Review

ABSTRACT
Biofilms formation stands out in context of persistent intestinal infections caused by Enterobacteriaceae, which are associated with a high resistance to antimicrobial agents’ and phagocytosis by host defense cells. Hence, understanding the mechanisms involved in this process becomes major for the development of new preventive and therapeutic strategies. Lactic acid bacteria, including species of the genus Lactobacillus, have been associated with the prevention or dispersion of biofilms formed by pathogenic microorganisms. This effect is often associated with the antimicrobial substances production, among them organic acids, bacteriocins, hydrogen peroxide and biorsurfactants. However, the antibiofilm action of Lactobacillus seems to be strain-specific and may not be demonstrated by strains of the same genus. Thus, diet supplementation with beneficial microorganisms represents a possible strategy for prevention and treatment of intestinal infectious diseases, such as persistent or acute diarrhea caused by enteropathogenic bacteria. However, in vitro and in vivo further studies are needed to clarify the efficacy of different probiotic candidates, including commercially available products.
Keyword: Enterobacteria, biofilm, lactobacillus, antimicrobials.
INTRODUCTION
PROBIOTICS MICROORGANISMS
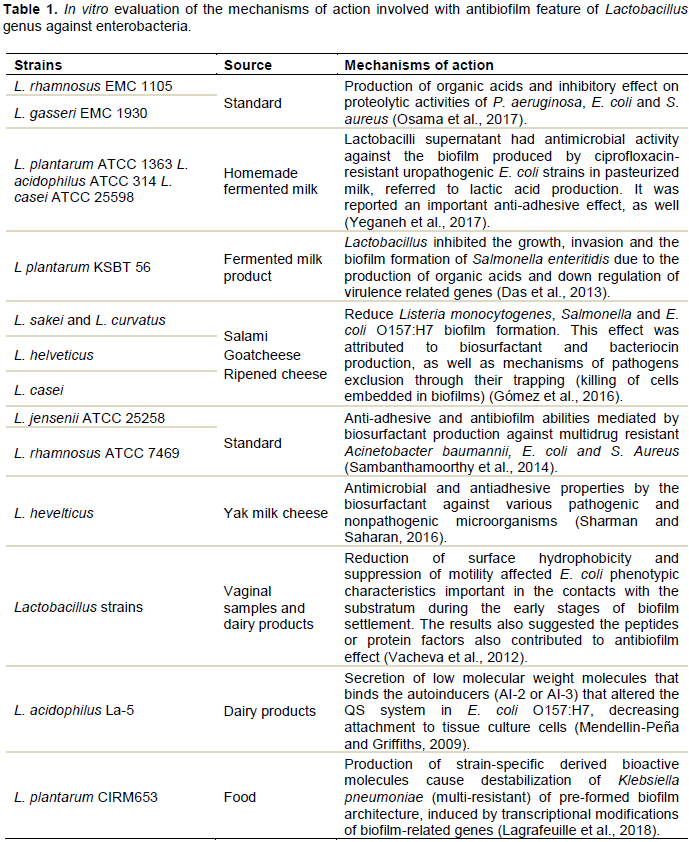
CONCLUSION
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENT
The authors show gratitude to the institutions University Center INTA and Federal University of Ceara for support during the course of this research.
REFERENCES
|
Abdelhamid AG, Esaam A, Hazaa MM (2018). Cell free preparations of probiotics exerted antibacterial and antibiofilm activities against multidrug resistant E. Coli. Saudi Pharmaceutical Journal 26(5):603-607. |
|
|
AL-Mamun M, Chowdhury T, Biswas B, Absar N (2018). Food poisoning and intoxication: A global leading concern for human health. Food Safety and Preservation pp. 307-352. |
|
|
Arioli S, Koirala R, Taverniti V, Fiore W, Guglielmetti S (2018). Quantitative recovery of viable Lactobacillus paracasei CNCM I-1572 (L. casei DG®) after gastrointestinal passage in healthy adults. Frontiers in Microbiology 9:1720. |
|
|
Balgir PP, Kaur B, Kaur T, Daroch N, Kaur G (2013). In vitro and in vivo survival and colonic adhesion of Pediococcus acidilactici MTCC5101 in human gut. BioMed research international 2013:1-9. |
|
|
Bamidele O, Jiang ZD, Dupont H (2019). Occurrence of putative virulence-related genes, aatA, aggR and aaiC, of Enteroaggregative Escherichia coli (EAEC) among adults with travelers' diarrhea acquired in Guatemala and Mexico. Microbial pathogenesis 128:97-99. |
|
|
Beltrán AR, Carraro-Lacroix LR, Bezerra CN, Cornejo M, Norambuena K,Toledo F, Araos J, Pardo F, Leiva A, Sanhueza C, Malnic G, Sobrevia L, Ramírez MA (2015). Escherichia coli heat-stable enterotoxin mediates Na+/H+ exchanger 4 inhibition involving cAMP in T84 human intestinal epithelial cells. PLoS One. 10(12):e0146042. |
|
|
Bjarnsholt T, Ciofu O, Molin S, Givskov M, Hoiby N (2013). Applying insights from biofilm biology to drug development-can a new approach be developed?. Nature Reviews Drug Discovery 12(10):791-808. |
|
|
Bustos SP, Chamorro JFV (2018). Probiotics in Acute, Antibiotic-associated and Nosocomial Diarrhea: Evidence in Pediatrics. Revista Colombiana de Gastroenterologia 33(1):41-48. |
|
|
Carey CM, Kostrzynska M, Ojha S, Thompson S (2008). The effect of probiotics and organic acids on Shiga-toxin2 gene expression in enterohemorrhagic Escherichia coli O157:H7. Journal of Microbiological Methods 73(2):125-132. |
|
|
Challinor VL, Bode HB (2015). Bioactive natural products from novel microbial sources. Annals of the New York Academy of Sciences 1354(1):82-97. |
|
|
Chau ML, Hartantyo SH, Yap M, Kang JS, Aung KT, Gutiérrez RA, Ng LC, Tam CC, Barkham T (2016). Diarrheagenic pathogens in adults attending a hospital in Singapore. BMC Infectious Diseases 16: 32. |
|
|
Chau TTH, Chau NNM, Le NTH, The HC, Vinh PV, To NTN, Ngoc NM, Tuan HM, Ngoc TLC, Kolader ME, Farrar JJ, Wolbers M, Thwaites GE, Baker S (2018). A double blind, randomized, placebo-controlled trial of Lactobacillus acidophilus for the treatment of acute watery diarrhea in vietnamese children. The Pediatric Infectious Disease Journal 37(1):35-42. |
|
|
Chen C, Wang LP, Yu JX, Chen X, Wang RN, Yang XZ, Zheng SF, Yu F, Zhang ZK, Liu SJ, Li ZJ, Chen Y (2019). Prevalence of enteropathogens in outpatients with acute diarrhea from urban and rural areas, southeast China, 2010-2014. American Journal of Tropical Medicine and Hygiene tpmd19-0171. |
|
|
Chew SY, Cheah YK, Seow HF, Sandai D, Than LTL (2015). Probiotic Lactobacillus rhamnosus GR-1 and Lactobacillus reuteri RC-14 exhibit strong antifungal effects against vulvovaginal candidiasis-causing Candida glabrata isolates. Journal of Applied Microbiology 118(5):1180-1190. |
|
|
Choi SY, Rashed SM, Hasan NA, Alam M, Islam T, Sadique A, Johura F-T, Eppinger M, Ravel J, Huq A, Cravioto A, Colwell RR (2016). Phylogenetic diversity of Vibrio cholerae associated with endemic cholera in Mexico from 1991 to 2008. mBio 7(2):e02160-15. |
|
|
Costa GN, Suguimoto HH, Miglioranzaand LHS, Gómez RJHC (2012). Antimicrobial activity of Lactobacillus and Bifidobacterium strains against pathogenic microorganisms "in vitro". Semina: Ciencias Agrárias 33(5):1839-1846. |
|
|
Cui X, Shi Y, Gu S, Yan X, Chen H, Ge J (2017). Antibacterial and Antibiofilm Activity of Lactic Acid Bacteria Isolated from Traditional Artisanal Milk Cheese from Northeast China Against Enteropathogenic Bacteria. Probiotics and Antimicrobial Proteins 1-10. |
|
|
Dallman T, Smith GP, O'Brien B, Chattaway MA, Finlay D, Gran, KA, Jenkins C (2012). Characterization of a verocytotoxin-producing enteroaggregative Escherichia coli serogroup O111:H21 strain associated with a household outbreak in Northern Ireland. Journal of Clinical Microbiology 50(12):4116-4119. |
|
|
Das JK, Mishra D, Ray P, Tripathy P, Beuria TK, Singh N, Suar M (2013) In vitro evaluation of anti-infective activity of a Lactobacillus plantarum strain against Salmonella enterica serovar Enteritidis. Gut Pathogens 5(1):11. |
|
|
Davoodabadi A, Dallal MMS, Lashani E, Ebrahimi MT (2015). Antimicrobial activity of Lactobacillus spp. isolated from fecal flora of healthy breast-fed infants against diarrheagenic Escherichia coli. Jundishapur Journal of Microbiology 8(12). |
|
|
Do Carmo MS, Santos CI, Araújo MC, Girón JA, Fernandes ES, Monteiro-Neto V (2018). Probiotics, mechanisms of action, and clinical perspectives for diarrhea management in children. Food & Function 9(10):5074-5095. |
|
|
Feltes MMC, Arisseto-Bragotto AP, Block JM (2017). Food quality, food-borne diseases, and food safety in the Brazilian food industry. Food Quality and Safety 1(1):13-27. |
|
|
Fernandes MSM (2019). Atividade antimicrobiana e antibiofilme do sobrenadante de cepas de Lactobacillus cell-free sobre isolados de Escherichia coli farmacorresistentes. MS Thesis. Federal University of Ceará, CE, Brazil. 2019. |
|
|
Fijan S (2014). Microorganisms with Claimed Probiotic Properties: An Overview of Recent Literature. Journal of Environmental Research and Public Health 11(5):4745-4767. |
|
|
Floch MH, Walker WA, Sanders ME, Nieuwdorp M, Kim AS, Brenner DA, Qamar AA, Miloh TA, Guarino A, Guslandi M, Dieleman LA, Ringel Y, Quigley EM, Brandt LJ (2015). Recommendations for probiotic use - 2015 Update: Proceedings and consensus opinion. Journal of Clinical Gastroenterology 49(Suppl 1):S69-S73. |
|
|
Gabliardi A, Totino V, Cacciotti F, Iebba V, Neroni B, Bonfiglio G, Trancassini M, Passariello C, Pantanella F, Schippa S (2018). Rebuilding gut microbiota ecosystem. Journal of Environmental Research and Public Health 15(8):1679. |
|
|
Gálvez A, Abrioul H, Benomar N, Lucas R (2010). Microbial antagonists to food-borne pathogens and biocontrol. Current Opinion in Biotechnology 21(2):142-148. |
|
|
Gómez NC, Ramiro JM, Quecan BXV, Franco BGDM (2016). Use of potential probiotic lactic acid bacteria (LAB) biofilms for the control of Listeria monocytogenes, Salmonella typhimurium and Escherichia coli O157:H7 biofilms formation. Frontiers in Microbiology 7:863. |
|
|
Gutiérrez S, Martínez-blanco H, Rodríguez-Aparicio LB, Ferrero MA (2016). Effect of fermented broth from lactic acid bacteria oh pathogenic bacteria proliferation. Journal of Dairy Science 99(4): 2654-2665. |
|
|
Guttenplan SB, Kearns DB (2013). Regulation of flagellar motility during biofilm formation. FEMS Microbiology Reviews 37(6):849-871. |
|
|
Halder D, Mandal S (2016). Antibacterial potentiality of commercially available probiotic Lactobacilli and curd Lactobacilli strains, alone and in combination, against human pathogenic bacteria. Translational Biomedicine 7:1-7. |
|
|
Haney EF, Trimble MJ, Cheng JT, Vallé Q, Hancock REW (2018). Critical assessment of methods to quantify biofilm growth and evaluate antibiofilm activity of host defence peptides. Biomolecules 8(2) 29. |
|
|
Hegar B, Waspada IM, Gunardi H, Vandenplas Y (2015). A double blind randomized trial showing probiotics to be ineffective in acute diarrhea in Indonesian children. Indian Journal Pediatrics 82(5):410-414. |
|
|
Hill C, Guarner F, Reid G, Gibson GR, Merenstein DJ, Pot B, Morelli L, Canani RB, Flint HJ, Salminem S, Calder PC, Sanders AE (2014). The international scientific association for probiotics consensus statement on the scope and appropriate use of the term probiotic. Nature reviews Gastroenterology & Hepatology 11(8):506-514. |
|
|
Hodges K, Gill R (2010). Infectious diarrhea: cellular and molecular mechanisms. Gut Microbes 1(1):4-21. |
|
|
Hu S, Wang L, Jiang Z (2017). Dietary additive probiotics modulation of intestinal microbiota. Protein and Peptides Letters 24(5):382-387. |
|
|
Hughes G, Webber MA (2017). Novel approaches to the treatment of bacterial biofilm infections. British Journal of Pharmacology 174:2237-2246. |
|
|
Iacob S, Iacob DG and Luminos LM (2019). Intestinal Microbiota as a Host Defense Mechanism to Infectious Threats. Frontiers in Microbiology 9:3328. |
|
|
Islam SU (2016). Clinical uses of probiotics. Medicine 95(5):e2658. |
|
|
Jiang Y, Ye L, Cui Y, Yang G, Yang W, Wang J, Hu J, Gu W, Shi C, Huang H, Wang C (2017). Effects of Lactobacillus rhamnosus GG on the maturation and differentiation of dendritic cells in rotavirus-infected mice. Beneficial Microbes 8(4):645-656. |
|
|
Kaur S, Sharma P, Kalia N, Singh J, Kaur S (2018). Anti-biofilm properties of the fecal probiotic Lactobacilli against Vibrio spp. Frontiers in Cellular and Infection Microbiology 8:120. |
|
|
Lääveri T, Pakkanen SH, Antikainen J, Riutta J, Mero S, Kirveskari J, Kantele A (2014). High number of diarrhoeal coinfections in travellers to Benin, West Africa. BMC Infectious Diseases 14:81. |
|
|
Lagrafeuille R, Miquel S, Balastrino D, Vareille-Delarbre M, Chain F, Langella P, Forestier C (2018). Opposing effect of Lactobacillus on in vitro Klebsiella pneumoniae in biofilm and in an in vivo intestinal colonization model. Beneficial Microbes 9(1):87-100. |
|
|
Li J, Wang W, Xu SX, Magarvey NA, McCormick JK (2011). Lactobacillus reuteri-produced cyclic dipeptides quench agr-mediated expression of toxic shock syndrome toxin-1 in staphylococci. Proceedings of the National Academy of Sciences 108(8):3360-3365. |
|
|
Liu J, Gu Z, Song F, Zhang H, Zhao J, Chen W (2019). Lactobacillus plantarum ZS2058 and Lactobacillus rhamnosus GG use different mechanisms to prevent Salmonella infection in vivo. Frontiers in Microbiology 10: 299. |
|
|
Liu J, Hu D, Chen Y, Huang H, Zhang H, Zhao J, Gu Z, Chen W (2018). Strain-specific properties of Lactobacillus plantarum for prevention of Salmonella infection. Food and Function 9(7):3673-3682. |
|
|
Liu W, Ran C, Liu Z, Gao Q, Xu S, Ringø E, Myklebust R, Gu Z, Zhou Z (2016). Effects of dietary Lactobacillus plantarum and AHL lactonase on the control of Aeromonas hydrophila infection in tilapia. MicrobiologyOpen 5(4):687-699. |
|
|
Lixa C, Mujo A, Anobom CD, Pinheiro AS (2015). A structural perspective on the mechanisms of quorum sensing activation in bacteria. Anais da Academia Brasileira de Ciências 87(4):2189-2203. |
|
|
Marchelletta RR, Gareau MG, McCole DF, Okamoto S, Roel E, Klinkenberg R, Guiney DG, Fierer J, Barrett KE (2013). Altered expression and localization of ion transporters contribute to diarrhea in mice with Salmonella-induced enteritis. Gastroenterology 145(6):1358-1368. |
|
|
Markowiak P, Slizewska K (2017). Effects of probiotics, prebiotics, and synbiotics on human health. Nutrients 9(9):1021. |
|
|
Mathur H, Field D, Rea1 MC, Cotter PD, Hill C, Ross RP (2017). Bacteriocin-antimicrobial synergy: A medical and food perspective. Frontiers in Microbiology 8:1205. |
|
|
McFarland LV, Evans CT, Goldstein EJC (2018). Strain-specificity and disease-specificity of probiotic efficacy: A systematic review and meta-analysis. Frontiers in Medicine 5:124. |
|
|
Mendellin-Peña MJ, Griffiths MW (2009). Effect of molecules secreted by Lactobacillus acidophilus strain La-5 on Escherichia coli O157:H7 colonization. Applied and Environmental Microbiology 75(4):1165-1172. |
|
|
Meng CY, Smith BL, Bodhidatta L, Richard SA, Vansith K, Thy B, Srijan A, Serichantalergs O, Mason CJ (2011). Etiology of diarrhea in young children and patterns of antibiotic resistance in Cambodia. The Pediatric Infections Diseases Journal 30(4):331-335. |
|
|
Miquel S, Lagrafeuille R, Souweine B, Forestier C (2016). Anti-biofilm Activity as a Health Issue. Frontiers in Microbiology 7:592. |
|
|
Misselwitz B, Barrett N, Kreibich S, Vonaesch P, Andritschke D, Rout S, Weidner K, Sormaz M, Songhet P, Horvath P, Chabria M, Vogel V, Spori DM, Jenny P, Hardt WD (2012). Near surface swimming of Salmonella typhimurium explains target-site selection and cooperative invasion. PLoS Pathogens 8(7):e1002810. |
|
|
Moreira CG, Palmer K, Whiteley M, Sircili MP, Trabulsi LR (2006). Bundle-forming Pili and EspA are involved in biofilm formation by Enteropathogenic Escherichia coli. Journal of Bacteriology 188(11):3952-3961. |
|
|
Nakazato G, Paganelli FL, Lago JC, Aoki FH, Mobilon C, Brocchi M, Stehling EG, Silveira WD (2011). Lactobacillus acidophilus decreases Salmonella typhimurium invasion in vivo. Journal of Food Safety 31:284-289. |
|
|
Nascimento HH, Silva LEP, Souza RT, Silva NP, Scaletsky ICA (2014). Phenotypic and genotypic characteristics associated with biofilm formation in clinical isolates of typical enteropathogenic Escherichia coli (aEPEC) strains. BMC Microbiology 14(1):184. |
|
|
Navaneethan U, Giannella RA (2008). Mechanisms of infectious diarrhea. Nature Reviews Gastroenterology & Hepatology 5(11):637-647. |
|
|
Novik G, Savich V (2019). Beneficial microbiota. Probiotics and pharmaceutical products in functional nutrition and medicine. Microbes and Infection In Press. |
|
|
Oelschlaeger TA (2010). Mechanisms of probiotic action - A review. International Journal of Medical Microbiology 300(1):57-62. |
|
|
Oliveira NM, Martinez-Garcia E, Xavier J, Durham WM, Kolter R, Kim W, Foster KR (2015) Biofilm formation as a response to ecological competition. PLoS Biology 13(7):e1002191. |
|
|
Osama DM, Elkhatib WF, Tawfeik AM, Mohammad M (2017). Antimicrobial, antibiofilm and immunomodulatory activities of Lactobacillus rhamnosus and Lactobacillus gasseri against some bacterial pathogens. International Journal of Biotechnology for Wellness Industries 6(1):12-21. |
|
|
Paixão LA, Castro FFS (2016). A colonização da microbiota intestinal e sua influência na saúde do hospedeiro. Universitas: Ciências da Saúde 14(1):85-96. |
|
|
Piescik-Lech M, Urbanska M, Szajewska H (2013). Lactobacillus GG (LGG) and smectite versus LGG alone for acute gastroenteritis: a double-blind, randomized controlled trial. European Journal of Pediatrics 172(2):247-253. |
|
|
Plaza-Diaz J, Ruiz-Ojeda FJ, Gil-Campos M, Gil A (2019). Mechanisms of action of probiotics. Advances in Nutrition 10:S49-S66. |
|
|
Prabhurajeshwar C, Chandrakanth RK (2017). Probiotic potential of Lactobacilli with antagonistic activity against pathogenic strains: an in vitro validation for the production of inhibitory substances. Biomedical Journal 40(5):270-283. |
|
|
Quigley L, Coakley M, Alemayehu D, Rea MC, Casey PG, O'Sullivan OÌ, Murphy E, Kiely B, Cotter PD, Hill C, Ross RP (2019). Lactobacillus gasseri APC 678 reduces shedding of the pathogen Clostridium difficile in a murine model. Frontiers in Microbiology 10:273. |
|
|
Rabin N, Zheng Y, Opoku-Temeng C, Du Y, Bonsu E, Sintim HO (2015). Biofilm formation mechanisms and targets for developing antibiofilm agents. Future Medicinal Chemistry 7(4):493-512. |
|
|
Ramos AN, Sesto Cabral ME, Noseda D, Bosch A, Yantorno OM, Valdez JC (2012). Antipathogenic properties of Lactobacillus plantarum on Pseudomonas aeruginosa: The potential use of its supernatants in the treatment of infected chronic wounds. Wound Repair and Regeneration 20(4):552-562. |
|
|
Rätsep M, Kõljalg S, Sepp E, Smidt I, Truusalu K, Songisepp E, Stsepetova J, Naaber P, Mikelsaar RH, Mikelsaar M (2017). A combination of the probiotic and prebiotic product can prevent the germination of Clostridium difficile spores and infection. Anaerobe 47:94-103. |
|
|
Rosenvinge EC, Maldarelli GA, Piepenbrink KH, Scott AJ, Freiberg JA, Song Y, Achermann Y, Ernst RK, Shirtliff ME, Sundberg EJ, Donnenberg MS (2016). Type IV pili promote early biofilm formation by Clostridium difficile. FEMS Pathogens and Disease 74(6):ftw061. |
|
|
Ruiz MJ, Colello R, Padolaand NL, Etcheverría AI (2017). Efecto inhibitorio de Lactobacillus spp. sobre bactérias implicadas em enfermidades transmitidas por alimentos. Revista Argentina de Microbiologia 49(2):174-177. |
|
|
Saldaña Z, Erdem AL, Schüller S, Okeke IN, Lucas M, Sivananthan A, Phillips AD, Kaper JB, Puente JL, Girón JA (2009). The Escherichia coli common pilus and the bundle-forming pilus act in concert during the formation of localized adherence by enteropathogenic E. coli. Journal of Bacteriology 191(11):3451-3461. |
|
|
Sambanthamoorthy K, Feng X, Patel R, Patel S, Paranavitana C (2014). Antimicrobial and antibiofilm potential of biosurfactants isolated from lactobacilli against multi-drug-resistant pathogens. BMC Microbiology 14(1):197. |
|
|
Saxelin M, Lassig A, Karjalainen H, Tynkkynen S, Surakka A, Vapaatalo H, Järvenpää S, Korpela R, Mutanen M, Hatakka K (2010). Persistence of probiotic strains in the gastrointestinal tract when administered as capsules, yoghurt, or cheese. International Journal of Food Microbiology 144(2):293-300. |
|
|
Schiebel J, Böhm A, Nitschke J, Burdukiewicz M, Weinreich J, Ali A, Roggenbuck D, Rödiger S, Schierack P (2017). Genotypic and phenotypic characteristics associated with biofilm formation by human clinical Escherichia coli isolates of different pathotypes. Applied Environmental Microbiology 83(24):e01660-17. |
|
|
Sharman D, Saharan BS (2016). Functional characterization of biomedical potential of biosurfactant produced by Lactobacillus helveticus. Biotechnology Reports 11:27-35. |
|
|
Sikorska H, Smoragiewicz W (2013). Role of probiotics in the prevention and treatment of meticillin-resistant Staphylococcus aureus infections. International Journal of Antimicrobial Agents 42(6):475-481. |
|
|
Singh S, Singh SK, Chowdhury I, Singh R (2017). Understanding the Mechanism of Bacterial Biofilms Resistance to Antimicrobial Agents. The Open Microbiology Journal 11:53-62. |
|
|
Sniffen JC, McFarland LV, Evans CT, Goldstein EJC (2018). Choosing an appropriate probiotic product for your patient: An evidence-based practical guide. PLoS One 13(12):e0209205. |
|
|
Suez J, Zmora N, Zilberman-Schapira G, Mor U, Dori-Bachash M, Bashiardes S, Elinav E (2018). Post-antibiotic gut mucosal microbiome reconstitution is impaired by probiotics and improved by autologous FMT. Cell 174(6):1406-1423. |
|
|
Szajewska H, Canani RB, Guarino A, Hojsak I, Indrio F, Kolacek S, Orel R, Shamir R, Vandenplas Y, Van Goudoever JB, Weizman Z (2016). Probiotics for the Prevention of Antibiotic-Associated Diarrhea in Children. Journal of Pediatric Gastroenterology and Nutrition 62(3):495-506. |
|
|
Taverniti V, Koirala R, Via AD, Gargari G, Leonardis E, Arioli S, Guglielmetti S (2019). Effect of cell concentration on the persistence in the human intestine of four probiotic strains administered through a multispecies formulation. Nutrients 11(2):285. |
|
|
Ten Bruggencate SJ, Girard SA, Floris-Vollenbroek EG, Bhardwaj R, Tompkins TA (2015). The effect of a multi-strain probiotic on the resistance toward Escherichia coli challenge in a randomized, placebo-controlled, double-blind intervention study. European Journal of Clinical Nutrition 69:385-391. |
|
|
Ten Bruggencate SJM, Girard SA, Floris-Vollenbroek EGM, Bhardwaj R, Tompkins TA (2014). The effect of a multi-strain probiotic on the resistance toward Escherichia coli challenge in a randomized, placebo-controlled, double-blind intervention study. European Journal of Clinical Nutrition 69(3):385-391. |
|
|
Thiagarajah JR, Donowirtz M, Verkman AS (2015). Secretory diarrhoea: mechanisms and emerging therapies. Nature Reviews Gastroenterology & Hepatology 12(8):446-457. |
|
|
Tolker-Nielsen T (2015). Biofilm Development. Microbiology Spectrum 3(2):MB-0001-2014. |
|
|
Turková K, Mavric A, Narat M, Rittich B, Spanova A, Rogelj I, Matijasic BB (2013). Evaluation of Lactobacillus strains for selected probiotic properties. Folia Microbiologica 58(4):261-267. |
|
|
Vacheva A, Gerogieva R, Danova S, Mihova R, Marhova R, Kostadinova S, Vasileva K, Bivolarska M, Stoitsova SR (2012). Modulation of Escherichia coli biofilm growth by cell-free spent cultures from lactobacilli. Central European Journal of Biology 7(2):219-229. |
|
|
Varavallo MA, Thomé JN, Teshima E (2008). Aplicação de bactérias probióticas para profilaxia e tratamento de doenças gastrointestinais. Semina: Ciências Biológicas e da Saúde 29(1):83-104. |
|
|
Vieco-Saiz N, Belguesmia Y, Raspoet R, Auclair E, Gancel F, Kempf I, Drider D (2019). Benefits and inputs from lactic acid bacteria and their bacteriocins as alternatives to antibiotic growth promoters during food-animal production. Frontiers in Microbiology 10:57. |
|
|
Vlasova AN, Chattha KS, Kandasamy S, Liu Z, Esseili M, Shao L, Rajashekara G, Saif LJ (2013). Lactobacilli and Bifidobacteria promote immune homeostasis by modulating innate immune responses to human rotavirus in neonatal gnotobiotic pigs. PLoS One 8(10):e76962. |
|
|
Vuotto F, Longo F, Donelli G (2014). Probiotics to counteract biofilm-associated infections: promising and conflicting data. International Journal of Oral Science 6(4):189-194. |
|
|
Wolska KI, Grudniak AM, Rudnicka Z, Markowska K (2016). Genetic control of bacterial biofilms. Journal of Applied Genetics 57(2):225-238. |
|
|
World Gastroenteroly Organisation (WGO) (2017). World Gastroenterology Organization Global Guidelines - Probiotics and Prebiotics. |
|
|
World Health Organization (WHO) (2002). Guidelines for the Evaluation of Probiotics in Food. Londres. |
|
|
Wu H, Moser C, Wang Q, Hoiby N, Song Z (2015). Strategies for combating bacterial biofilm infections. International Journal of Oral Science 7(1):1-7. |
|
|
Yeganeh M, Hosseini H, Mehrabian S, Torbatti ES, Zamir AM (2017). Antibiofilm effects of lactobacilli against ciprofloxacin-resistant uropathogenic Escherichia coli strains in pasteurized milk. Applied Food Biotechnology 4(4):241-250. |
|
|
Zhang Z, Xiang Y, Li N, Wang B, Ai H, Wang X, Huang L, Zheng Y (2013). Protective effects of Lactobacillus rhamnosus GG against human rotavirus-induced diarrhoea in a neonatal mouse model. Pathogens and Disease 67(3):184-191. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


