Review

ABSTRACT
Filamentous fungi have been investigated as producer of xylanases with relevant characteristics for application in different industrial sectors, such as bakery, beverage, biofuel, textile, animal feed, pharmaceutical, pulp and paper. Thus, this review will focus on biochemical properties and industrial use of thermostable xylanases produced by different filamentous fungi, as well as mechanisms of adaptation of thermophilic organisms to tolerate in high-temperature environments. These enzymatic properties of thermal and pH stability are crucial, especially in processes such as the manufacture of animal feed, pulp and paper industry. Reports on changes in enzyme structure, such as site-directed mutagenesis, insertion or substitution of amino acids, addition of disulfide bonds in the alpha helix or beta-sheet structure for improving the thermal stability will also be reported. However, strains of Thermomyces lanuginosus has been described as good producers of thermostable xylanases, as well as promising enzymes, because it does not require any change in structure to increase the tolerance to high temperatures.
Key words: Hemicellulase, thermostability, Thermomyces.
INTRODUCTION
The global enzyme market achieved a value of $4.8 billion in 2013 and it is expected to achieve a value of $7.1 billion in 2018; this translates into an expected annual growth of 8.2% between 2013 and 2018 (BBC Research, 2014). Therefore, the industrial market demand for thermostable enzymes and the search for microbial sources that are easily accessible and produce high levels of enzymes have constituted a challenge for researchers over the years (Araújo et al., 2008; Haki and Rakshit, 2003). In addition, there are also some reports of efforts to improve the thermal stability of enzymes by recombinant DNA technology (Haltrich et al., 1996; Beg et al., 2001).
Microorganisms such as fungi and bacteria are known to produce different types of enzymes that can act on complex components in the plant cell wall and hydrolyzing them into smaller molecules (Badhan et al., 2007). The plant cell wall consists of three major components: cellulose (40%), hemicellulose (33%) and lignin (23%), whose composition varies according to the plant source (Carvalho et al., 2009; Dhiman et al., 2008). The hemicellulose is a heteropolysaccharide component of the plant wall; it primarily comprises xylan, whose main chain is composed of residues of D-xylose joined by (1 → 4)-β-glycosidic linkages and the side chains by L-arabinofuranose, D-glucuronic acid or 4-O-methyl-D-glucuronic acid (Wakiyama et al., 2008; Adsul et al., 2011). The complete depolymerization of xylan requires the synergic action of several enzymes: endo-β-1,4-xylanase (EC 3.2.1.8) and β-xylosidases (EC 3.2.1.37) to hydrolyze the main chain; acetylxylan esterases (EC 3.1.1.72), α-L-arabinofuranosidases (EC 3.2.1.55), ρ-coumaric acid esterases (EC 3.1.1.-), α-glucuronidases (EC 3.2.1.139), and feruloyl esterase (EC 3.1.1.7-) to act on the side chains (Masui et al., 2012).
The xylanases belong to the family of glycosyl hydrolases (GH), which catalyze the hydrolysis of 1,4-linked β-D-xylosidic linkages in the main chain of xylan (Collins et al., 2005; Heinen et al., 2014). Based on the similarities between their amino acid sequences and the hydrophobic groups of their catalytic domains, fungal xylanases are classified into GH10 and GH11 families (Verma and Satyanarayana, 2012). GH10 family groups xylanases with higher molecular weight (approximately 40 kDa) pI acid and three dimensional (β/α)8 structures, while the GH11 family have lower molecular weights (approximately 20 kDa) basic pI, and three-dimensional β barrel structures (Beaugrand et al., 2004; Georis et al., 2000). Xylanases of both families (GH10 and GH11) have two glutamate residues conserved in their active sites and may possess carbohydrate binding modules (CBMs) or amino- or carboxyl-terminal regions (Sydenham et al., 2014).
The use of microbial xylan-degrading enzymes, which are stable in alkaline pH and at high temperatures, in the biobleaching of pulp has increased because they help to reduce the consumption of chlorinated compounds (Goluguri et al., 2012). However, these enzymes should possess characteristics such as high specific activity, resistance to chemicals (metal cations), and absence of cellulase activity (Walia et al., 2014). Thus, this review will focus on thermophilic fungi producing xylanase and its industrial applications, as well as mechanisms of adaptation of thermophilic organisms to tolerate the high-temperature environments. In addition, biochemical properties of xylanase produced by strains of Thermomyces lanuginosus will be emphasized.
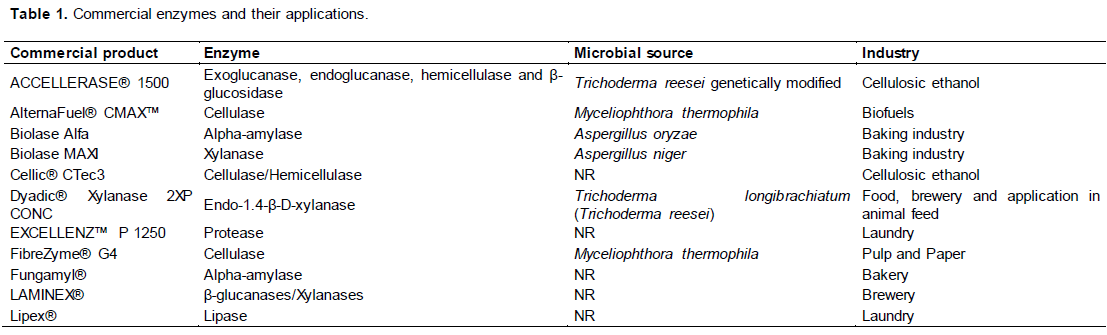
In this context, the isolation of new thermophilic species capable of producing thermostable enzymes is advantageous (in relation to mesophilic microorganisms) since these enzymes do not require any structural modification to resist higher temperatures (Benassi et al., 2014). Many companies have commercialized enzymes derived from fungi, mainly for application in different industrial sectors. Table 1 shows some commercial enzymes, their characteristics and the associated industries.
THERMOPHILIC MICROORGANISMS AND ADAPTATION TO TOLERATE HIGH-TEMPERATURE ENVIRONMENTS
Organisms are generally grouped according to their optimum temperature range for growth: psychrophilic (below 15°C), mesophilic (15 to 45°C), thermophilic (between 45 and 80°C) and (above 80°C) hyperthermophilic (Taylor and Vaisman, 2010). Among the species of thermophilic fungi are found T. lanuginosus and Talaromyces thermophilus, which grow at 50°C (Romdhane et al., 2010; Singh et al., 2003), and Rhizomucor miehei and Myceliophthora species, which grow at 45°C (Badhan et al., 2008; Fawzi, 2011).
Adaptations to tolerate adverse conditions such as extreme pH, high salt concentrations, and high temperatures are inherent in a few microorganisms. Among them, high temperatures have greater influence on the maintenance of biological structures and biomolecules functions, and they are related to changes in the composition of the cytoplasmic membrane, DNA and proteins (Gomes et al., 2007; de Oliveira et al., 2015).
The cytoplasmic membranes of thermophilic organisms consist of saturated fatty acids that confer increased stability and physical and functional integrity, unlike those of mesophilic organisms, which have unsaturated fatty acids (Gomes et al., 2007). The maintenance of DNA structure is an essential factor for all organisms, especially for hyperthermophilic which survive in high temperature environments. According to Mehta and Satyanarayana (2013), some factors may combine to provide thermal stability to DNA in thermophiles, including high levels of K+ and cyclic 2,3 diphosphoglycerate (2,3-DPG) that has been detected in the cytoplasm of thermophilic methanogens such as Methanothermus fervidus, Methanothermus sociabilis and Methanopyrus kandleri (Gomes et al., 2007; Scholz et al., 1992). Marguet and Forterre (1998) reported that 2,3-DPG prevents depurination or depyrimidation of DNA, which causes mutations at high temperatures. In addition, all hyperthermophilic organisms produce a different form of reverse DNA topoisomerase called DNA gyrase, which introduces positively supercoiled DNA. The positive supercoiling promotes greater DNA resistance to thermal denaturation (Gomes et al., 2007; López-García, 1999). The molecular chaperones are also important for thermophilic adaptation of microorganism. They result in the folding and refolding of proteins, preventing possibly irreversible protein denaturation (Conway and Macario, 2000). Nevertheless, the explanation of the mechanism involved in the survival of microorganisms, especially eukaryotes, at high temperatures is still not fully understood. On the other hand, there are many reports on biochemical characteristics of thermostable xylanases produced by thermophilic organisms (Table 2).
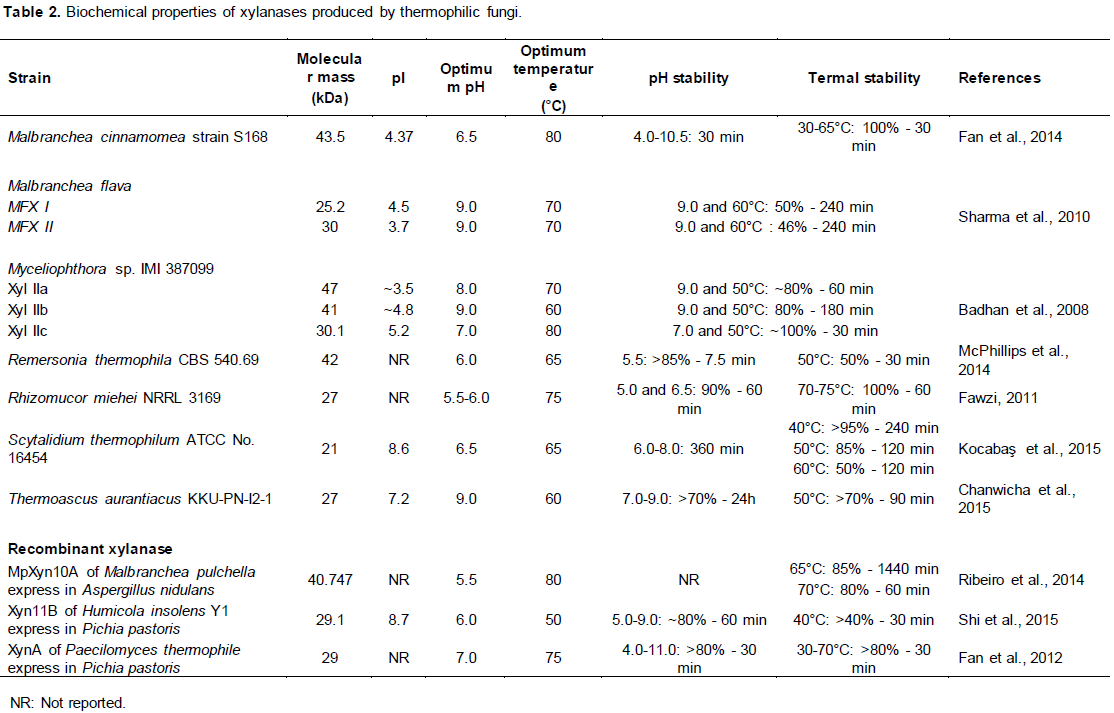
Xylanases derived from thermophilic fungi have exhibited molecular weights ranging from 21 to 47 kDa and a broad isoelectric point range of 3.5 to 8.7 (Table 2). There is a predominance of enzymatic activity in alkaline pH and high temperatures (up to 60°C). Most of the xylanases exhibit stability in a wide pH range (5.0 to 9.0), as Thermoascus aurantiacus KKU-PN-I2-1xylanase, which maintained activity in alkaline pH (9.0) for 24 h (Table 2). In addition, R. miehei NRRL 3169 xylanase showed high thermostability, retaining 100% of activity after 60 min of incubation at 70 to 75°C (Table 2).
THERMOSTABLE XYLANASE AND ITS PROPERTIES
Thermostability studies of some fungal enzymes have been described in the literature, but this attribute is still not fully understood. Higher thermal stability is one of the fundamental requirements for the application of an enzyme in industrial processes; it increases the efficiency of enzyme. Therefore, searching for thermostable enzymes or improve the thermostability of enzymes has been the priority for researchers over the years (Nirmal and Laxman, 2014). The factors affecting thermostability are important for understanding the functions of proteins and their use in various industries (Ruller et al., 2008). The advantages of employing enzymes with high optimum temperatures in biotechnological processes or biocatalytic conversions industrial include the lower risk of microbial contamination by common mesophiles, the improvement of substrate solubility, increased reaction rates and decreased viscosity (Joo et al., 2011; Haki and Rakshit, 2003). The tolerance of the enzymes to high temperatures for long periods may be associated with their conformational structures, composition and/or amino acid sequences, and the origins of the enzymes (Gomes et al., 2007; Techapun et al., 2003). In general, a strategy for obtaining thermostable enzymes is search in organisms that grow in high temperature environments because their enzymes are more thermostable than those from mesophilic (Techapun et al., 2003). Several authors have reported thermostable xylanases obtained from thermophilic fungi, such as, Humicola brevis var. thermoidea (Masui et al., 2012), Paecilomyces themophila J18 (Yang et al., 2006) and T. aurantiacus RCKK (Jain et al., 2015). Most of these fungal xylanases thermostable belong to GH10 and GH11 family, and enzymes belonging to the same family, they present the same structure and amino acid sequence, and thus the thermal stability also did not differ (Collins et al., 2005; You et al., 2010).
It is also possible to obtain thermostable enzymes by the improvement of characteristics through small alteration in enzyme structure. These include site-directed mutagenesis (Wang et al., 2014; Zheng et al., 2014; Xie et al., 2011), replacing the N-terminal region of a mesophilic xylanase by N-terminal region of thermophilic organisms (Zhang et al., 2010) and the addition of disulfide bridges in the N-terminal region of the α- helix. Hakulien et al. (2003) analyzed crystal
structures of 12xylanases belonging to family 11 (GH11), and concluded that the xylanase structures of mesophilic and thermophilic organisms were similar and that minor modifications altered thermostability. Such modifications included larger numbers of amino acid residues in beta sheets and the stabilization of the alpha-helix region; higher proportions of threonine:serine; increasing charged amino acid residues such as arginine, leading to an improvement in polar interactions; more compact structures and more aromatic residues or pairs of ions in protein surfaces. Also, according to You et al. (2010), the replacement of the cysteine at site 201 of the xylanase (GH11) improved its thermostability due to the strong hydrophobic interaction with the cysteine at site 50. Therefore, disulfide bond formation and hydrophobic interactions contributed to the rise in the thermal stability of xylanase. Similarly, Song and colleagues (2015) reported the importance of the amino acid of the N-terminal region of the Aspergillus niger (Xyn10A_ASPNG) xylanase for improving its thermostability, and the thermal inactivation half-life (t1/2) at 60°C was prolonged by 30 times in comparison with wild-type enzyme.
APPLICATION OF THERMOSTABLE XYLANASES IN INDUSTRIAL PROCESSES
The application of enzymes in industry has grown over the years. Some enzymes, such as xylanases, lipases, cellulases, proteases, amylases and phytases, have been used in feed industry to reduce the viscosity of the food and improve the absorption of nutrients in the digestive tracts of animals (Polizeli et al., 2005). These enzymes may act when the feeds are being processed (for example, when thermostable xylanases are added before the pelletization process (70-95°C), transported, and stored; they can also facilitate digestion in the gastrointestinal tract of the animal (Pariza and Cook, 2010; Collins et al., 2005).
Furthermore, xylanases are used in the food industry. For example, in the production of bread, enzymes are added to improve its softness and shelf life (Sharma and Kumar, 2013). A study by Jiang et al. (2005) evaluated the effect of xylanase purified from T. lanuginosus CAU44 on the quality of bread and its staling rate during storage and reported that thermostable xylanase could be used in the bakery industry. Similarly, the potential of xylanase XYNZG from Plectosphaerella cucumerina for baking by heterologously expressed in Kluyveromyces lactis was studied. They obtained improvement in sensorial characteristics, volume, texture, and handling time (6.5 to 6.0 min) (Zhan et al., 2014). The purified xylanase from Remersonia thermophila CBS 540.59 (Rtl) also showed increased loaf volume by 8%, softness (19.6%) and decreased in 20.4% hardening of the bread after four days of storage compared to the control (McPhillips et al., 2014). The enzyme cocktail that the thermophilic fungus T. aurantiacus (CBMAI 756) produced was used (35 U of xylanase/100 g of flour), resulting in the increase of loaf volume (by 22%), the reduction in crumb firmness (by 25%), and amylopectin retrogradation (by 17%) (Oliveira et al., 2014).
Xylanases are also used in the beverage industry together with pectinases, cellulases and amylases, which act to recover flavor, reduce viscosity and turbidity, and stabilize fruit pulp (Polizeli et al., 2005). Thermoacidophilic xylanase from Penicillium pinophilum C1 was employed in the brewing industry. The enzyme has improved the filtration rate to 22.3% and the viscosity of wort to 5.0% with 40 U of xylanase (purified XYN10C1) (Cai et al., 2011).
Moreover, the addition of a higher dose of the enzyme (80 U) improved these parameters, resulting in the filtration rate to 26.7% and the viscosity of wort to 9.8%. Similarly, the recombinant xylanase from Gloeophyllum trabeum reduced the specific filtration rate and viscosity of wort to 17.2 and 7.1%, respectively using 40 U of GtXyb10 enzyme, but it was more effective with 80 U of the enzyme, reducing the filtration rate to 31.3% and the viscosity of wort to 12.8% (Wang et al., 2016).
In the pulp and paper industries, thermophilic, alkalophilic, and cellulase-free xylanases have been used in the biobleaching of pulp. They facilitate the depolymerization of xylan, leading to the formation of pores and giving chlorinated reagents the necessary access for the removal of lignin from the wood pulp. Thus, these enzymes provide eco-friendly alternative for the effective bleaching of pulp, reducing the use of toxic chlorine compounds (Christopher et al., 2005; Subramaniyan and Prema, 2000; Kanwar and Devi, 2012; Sharma et al., 2015). Thus, several studies have focused on the use of enzymes in the pretreatment of pulp. For instance, the cellulase-free xylanase from Trichoderma viride was used on kraft pulp from Eucalyptus grandis, resulting in a decrease in its Kappa number and its maintenance of viscosity compared to those of the control, with parameters (enzyme dose, time, temperature, and pH). The results indicated that the enzyme showed potential for use in the pulp and paper industry (Fortkamp and Knob, 2014). Similar studies by Guimarães et al. (2013) using a xylanase from Aspergillus aculeatus var aculeatus in the pretreatment of E. grandis pulp, and by Silva et al. (2016) described the effective use of xylanase from Penicillium crustosum to bleach the kraft pulp of E. grandis, obtained a significant reduction in Kappa number (5.27 points) corresponding to a 35.04% Kappa efficiency. Although, there are some reports in the literature on the use of thermostable microbial xylanases, especially in the paper industry, there is still a deficit of enzymes with stability at high temperatures and alkaline pH. Thus, the search for new strains of fungi capable of producing thermostable enzymes at high temperatures and alkaline pH remains relevant.
Furthermore, alkaline xylanases are used in the detergent industry. They are additives in the formulation of detergents and, thus, improve the removal of stains of vegetable origin (Kamal et al., 2004). In the textile, enzymes act on cotton cleaning (cotton biopolishing), in order to improve the physical characteristics of the tissue, to assist in the removal of non-cellulosic materials and facilitate alkaline extraction step to improve the access of compounds chemicals to the fiber, leading to a reduction in consumption of these agents and possible environmental damage (Battan et al., 2012; Csiszár et al., 2006).
T. lanuginosus
T. lanuginosus is a thermophilic filamentous fungus, synonymous with Humicola lanuginosa. The Thermomyces genus includes four species: T. lanuginosus Tsiklinsky, which was first isolated from garden soil in 1899 (Pugh et al., 1964); Thermomyces ibadanensis Apinis & Eggins, a thermophilic and lipolytic fungus isolated from palm fruit in 1966 (Apinis and Eggins, 1966); Thermomyces stellatus (Bunce) Apinis, a thermophilic species found in hay 1961 (Bunce, 1961); and Thermomyces verrucosus Pugh, Blakeman and Morgan-Jones, a mesophilic species discovered in 1964 (Pugh et al., 1964). T. lanuginosus grows at temperatures above 45°C and is initially white in color, becoming dark brown after maturation (Khucharoenphaisan and Sinma, 2010).
There are many reports of strains of T. lanuginosus with high ability to produce thermostable enzymes using inexpensive carbon sources such as sorghum straw (Singh et al., 2000; Sonia et al., 2005) and corn cobs (Winger et al., 2014). These alternative carbon sources can reduce the investment in the production of enzymes. Moreover, the reuse of agricultural waste can contribute to ecologically correct efforts to reduce the disposal of materials in the environment and convert raw materials into valuable products such as biofuels (Howard et al., 2003).
T. lanuginosus is an interesting fungus because it can produce enzymes of industrial interest such as invertase (Chaudhuri and Maheshwari, 1996), β-xylosidase (Corrêa et al., 2016), chitinase (Khan et al., 2015; Chen et al., 2012; Zhang et al., 2015), protease (Li et al., 1997), inulinase (Flores-Gallegos et al., 2015), esterase (Li et al., 2014), amylase (Kunamneni et al., 2005), glucoamylase (Gonçalves et al., 2008), α-galactosidase (Rezessy-Szabó et al., 2007), and the two most reported enzymes, lipase (Wang et al., 2015; Ávila-Cisneros et al., 2014; Fang et al., 2014) and xylanase (Jiang et al., 2015; Stephens et al., 2014; Shrivastava et al., 2013; Manimaran et al., 2009). Furthermore, T. lanuginosus is known for producing cellulase-free xylanases (Manimaran et al., 2009; Li et al., 2005; Damaso et al., 2002). This characteristic free cellulase is essential for the application of xylanase in pulp and paper industry because it prevents degradation of the cellulose (Beg et al., 2001). Furthermore, xylanases from T. lanuginosus has been reported to belong to family 11 glycoside hydrolases (Wang et al., 2012; Gruber et al., 1998).
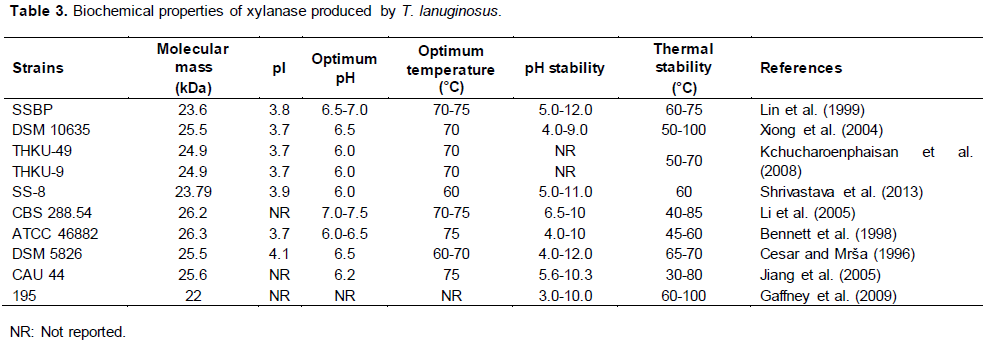
According to Gruber et al. (1998), the crystal structure of the T. lanuginosus xylanase is a compact globular protein consisting of two highly twisted β-sheets and one α-helix, and it is surrounded by water molecules on its surface as well as one (O201) on the inside. Moreover, the enzyme has a disulfide bond that is absent in most other xylanases of the GH11 family. This could be related to electrostatic interaction through the ion pairs and could explain their tolerance to higher temperatures. Table 3 summarizes some biochemical characteristics of xylanases from different strains of T. lanuginosus.
The xylanases from different strains of T. lanuginosus exhibit low molecular weights (22 to 26 kDa), facilitating their passage through hemicellulose networks and resulting in the improvement of enzymatic hydrolysis (Juturu and Wu, 2012), enzymatic activity at high temperatures (60 to 75°C), and thermal stability (30 to 100°C). This ensures that they can be used at high temperatures and remain stable in a wide pH range (3 to 12), making them relevant in different biotechnological processes (Chen et al., 2014; Mamo et al., 2009).
CONCLUSION
Despite the biotechnological advances of recent decades, most studies have focused on the production, purification, biochemical characterization, and regulation of thermostable fungal xylanases. Although there are some reports in the literature regarding the use of thermostable microbial xylanases, especially in the paper industry, they lack stability at high temperatures and in alkaline pH. Some studies have shown that small changes in enzyme structure, through site-directed mutagenesis, insertion or substitution of amino acids, or addition of disulfide bridges to stabilize alpha-helix structures or beta sheets, result in the improved thermal stability of the enzyme. On the other hand, there are many strains of T. lanuginosus that are good producer of thermostable xylanases with activity at high temperatures (60 to 75°C) and in a wide range of thermal stability (30 to 100°C). Xylanases with this property would be particularly relevant and advantageous to detergents industry and pulp and paper, because it does not require any change in structure to increase the tolerance to high temperatures.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
Carla Lieko Della Torre is recipient of fellowship from Brazilian Federal Agency for Support and Evaluation of Graduate Education (CAPES)/Fundação Araucária, Brazil.
REFERENCES
|
Adsul MG, Singhvi MS, Gaikaiwari SA, Gokhale DV (2011). Development of biocatalysts for production of commodity chemicals from lignocellulosic biomass. Biores. Technol. 102(6):4304-4312. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Apinis AE, Eggins HOW (1966). Thermomyces Ibadanensis sp.nov. from oil palm kernel stacks in Nigeria. T. Brit. Mycol. Soc. 49(4):629-632. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Araújo R, Casal M, Cavaco-Paulo A (2008). Application of enzymes for textile fibres processing. Biocatal. Biotransfor. 26(5):332–349. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Ávila-Cisneros N, Velasco-Lozano S, Huerta-Ochoa S, Córdova-López J, Gimeno M, Favela-Torres M (2014). Production of thermostable lipase by Thermomyces lanuginosus on solid-state fermentation: selective hydrolysis of sardine oil. Appl. Biochem. Biotechnol. 174(5):1859-1872. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Badhan AK, Chadha BS, Kaur J, Saini HS, Bhat MK (2007). Production of multiple xylanolytic and cellulolytic enzymes by thermophilic fungus Myceliophthora sp. IMI 387099. Bioresour. Technol. 98(3):504-510. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Badhan AK, Chadha BS, Saini HS (2008). Purification of the alkaliphilic xylanases from Myceliophthora sp. IMI 387099 using cellulose-binding domain as an affinity tag. World J. Microbiol. Biotechnol. 24(7):973-981. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Battan B, Dhiman SS, Ahlawat S, Mahajan R, Sharma J (2012). Application of thermostable xylanase of Bacillus pumilus in textile processing. Indian J. Microbiol. 52(2):222-229. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
BCC Research: Global markets for enzymes in industrial applications. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Beaugrand J, Chambat G, Wong VW, Goubet F, Rémond C, Paës G, Benamrouche S, Debeire P, O'Donohue M, Chabbert B (2004). Impact and efficiency of GH10 and GH11 thermostable endoxylanases on wheat bran and alkali-extractable arabinoxylans. Carbohydr. Res. 339(15):2529-2540. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Beg QK, Kapoor M, Mahajan L, Hoondal GS (2001). Microbial xylanases and their industrial applications: a review. Appl. Microbiol. Biotechnol. 56(3-4):326-338. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Benassi VM, Lucas RC, Jorge JA, Polizeli MLTM (2014). Screening of thermotolerant and thermophilic fungi aiming β-xylosidase and arabinanase production. Braz. J. Microbiol. 45(4):1459-1467. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Bennett NA, Ryan J, Biely P, Vrsanska M, Kremnicky L, Macris BJ, Kekos D, Christakopoulos P, Katapodis P, Claeysses M, Nerinckx W, Ntauma P, Bhat MK (1998). Biochemical and catalytic properties of an endoxylanase purified from the culture filtrate of Thermomyces lanuginosus ATCC 46882. Carbohydr. Res. 306(3):445-455. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Bunce ME (1961). Humicola Stellatus sp.nov., a thermophilic mould from hay. T. Brit. Mycol. Soc. 44(3):372-385. Doi:10.1016/S0007-1536(61)80031-4. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Cai H, Shi P, Bai y, Huang H, Yuan T, Yang P, Luo H, Meng K, Yao B (2011). A novel thermoacidophilic family 10 xylanase from Penicillium Pinophilum C1. Process Biochem. 46(12):2341-2346. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Carvalho W, Canilha L, Ferraz A, Milagres AMF (2009). Uma visão sobre a estrutura, composição e biodegradação da madeira. Quim. Nova. 32(8):2191-2195. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Cesar T, Mrša V (1996). Purification and properties of the xylanase produced by Thermomyces lanuginosus. Enzyme Microb. Technol. 19(4):289-296. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Chanwicha N, Katekaew S, Aimi T, Boonlue S (2015). Purification and characterization of alkaline xylanase from Thermoascus Aurantiacus var. levisporus KKU-PN-I2-1 cultivated by solid-state fermentation. Mycoscience 56(3):309-318. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Chaudhuri A, Maheshwari R (1996). A novel invertase from a thermophilic fungus Thermomyces lanuginosus: its requirement of thiol and protein for activation. Arch. Biochem. Biophys. 327(1):98-106. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Chen CC, Luo H, Han X, Lv P, Ko TP, Peng W, Huang CH, Wang K, Gao J, Zheng Y, Yang Y, Zhang J, Yao B, Guo RT (2014). Structural perspectives of an engineered β-1,4-xylanase with enhanced thermostability. J. Biotechnol. 189:175-182. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Chen Z, Jia H, Yang Y, Yan Q, Jiang Z, Teng C (2012). Secretory expression of a β-xylosidase gene from Thermomyces lanuginosus in Escherichia Coli and characterization of its recombinant enzyme. Lett. Appl. Microbiol. 55(5):330-337. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Christopher L, Bissoon S, Singh S, Szendefy J, Szakacs G (2005). Bleach-enhancing abilities of Thermomyces lanuginosus xylanases produced by solid state fermentation. Process Biochem. 40(10):3230-3235. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Collins T, Gerday C, Feller G (2005). Xylanases, xylanase families and extremophilic xylanases. FEMS Microbiol. Rev. 29(1):3-23. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Conway M, Macario AL (2000). Stressors, stress and survival: overview. Front. Biosci. 5:780-786. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Corrêa JM, Christi D, Torre CLD, Henn C, Conceição-Silva JL, Kadowaki MK, Simão RCG (2016). High levels of β-xylosidase in Thermomyces Lanuginosus: potential use for saccharification. Braz. J. Microbiol. 47(3):680-690. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Csiszár E, Losonczi A, Koczka B, Szakács G, Pomlényi A (2006). Degradation of lignin-containing materials by xylanase in biopreparation of cotton. Biotechnol. Lett. 28(10):749-753. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Damaso MCT, Andrade CMMC, Pereira N (2002). Production and properties of the cellulase-free xylanase from Thermomyces Lanuginosus IOC-4145. Braz. J. Microbiol. 33:333-338. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
de Oliveira TB, Gomes E, Rodrigues A (2015). Thermophilic fungi in the new age of fungal taxonomy. Extremophiles 19(1):31-37. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Dhiman SS, Sharma J, Battan B (2008). Industrial applications and future prospects of microbial xylanases: a review. BioResources 3(4):1377-1402. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Fan G, Katrolia P, Jia H, Yang S, Yan Q, Jiang Z (2012). High-Level expression of a xylanase gene from the thermophilic fungus Paecilomyces Thermophila in Pichia Pastoris. Biotechnol. Lett 34(11):2043-2048. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Fan G, Yang S, Yan Q, Guo Y, Li Y, Jiang Z (2014). Characterization of a highly thermostable glycoside hydrolase family 10 xylanase from Malbranchea Cinnamomea. Int. J. Biol. Macromol. 70:482-489. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Fang Z, Xu L, Pan D, Jiao L, Liu Z, Yan Y (2014). Enhanced production of Thermomyces Lanuginosus lipase in Pichia Pastoris via genetic and fermentation strategies. J. Ind. Microbiol. Biot. 41(10):1541-1551. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Fawzi E (2011). Highly thermostable xylanase purified from Rhizomucor miehei NRL 3169. Acta Biol. Hung. 62(1):85-94. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Flores-Gallegos AC, Contreras-Esquivel JC, Morlett-Chávez JA, Aguilar CN, Rodríguez-Herrera R (2015). Comparative study of fungal strains for thermostable inulinase production. J. Biosci. Bioeng. 119(4):421-426. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Fortkamp D, Knob A (2014). High xylanase production by Trichoderma viride using pineapple peel as substrate and its apllication in pulp biobleaching. Afr. J. Biotechnol. 13(22):2248-2259. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Gaffney M, Carberry S, Doyle S, Murphy R (2009). Purification and characterisation of a xylanase from Thermomyces lanuginosus and its functional expression by Pichia pastoris. Enzyme Microb. Technol. 45 (5): 348–354. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Georis J, de Lemos Esteves F, Lamotte-Brasseur J, Bougnet V, Devreese B, Giannotta F, Granier B, Frère JM (2000). An additional aromatic interaction improves the thermostability and thermophilicity of a mesophilic family 11 xylanase: structural basis and molecular study. Protein Sci. 9 (3):466-475. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Goluguri BR, Thulluri C, Cherupally M, Nidadavolu N, Achuthananda D, Mangamuri LN, Addepally U (2012). Potential of thermo and alkali stable xylanases from Thielaviopsis Basicola (MTCC-1467) in biobleaching of wood kraft pulp. Appl. Biochem. Biotechnol. 167(8):2369-2380. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Gomes E, Guez MAU, Martin N, Silva R (2007). Enzimas termoestáveis: fontes, produção e aplicação industrial. Quim. Nova 30(1):136-145. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Gonçalves AZL, Carvalho AFA, Silva R, Gomes E (2008). Localization and partial characterization of thermostable glucoamylase produced by newly isolated Thermomyces Lanuginosus TO3 in submerged fermentation. Braz. Arch. Biol. Technol. 51(4):857-865. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Gruber K, Klintschar G, Hayn M, Schlacher A, Steiner W, Kratky C (1998). Thermophilic xylanase from Thermomyces Lanuginosus: high-resolution x-ray structure and modeling studies. Biochemistry 37(39):13475-13485. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Guimarães NCA, Sorgatto M, Peixoto-Nogueira SC, Betini JHA, Zanoelo FF, Marques MR, Polizeri MLTM, Giannesi GC (2013). Xylanase production from Aspergillus Japonicus var aculeatus: production using agroindustrial residues and biobleaching effect on pulp. J. Biocatal. Biotransform. 2(1):1-6. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Haki GD, Rakshit SK (2003). Developments in industrially important thermostable enzymes: a review. Bioresour. Technol. 89(1):17-34. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Hakulinen N, Turunen O, Janis J, Leisola M, Rouvinen J (2003). Three-dimensional structures of thermophilic beta-1,4-xylanases from Chaetomium thermophilum and Nonomuraea flexuosa. comparison of twelve xylanases in relation to their thermal stability. Eur. J. Biochem. 270(7):1399-1412. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Haltrich D, Nidetzky B, Kulbe KD, Steiner W, ŽupanÄiÄ S (1996). Production of fungal xylanases. Bioresour. Technol. 58(2):137-161. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Heinen PR, Henn C, Peralta RM, Bracht A, Simão RCG, Conceição Silva JL, Polizeli MLTM, Kadowaki MK (2014). Xylanase from Fusarium heterosporum: properties and influence of thiol compounds on xylanase activity. Afr. J. Biotechnol. 13(9):1047-1055. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Howard RL, Abotsi E, Jansen REL, Howard S (2003). Lignocellulose biotechnology: issues of bioconversion and enzyme production. Afr. J. Biotechnol. 2(12):602-619.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


