspergillus section Nidulantes) also produce B1 (Cary et al., 2005; Klich et al., 2000; Frisvad et al., 1999, 2005).
The predominant aflatoxigenic species of Aspergillus; A. flavus and A. parasiticus produce aflatoxins, that are a group of 20 closely related secondary metabolites (Liu and Wu, 2010; Snigdha et al., 2013). These fungi are ubiquitous, they are common soil inhabitants, air-borne, and are also found in crops and foods at both pre-and post-harvest stages (Waliyar et al., 1994; Jaime-Garcia and Cotty, 2004; Williams et al., 2004). Among different aflatoxins, the naturally occurring and well-known ones are aflatoxin B1 (AFB1), B2 (AFB2), G1 (AFG1), and G2 (AFG2) (Gimeno, 2004; Saleemullah et al., 2006; Strosnider et al., 2006). Of them, AFB1 is categorized as a group 1 carcinogen (Castegnaro and McGregor, 1998), and is the most prevalent aflatoxin. Predominantly, these toxins affect peanuts, corn, cotton seed, tree nuts, pea, sorghum, rice, pistachio, oilseed rape, sunflower seed, figs, spices, meats, dairy products, and fruit juices (apple, guava) (Abdin et al., 2010). Further, to this, in occupational settings, mycotoxin exposure is one of the major health concerns. Monitoring for mycotoxin exposure is therefore mandatory so as to take adequate precautionary steps. Presently, monitoring on Aflatoxin B1 is rare at occupational settings. In this context, occupational settings with potential residents of aflatoxigenic A. flavus strains needs enumeration. It is also at this juncture, the application aspects of the current review would be prudent. Knowledge on differentiation of atoxigenic and toxigenic strains would finally contribute to enhancement of precautionary safety systems in various occupational settings.
There are morphological similarities between aflatoxigenic and non-aflatoxigenic Aspergillus species. For example, A. sojae is morphologically similar to A. parasiticus and it is believed that A. sojae is a domesticated strain of A. parasiticus (Chang et al., 2007). However, few distinguishing characters such as color, texture and conidial diameter separate these two species (Klich, 2002). Precise detection of aflatoxigenic Aspergillus species is important for both research and mitigation. In this paper, we have comprehensively reviewed cultural and molecular methods of detection of aflatoxigenic Aspergillus spp. and their differentiation using cultural and molecular methods. Current analytical methods used to validate the aflatoxin production and quantification include thin layer chromatography (TLC) (Stroka and Anklam, 2000), high-performance liquid chromatography (HPLC) (Seitz, 1975; Sobolev and Dorner, 2002; Trucksess et al., 1991), liquid chromatography /mass spectroscopy (LC/MS), enzyme linked immune-sorbent assay (ELISA) (Patey et al., 1989) and immunoaffinity with fluorescence (Nasir and Jolley, 2002). Each of these methods has advantages and limitations. For example, cultural methods though inexpensive, but are less sensitive, affecting accuracy (Abbas et al., 2004a). On the other hand, molecular techniques provide rapid diagnosis because of their high sensitivity, specificity, and are currently in use for detection of aflatoxigenic strains of A. flavus and A. parasiticus (Shapira et al., 1996; Sweeney et al., 2000).
Since, the toxigenic profiles of both A. flavus and A. parasiticus are mostly different; adopting a single method has not yet been reliable in differentiation. The compiled information may be useful in devising a polyphasic, cost effective and robust approaches using one or more methods that allow accurate differentiation between aflatoxin producing and non-producing strains of Aspergillus.
CULTURAL METHODS
Cultural methods for detecting aflatoxins either rely on quantification of purified extracts (Filtenborg and Frisvad, 1980; Shotwell et al., 1966), or on qualitative assess-ments of fluorescence (Bennett and Goldblatt, 1973; de Vogel et al., 1965; Hara et al., 1974; Lin and Dianese, 1976) or UV absorption (Yabe et al., 1987). They include a) blue fluorescence (FL) (particularly in the presence of an enhancer in the medium such as β-cyclodextrin (Fente et al., 2001; Ordaz et al., 2003); b) yellow pigmentation (YP) on the undersides of colonies (Gupta and Gopal, 2002; Lin and Dianese, 1976; Odhiambo et al., 2014); and c) color change of the yellow pigment to plum-red on exposure of the culture to ammonium hydroxide vapor (AV) (Saito and Machida, 1999; Abbas et al., 2004a; Odhiambo et al., 2014).
Different media are in use for growing aflatoxigenic Aspergillus spp. They include Aspergillus flavus and parasiticus agar (AFPA) (Pitt et al., 1983), Czapek’s yeast extract agar (CYA), yeast extract sucrose agar medium (YES), coconut agar medium (CAM), aflatoxin producing ability medium (APA) (Jaimez et al., 2003b). However, the toxin production in these media varies with extraneous factors such as pH, temperature and time. Incubation for a period of five days is necessary for toxin production in YES (Gqaleni et al., 1996; Leontopoulos et al., 2003). Aflatoxin production (AFB1) by A. flavus and A. parasiticus in cheese and rice for toxigenic isolates can be peak at 7, 10, 14, 21 and 28 days. Further, maximum production of AFB1 will be at 14 days (Park and Bullerman, 1983).
Different reports on the potential of media in supporting toxin production are available. Ritter et al. (2011) showed that YES media is better compared to CYA, whereas Abranson and Clear (1996) proved that CYA agar was slightly better than YES agar. Reddy et al. (2009) reported that all toxigenic isolates of A. flavus produced AFB1 on YES media, whereas, 65%, and 53% of the same isolates produced aflatoxin on Czapek’s and PDA agar, respectively. However, none of the toxigenic isolates produced AFB1 in AFPA medium. In another study, Fente et al. (2001) showed that YES media was comparatively superior in supporting aflatoxin production over Czapek’s, aflatoxin producing ability (APA) media, and different coconut agars. In contrast, Desai and Ghosh (2003) reported high AFB1 production in Czapek’s agar over APA and CAM.
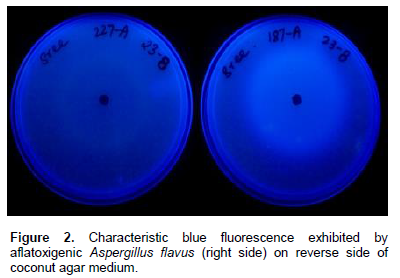
A combination of media can be of great advantage in determining the toxigenic potential of fungi. For example, combined use of CYA and YES media is advantageous since both these media allow production of different mycotoxins (Bragulat et al., 2001; Frisvad and Filtenborg, 1983). A new media containing Czapek’s yeast extract agar (CYA) and YES + 0.6% of YCSD (commercial YES medium supplemented with 0.3% Mβ-cyd and 0.6% sodium desoxycholate) is used to detect aflatoxigenic potential of A. flavus strains. When grown on this media, previously verified toxigenic strains have developed a fluorescent ring under UV light. Other media that have potential to detect aflatoxigenic A. flavus and A. niger are PDA + NaCl and APA media that produce fluorescence around the colonies. Further, the diameter of the beige ring and the intensity of its fluorescence under UV increased over time up to four days (Almoammar et al., 2013). Coconut agar medium (CAM) is also generally used for rapid detection of aflatoxin production by Aspergillus spp. (Lin and Dianese, 1976). Cultivation on CAM is generally a preliminary tool to detect aflatoxin production (de Vogel et al., 1965; Arseculeratne et al., 1969; Lin and Dianese, 1976; Davis et al., 1987). Liquid formulation of CAM is also used for detecting aflatoxins by a microplate fluorescent reader-based assay (Degola et al., 2011). A visible beige ring can be observed, under UV light, in cultures of aflatoxigenic fungi (Figure 1). Further, a blue fluorescence surrounding aflatoxigenic colonies under UV light is also seen on the reverse of the plates (Almoammar et al., 2013) (Figure 2). Isolates thought to be atoxigenic also produced toxins in CAM under optimal conditions (Hoeltz, 2005). However, due to the frequent false negative results, CAM is an unreliable method for detecting the toxigenic potential of A. flavus and A. parasiticus (Taniwaki, 1996). Other coconut culture media in use for detecting toxigenic strains include coconut milk agar (CMA), coconut extract agar, coconut cream agar (Fente et al., 2001; Davis et al., 1987; Dyer and McCammon, 1994), fresh coconut extract (FCE) and commercial coconut extract (CCE) as substrates (Lin and Dianese, 1976). Coconut cream agar is relatively more effective than other synthetic coconut media due to desiccated coconut as the ingredient (Dyer and McCammon, 1994). Another reliable substrate for rapid detection of aflatoxigenic fungi is palm kernel. Yellow pigmentation of toxigenic isolates in palm kernel media is indicative of the presence of aflatoxins. Compared to desiccated coconut agar, the fluorescence nature, intensity of diffusion of water soluble fluorescent compounds of isolates was unique in palm kernel media (Atanda et al., 2005).
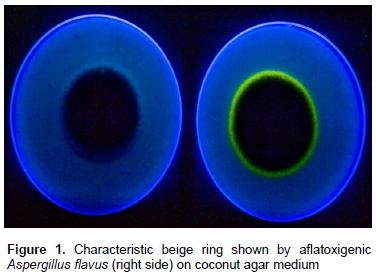
Presence of a beige ring around the colonies on media and its fluorescence under UV light is an indication of aflatoxin-producing ability of strains. Quantitative measure of agar medium fluorescence for estimating the aflatoxins are also available. Simple fluorescence is one such rapid estimation of aflatoxin levels in a solid culture medium (Cotty, 1988). In this procedure, glass test tubes containing 5 ml of agar are inoculated with spores of Aspergillus spp. and incubated for 3 days at 30°C. Fluorescence in the agar by UV light is observed. Further, the agar fluorescence is directly correlated with concentration of aflatoxins (Cotty, 1988). Hara et al. (1974) described in detail the fluorescence method of detecting aflatoxigenic strains of A. flavus by utilizing UV induced fluorescence of toxin in a modified Czapek’s solution agar containing corn steep liquor, HgCl2, and (NH4)H2PO4 instead of NaNO3. Further, the toxin presence is confirmed by thin layer chromatography (TLC) of CHCl3 extracts of the fluorescing agar. In another study, Atanda et al. (2005) reported that aflatoxigenic isolates exhibit a characteristic blue or blue green fluorescence of agar under long wave UV light against a pink background as confirmed by TLC. However, certain non-aflatoxigenic isolates also fluorescence under UV light (Almoammar et al., 2013). For example, strains of A. flavus and A. oryzae produce several substances other than aflatoxins that give a blue fluorescence under UV light. They include asperopterin A or B (Kaneko and Sanada, 1969), flavacol and deoxy-hydroxy-aspergillic acid (Yokotsuka et al., 1967).
Several other cultural media used for detection of toxigenic strains through observations on fluorescence or visible color of pigments are potato dextrose agar (PDA) and coconut agar (Davis et al., 1987; Gupta and Gopal, 2002; Lemke et al., 1988; Lin and Dianese, 1976; Saito and Machida, 1999), corn steep liquor (Hara et al., 1974; Wicklow et al., 1981) and glucose-yeast extract medium (GY-agar) (Filtenborg and Frisvad, 1980; Yabe et al., 1987; Cotty, 1988), Sabouraud dextrose and yeast extract agar when fortified with cyclodextrin (β-CD) was also a screening substrate for detecting aflatoxigenic strains (Fente et al., 2001). Complex agar medium containing sucrose, various salts, and an aqueous extract of aflatoxin-free groundnut (de Vogel et al., 1965); synthetic liquid medium (Adye and Mateles, 1964) and silica gel medium (Torrey and Marth, 1976) are also used (Fente et al., 2001). Dichloran 18% glycerol agar, DG18; dichloran rose bengal agar, DRBC; malt extract agar, MA; oxytetracycline glucose yeast extract agar, OGY; tryptic soy with lecithin and polysorbate 80, TSA; Dichloran chloramphenicol peptone agar, DCPA are also used in certain investigations on phosphorescence phenomenon to allow the detection of aflatoxigenic strains in culture media (Rojas-Duran et al., 2007).
Enrichment of culture media with cyclodextrins
Fluorescence enhancers such as cyclodextrins (CDs) are generally used to confirm the production of fluorescence by aflatoxigenic A. flavus strains. Fluorescence by AFB1 and AFG1 is greatly enhanced in presence of CDs. These CDs are cyclic oligosaccharides consisting of (α-1, 4)-linked α-D-glucopyranose units. The behaviour of AFB1 in presence of CDs was investigated by several researchers (Blais et al., 1988; Cepeda et al., 1988; Francis et al., 1988; and Vazquez et al., 1991). However, the exact mechanism behind fluorescence enhancement by aflatoxins through β-CD complexation is not clearly understood (Vazquez et al., 1992). Recently, a theoretical mechanism on fluorescence enhancement of AFB1 by β- CD was proposed (Fente et al., 2001). The β-CD form is relatively more effective in the formation of aflatoxin inclusion complexes when both α and β-CD are used (Cepeda et al., 1996). However, no significant differences on intensity of color change at different concentrations of β-CD and methylated β-CD derivatives in different media were noticed. Further, the intensity of fluorescence had increased with time (Fente et al., 2001). Conjunctive use of hydroxypropyl-β-cyclodextrin (HBC) (Cavasol®) and bile salts (cholic acid, sodium taurocholate and sodium dehydrocholate) also resulted in better detection of aflatoxigenic strains (Fente et al., 2001; Jaimez et al., 2003a, b; Rojas et al., 2005).
Pigments as indicators
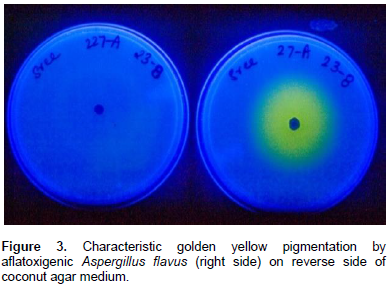
Production of yellow to orange pigments by aflatoxigenic A. flavus strains was observed for the first time by Wiseman et al. (1967). Yellow pigment formation in mycelia and media is also a basis for diagnosis of aflatoxigenic isolates (Figure 3) (Abbas et al., 2004a; Shier et al., 2005; Odhiambo et al., 2014). Aflatoxin producing A. flavus cultures isolated from insect pests on coconut produced a bright yellow pigmentation (Gupta and Gopal, 2002). The degree of yellow pigmentation is proportional to blue fluorescence in culture media (Lin and Dianese, 1976).
Vapor tests for color change
A new and rapid method for detecting toxigenic strains of A. flavus and A. parasiticus is through vapor tests. By exposing the aflatoxigenic colonies to ammonium hydroxide vapors using standard procedures (Abbas et al., 2004b), will result in quick color change of the reverse side from brownish/yellowish color to plum-red (Saito and Machida, 1999) (Figure 4). Biochemical basis of vapor tests was investigated by extracting pigments from lyophilized cultures of toxigenic strains grown on PDA. Further mixing of these pigments with ammonium hydroxide or other bases (sodium hydroxide, potassium hydroxide, sodium carbonate, and sodium bicarbonate) resulted in color change to plum-red (Abbas et al., 2004b). The pigments associated with color change are norsolorinic acid, averantin, averufin, versicolorin C, versicolorin A, versicolorin A hemiacetal, and nidurufin (Aucamp and Holzapfel, 1970; Cole, 1981; Heathcote and Dutton, 1969). All these pigments are anthraquinone intermediates in aflatoxin biosynthetic pathway (Bhatnagar et al., 2003). Of these, averufin was produced by a non-aflatoxigenic mutant of A. parasiticus (Donkersloot et al., 1972).

MOLECULAR METHODS
Application of molecular techniques in aflatoxin research is multitude. For example, molecular approaches have been used for differentiating the Aspergillus section Flavi complex, determining the phylogenetic analysis, characterization of isolates, identification of aflatoxigenic isolates from different food and feeds, diversity studies and in understanding the non-aflatoxigenicity of certain Aspergillus spp. (Montiel et al., 2003; Almoammar et al., 2013; Chang et al., 2007; Baird et al., 2006; Hatti et al., 2010; Yin et al., 2009). Applications of such studies can be useful in bringing out effective atoxigenic strains rapidly for their further use as potential biocontrol agents (Yin et al., 2009). Differentiation of aflatoxigenic and non-aflatoxigenic strains of Aspergillus section Flavi group through molecular approaches is important because conventional approaches are not entirely reliable (Criseo et al., 2001). Moreover, the molecular mechanisms responsible for loss of aflatoxigenicity of Aspergillus are not clearly understood (Schmidt-Heydt et al., 2008). Earlier reports indicated that atoxigenic A. flavus isolates were found to be majorly associated with the deletions of a part or the entire aflatoxin gene cluster (Chang et al., 2005). Defects in the aflatoxin gene, pksA in A. flavus AF36 isolate of cotton seed is also responsible for its atoxigenicity (Ehrlich and Cotty, 2004). Other reasons for atoxigenicity of Aspergillus can be attributed to large deletions in the aflatoxin gene cluster (Prieto et al., 1996). Jiang et al. (2009) reported a large fragment deletion in the aflatoxin gene cluster and further replacement of its location by a heterologous insert. PCR assays have revealed that Aspergillus isolates with entire aflatoxin gene cluster could not produce aflatoxins (Yin et al., 2009). Hence, analysis of deletion within aflatoxin gene cluster can be an effective method for rapid identification of true non-aflatoxigenic Aspergillus strains (Yin et al., 2009).
Based on molecular characterization studies, it is concluded that both toxigenic and atoxigenic A. flavus isolates are genetically similar, but some atoxigenic isolates having deletions within the aflatoxin gene cluster can be identified readily by PCR assays (Yin et al., 2009). Complete differentiation of atoxigenic and toxigenic strains is however not feasible with molecular methods such as random amplified polymorphic DNA (RAPD), Microsatellite-primed PCR (MP-PCR) profiles and DNA sequences of internal transcribed spacer (ITS) regions, pksA and omtA genes (Yin et al., 2009). Previous research has also indicated that molecular techniques such as RAPD and AFLP (amplified fragment length polymorphism) could not differentiate toxigenic and atoxigenic strains (Tran-Dinh et al., 1999; Montiel et al., 2003; Barros et al., 2007). For example, the atoxigenic A. sojae and the toxigenic A. parasiticus could not be differentiated through sequencing of the ribosomal DNA ITS (internal transcribed spacers) (Montiel et al., 2003). However, this technique could separate the A. flavus/A. parasiticus subgroup from the A. oryzae/A. flavus subgroup. In another study, Baird et al. (2006) reported that DNA amplification fingerprinting (DAF) was not effective in distinguishing aflatoxigenic and non-aflatoxigenic isolates of A. flavus. However, use of mini-hairpin primers using the arbitrary signatures from amplification profiles (ASAP) technique successfully distinguished the majority of aflatoxigenic from non-aflatoxigenic isolates (Baird et al., 2006). AFLP fingerprints however can be used for genetic differen-tiation of Aspergillus section Flavi complex fungi. In a study by Montiel et al. (2003), analysis of AFLP data consistently and clearly separated A. sojae/A. parasiticus isolates from the A. oryzae/A. flavus isolates. Further studies by Montiel et al. (2003) have identified poly-morphic sequences that could be developed into markers for aflatoxigenic A. parasiticus isolates.
Use of multiplex PCR with three sets of primers specific for three structural genes of the aflatoxin pathway (nor-1, ver-1 and omt-A) could differentiate only the aflatoxin producing fungi, A. flavus and A. parasiticus from others, but not aflatoxin producing and non-producing strains of the same species (Geisen, 1996). In another study, Rashid et al. (2008) categorized A. flavus and A. parasiticus isolates of stored wheat into aflatoxi-genics and non-aflatoxigenics using multiplex PCR based assay. Their studies on detection of four genes, AflR (regulatory gene) and structural genes such as Nor 1 (norsolorinic acid), Ver 1 (Versicolorin) and Omt (O methylsterigmatocystin) indicated that only one of the tested strains of A. parasiticus (AP4) out of 36 strains has shown the presence of these four genes. Further, it was also observed that strains with presence of aflR gene also did not produce aflatoxins (Rashid et al., 2008). Criseo et al. (2001) worked with quadruplex PCR using primers for aflR, nor-1, ver-1 and omt-A genes of the aflatoxin biosynthetic pathway for differentiating aflatoxigenic and non-aflatoxigenic strains. Their results indicated that aflatoxigenic strains gave a quadruplet pattern, indicating the presence of all the genes involved in the aflatoxin biosynthetic pathway. However, for non-aflatoxigenic strains, the results are variable with one, two, three or four banding patterns. Further, a banding pattern in few non-aflatoxigenic strains resulted in non-differentiation between these two strains.
Aflatoxin production by A. flavus is more stable in nature than in culture (Bayman and Cotty, 1993). Gene expression between Aspergillus spp. can be different despite their close relatedness at the DNA level according to microarray analysis (Wilkinson et al., 2007). Hence, understanding the aflatoxigenicity and non-aflatoxigenicity through molecular means at gene level is an important step for further rapid and precise detection of these species. Developing rapid, precise, cost-effective and less cumbersome methods in this direction will further aid in devising strategies that prevent aflatoxin contamination in agriculturally important crops.
CONCLUSIONS
For precise differentiation of toxigenic and atoxigenic strains of A. flavus, a polyphasic approach involving the use of different cultural and molecular methods is apt. False positives with respect to aflatoxin production are also not uncommon in aflatoxigenic strain detection. In this regard, analytical methods such as HPLC, TLC and ELISA that detect toxin production in the substrate can also be confirmative when used in polyphasic approach along with cultural and molecular methods. An elaborative and updated review in this area would be useful for aflatoxin researchers especially in detecting and differentiating atoxigenic and toxigenic Aspergillus spp. in crop soils that are posing severe health hazards and trade losses in several major crops.
CONFLICT OF INTERESTS
The authors did not declare any conflict of interest.
ACKNOWLEDGMENTS
The authors are thankful to the support provided under the CGIAR Research Program on Agriculture for Nutrition and Health (CRP-A4NH). This work has been undertaken as part of the research project sponsored under CRP-A4NH. ICRISAT is a member of CGIAR Consortium.
REFERENCES
|
Abbas HK, Shier WT, Horn BW, Weaver MA (2004a). Cultural methods for aflatoxin detection. Toxin Rev. 23:295-315. |
||
|
Abbas HK, Zablotowicz RM, Weaver MA, Horn, BW, Xie W, Shier WT (2004b). Comparison of cultural and analytical methods for determination of aflatoxin production by Mississippi Delta Aspergillus isolates. Can. J. Microbiol. 50(3):193-199. |
||
|
Abdin MZ, Ahmad MM, Javed S (2010). Advances in molecular detection of Aspergillus: an update. Arch. Microbiol. 192(6): 409-425. |
||
|
Abranson D, Clear RM (1996). A convenient method for assessing mycotoxin production in cultures of Aspergilli and Penicillia. J. Food Prot. 59:142-144. |
||
|
Adye J, Mateles RI (1964). Incorporation of labelled compounds into aflatoxins. Biochim. Biophys. Acta 86:418-420. |
||
|
Almoammar H, Bahkali AH, Abd-Elsalam KA (2013). A polyphasic method for the identification of aflatoxigenic Aspergillus species isolated from Camel feeds. Aust. J. Crop Sci. 7(11): 1707-1713. |
||
|
Arseculeratne SN, De Silva LM, Wijesundera S, Bandunatha CHSR (1969). Coconut as a medium for the experimental production of aflatoxin. Appl. Microbiol. 18(1):88-94. |
||
|
Atanda I, Rati ER, Ozoja M (2005). Palm Kernel: A potential substrate for rapid detection of aflatoxigenic fungi. Food Sci. Technol. Int. 11(1):67-74. |
||
|
Aucamp PJ, Holzapfel CW (1970). Polyhydroxy anthraquinones from Aspergillus versicolor, Aspergillus nidulans and Bipolaris sp. Their significance in relation to biogenetic theories on aflatoxin B1. J. S. Afr. Chem. Inst. 23: 40-56. |
||
|
Baird RE, Trigiano RN, Windham G, Williams P, Kelley R, Abbas HK, Moulton JK, Scruggs ML (2006). Comparison of aflatoxigenic and non aflatoxigenic isolates of Aspergillus flavus using DNA amplification fingerprinting techniques. Mycopathologia 161: 93-99. |
||
|
Barros GG, Chiotta ML, Reynoso MM, Torres AM, Chulze SN (2007). Molecular characterization of Aspergillus section Flavi isolates and Aspergillus flavus S and L morphotype isolates. Int. J. Food Microbiol. 108:172-177. |
||
|
Bayman P, Cotty P (1993). Genetic diversity in Aspergillus flavus: association with aflatoxin production and morphology. Can. J. Bot. 71:23-31. |
||
|
Bennett JW, Goldblatt LA (1973). The isolation of mutants of Aspergillus flavus and A. parasiticus with altered aflatoxin producing ability. Sabouraudia 11: 235-241. |
||
|
Bhatnagar D, Ehrlich KC, Cleveland TE (2003). Molecular genetic analysis and regulation of aflatoxin biosynthesis. Appl. Microbiol. Biotechnol. 61: 233-93. |
||
|
Blais J, Prognon P, Mahuzier G, Vigny P (1988). Relative affinity of 5-methoxypsoralen and 8-methoxypsoralen towards β-cyclodextrin: a fluorescence, circular dichroism and chromatographic study. J. Photochem. Photobiol. B 2(4): 455-465. |
||
|
Bragulat MR, Abarca ML, Cabanes FJ (2001). An easy screening method for fungi producing ochratoxin A in pure culture. Int. J. Food Microbiol. 71:139-144. |
||
|
Cary JW, Ehrlich KC (2006). Aflatoxigenicity in Aspergillus: molecular genetics, phylogenetic relationships and evolutionary implications. Mycopathologia 162(3):167-177. |
||
|
Cary JW, Klich MA, Beltz SB (2005). Characterization of aflatoxin-producing fungi outside of Aspergillus section Flavi. Mycologia 97: 425-432. |
||
|
Castegnaro M, McGregor D (1998). Carcinogenic risk assessment of mycotoxins. Rev. Med. Vet-Toulouse 149: 671-678. |
||
|
Cepeda A, Franco CM, Fente CA, Vazquez BI, Rodriguez JL, Prognon P, Mahuzier G (1996). Post column excitation of aflatoxins using cyclodextrins in liquid chromatography for food analysis. J. Chromatogr. A. 721(1): 69-74. |
||
|
Cepeda A, Prognon P, Mahuzier G, Blais J (1988). Improvement in the fluorometric detection of 5-methoxypsoralen by using β-cyclodextrin column. Anal. Chim. Acta 211: 333-337. |
||
|
Chang PK, Horn BW, Dorner JW (2005). Sequence breakpoints in the aflatoxin biosynthesis gene cluster and flanking regions in non aflatoxigenic Aspergillus flavus isolates. Fungal Genet. Biol. 42: 914-923. |
||
|
Chang PK, Matsushima K, Takahashi T, Yu Jiujiang, Abe K, Bhatnagar D, Yuan GF, Koyama Y, Cleveland TE (2007). Understanding non aflatoxigenicity of Aspergillus sojae: a windfall of aflatoxin biosynthesis research. Appl. Microbiol. Biotechnol. 76: 977-984. |
||
|
Cole RJ (1981). Versicolorin group. In: Handbook of toxic fungal metabolites. New York: Academic Press, pp. 94-127. |
||
|
Cotty PJ (1988). Simple fluorescence method for rapid estimation of aflatoxin levels in a solid culture medium. Appl. Environ. Microbiol. 54: 274-276. |
||
|
Criseo G, Bagnara A, Bisignano G (2001). Differentiation of aflatoxin-producing and non-producing strains of Aspergillus flavus group. Lett. Appl. Microbiol. 33: 291-295. |
||
|
Davis ND, Iyer, SK, Diener UL (1987). Improved method of screening for aflatoxin with a coconut agar medium. Appl. Environ. Microbiol. 53: 1593-1595. |
||
|
De Vogel P, Van Rhee R, Koelensmid W (1965). A rapid screening test for aflatoxin-synthesizing Aspergilli of the flavus-oryzae group. J. Appl. Bacteriol. 28: 213-220. |
||
|
Degola F, Berni E, Restivo, FM (2011). Laboratory tests for assessing efficacy of atoxigenic Aspergillus flavus strains as biocontrol agents. Int. J. Food Microbiol. 146(3): 235-243. |
||
|
Desai MR, Ghosh S (2003). Occupational exposure to airborne fungi among rice mill workers with special reference to aflatoxin producing A. flavus strains. Ann. Agric. Environ. Med. 10(2):159-162. |
||
|
Donkersloot JA, Mateles RI, Yang SS (1972). Isolation of averufin from a mutant of Aspergillus parasiticus impaired in aflatoxin biosynthesis. Biochem. Biophys. Res. Commun. 47(5): 1051-1055. |
||
|
Dyer SK, McCammon S (1994). Detection of toxigenic isolates of Aspergillus flavus and related species on coconut cream agar. J. Appl. Bacteriol. 76: 75-78. |
||
|
Ehrlich KC, Cotty PJ (2004). An isolate Aspergillus flavus used to reduce aflatoxin contamination in cottonseed has a defective polyketide synthase gene. J. Microbiol. Biotechnol. 65: 473-478. |
||
|
Ehrlich KC, Kobbeman K, Montalbano BG, Cotty PJ (2007). Aflatoxin-producing Aspergillus species from Thailand. Int. J. Food Microbiol. 114: 153-159. |
||
|
Fente CA, Ordaz JJ, Vazquez BI, Franco CM, Cepeda A (2001). New additive for cultural media for rapid identification of aflatoxin producing Aspergillus strains. Appl. Environ. Microbiol. 67: 4858-4862. |
||
|
Filtenborg O, Frisvad JC (1980). A simple screening method for toxigenic fungi in pure cultures. Lebensm. Wiss-Technol. 13:128-130. |
||
|
Francis OJ, Kirschenheuter GP, Ware GM, Barman AS, Kuan SS (1988). β-Cyclodextrin post-column fluorescence enhancement of aflatoxins for reverse-phase liquid chromatographic determination in corn. J. AOAC Int. 71: 725-728. |
||
|
Frisvad JC, Filtenborg O (1983). Classification of terverticillate penicillia based on profiles of mycotoxins and other secondary metabolites. Appl. Environ. Microbiol. 46(6): 1301-1310. |
||
|
Frisvad JC, Houbraken J, Samson RA (1999). Aspergillus species and aflatoxin production: a reappraisal, In: A.C.J. Tuijtelaars, R.A. Samson, F.M. Rombouts, S. Notermans (Eds.), Food Microbiology and Food Safety into the Next Millennium, Foundation Food Micro '99, Zeist, pp.125-126. |
||
|
Frisvad JC, Skouboe P, Samson RA (2005). Taxonomic comparison of three different groups of aflatoxin producers and a new efficient producer of aflatoxin B1, sterigmatocystin and 3-O-methylsterigmatocystin, Aspergillus rambellii sp. nov. Syst. Appl. Microbiol. 28(5): 442-453. |
||
|
Geisen R (1996). Multiplex polymerase chain reaction for the detection of potential aflatoxin and sterigmatocystin producing fungi. Syst. Appl. Microbiol. 19(3): 388-392. |
||
|
Gimeno A (2004). Aflatoxina M1 no leite. Riscos para a saúde pública, prevenção e controlo. Alimentação Animal (Revista de la Associação Portuguesa dos Industriais de Alimentos Compostos para Animais (IACA)) 49: 32-44. |
||
|
Godet M, Munaut F (2010). Molecular strategy for identification in Aspergillus section Flavi. FEMS Microbiol. Lett. 304: 157-168. |
||
|
Gqaleni N, Smith JE, Lacey J (1996). Co-production of aflatoxins and cyclopiazonic acid in isolates of Aspergillus flavus. Food Addit. Contam. 13(6): 677-685. |
||
|
Gupta A, Gopal M (2002). Aflatoxin production by Aspergillus flavus isolates pathogenic to coconut insect pests. World J. Microbiol. Biotechnol. 18(4): 329-335. |
||
|
Hara S, Fenell DL, Hesseltine CW (1974). Aflatoxin producing strains of Aspergillus flavus detected by fluorescence of agar medium under ultraviolet light. Appl. Microbiol. 27: 1118-1123. |
||
|
Hatti AD, Taware SD, Taware AS, Pangrikar PP, Chavan AM, Mukadam DS (2010). Genetic diversity of toxigenic and non-toxigenic Aspergillus flavus strains using ISSR markers. Int. J. Curr. Res. 5: 61-66. |
||
|
Heathcote JG, Dutton MF (1969). New metabolites of Aspergillus flavus. Tetrahedron 25(7): 1497-1500. |
||
|
Hoeltz M (2005). Estudo da Influencia de Manejos Pos-Clolheita na Incidencia de Fungos e Micotoxinas no Arroz (Oryza sativa L.). Dissertacao (Mestrado em Microbiologia Agricola e do Ambiente)-Universidade Federal do Rio Grande do Sul. Porto Alegre. |
||
|
Jaime-Garcia R, Cotty PJ (2004). Aspergillus flavus in soils and corn cobs in South Texas: Implications for management of aflatoxins in corn-cotton rotations. Plant Dis. 88: 1366-1371. |
||
|
Jaimez J, Fente CA, Franco CM, Cepeda A, Vazquez B (2003a). Application of a modified culture medium for the simultaneous counting of molds and yeasts and detection of aflatoxigenic strains of Aspergillus flavus and Aspergillus parasiticus. J. Food Prot. 66: 311-318. |
||
|
Jaimez J, Fente CA, Vazquez BI, Franco CM, Cepeda A (2003b). Development of a method for direct visual determination of aflatoxin production by colonies of the Aspergillus flavus group. Int. J. Food Microbiol. 83:219-225. |
||
|
Jiang H, Yan L, Ma Z (2009). Molecular characterization of an atoxigenic Aspergillus flavus strain AF051. Appl. Microbiol. Biotechnol. 83(3): 501-505. |
||
|
Kaneko Y, Sanada M (1969). Studies on the fluorescent substances produced by Aspergillus fungi. VII. Purification and isolation of asperopterin B and chemical properties of asperopterin B and A. J. Ferment. Technol. 47:8-19. |
||
|
Klich MA (2002). Identification of common Aspergillus species. Centralbureau voor Schimmelcultures, The Netherlands. |
||
|
Klich MA, Mullaney EJ, Daly CB, Cary JW (2000). Molecular and physiological aspects of aflatoxin and sterigmatocystin biosynthesis by Aspergillus tamarii and Aspergillus ochraceoroseus. Appl. Microbiol. Biot. 53(3): 605-609. |
||
|
Klich MA, Pitt JI (1988). A laboratory guide to common Aspergillus species and their teleomorphs. CSIRO Division of Food Processing, North Ryde, NSW. |
||
|
Kumeda Y, Asao T (2001). Heteroduplex panel analysis, a novel method for genetic identification of Aspergillus section Flavi strains. Appl. Environ. Microbiol. 67(9): 4084-4090. |
||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


