Full Length Research Paper

ABSTRACT
Inoculation of promiscuous soybean with rhizobia has been proven to improve yield. Screenhouse pot experiments were conducted to assess the response of promiscuous soybean, TGx1448-2E to rhizobia inoculation and to determine the relative efficiency of indigenous rhizobia from soils collected from the Northern and Southern Guinea and Sudan Savanna agro-ecological zones of Nigeria. Soils were collected at 0 to 20 cm depth from 45 sites spread in 15 local government areas of Kaduna and Kano states. The treatments used were uninoculated control, mineral nitrogen and Legumefix, a commercial rhizobia inoculant which were arranged in randomized complete block design and replicated three times. Nodulation and biomass were assessed at eight weeks after planting. Response to inoculation and relative efficiency were also determined. The results showed that response to inoculation was higher in the Sudan Savanna than in the Northern Guinea Savanna. These were observed in 12 out of 21 and 9 out of 19 sites in the Sudan Savanna and Northern Guinea Savanna, respectively. Of these, inoculation response reached 62 and 90% in two sites in the Sudan Savanna while it barely reached 20% in the remaining sites. The results also showed a relative efficiency which is greater than 100% in some locations. This implies that the indigenous rhizobia populations were superior to mineral nitrogen and Legumefix inoculant. Notable among the locations were ALB 1, GRK 1, SOB 4 and KOL, which spread across all the agro-ecological zones. Strains from such locations have high potential for use as inoculants.
Key words: Legumefix, mineral nitrogen, inoculation, indigenous strains, agro-ecological zones.
INTRODUCTION
Soybean can contribute to the N economy of nutrient-poor soils through its capacity to form symbiotic relationship with different rhizobia species. It is estimated that rhizobia can fix about 50 to 300 kg N ha-1 (Bokhtiar and Sakurai, 2005). Thus, their contribution to the N economy of the soil can be quite substantial. Soybean has the capability to improve soil fertility in cereal based cropping systems in the Guinea Savanna (Yusuf et al., 2006). It can fix as much as 100 kilograms of nitrogen per hectare (Sanginga et al., 2003).
In the early 1980s, it was assumed that most tropical countries did not have the facilities and personnel required for inoculum production, its storage, and distribution. They depended on the importation of the crop’s final product (Pulver et al., 1982). Soybean breeders at the International Institute of Tropical Agriculture (IITA), Nigeria developed promiscuous soybean genotypes in order to take advantage of high yielding American and promiscuous Asian soybean varieties that were introduced to the country. These genotypes, known as Tropical Glycine cross (TGx) nodulate effectively with Bradyrhizobium spp. populations that are indigenous to African soils (Abaidoo et al., 2007). Okereke et al. (2000) reported that promiscuous soybean may also need to be inoculated with exotic Bradyrhizobia depending on the effectiveness and population of the indigenous bradyrhizobia in the locality.
Furthermore, in order to ensure optimum productivity by these promiscuous genotypes, it is essential to assess the relative effectiveness of the indigenous rhizobia in the soil. The number of indigenous Bradyrhizobia as well as the soil available nitrogen has been cited as some of the main factors that have a significant effect on symbiotic relationships (Keyser and Li, 1992; Maingi et al, 2006). Sanginga et al. (1999) found that the need for inoculation of promiscuous soybean depended on the effectiveness of indigenous Bradyrhizobia population. An increase in nodulation and N2 fixation of promiscuous soybean cultivars using commercial Bradyrhizobium inoculant was reported by Okereke and Eaglesham (1993) and Yusuf et al. (2012), while Wasike et al. (2009) reported nodulation of promiscuous soybean varieties with indigenous rhizobia in Western and Eastern Kenya. Most of the studies conducted in Nigeria are limited in terms of number and location. They are few studies mostly conducted on-station.
This study was conducted to assess the response of promiscuous soybean to inoculation in soils of the moist Savanna of Nigeria and also to examine the relative effect of the indigenous rhizobia population on soybean biomass. The present investigation attempts to expand on the existing studies on the response of promiscuous soybean to inoculation in these agro-ecologies.
MATERIALS AND METHODS
Study location
The Northern Guinea Savanna of Nigeria covers an area of about 34 million hectares in West and Central Africa (Kolawole et al, 2007). Its temperatures range from 27.3-34.0oC (maximum) and 18.6-21.6°C (minimum). The mean annual rainfall of the area is between 1200 and 1700 mm, with a growing period between May and October (Wall, 1979). The Southern Guinea Savanna is characterized by a mean annual rainfall of 1284mm and a maximum temperature of about 33.5°C (Ojanuga, 2006). The Sudan Savanna zone covers about 22.8 million hectares (Manyong et al., 1995), and it has high annual average temperature of 28 to 32°C (Sowunmi and Akintola, 2010) and annual average rainfall of 884 to 1200 mm (Shehu et al., 2015).
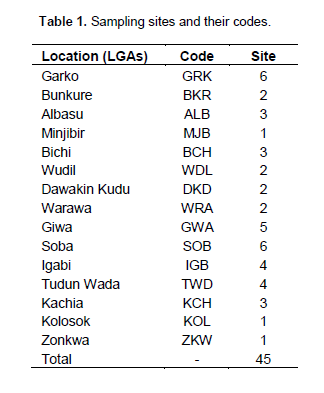
The experiment was set up in a screenhouse in the Department of Soil Science, Ahmadu Bello University, Zaria (110 9’ N, 70 36’ E). Soil samples were collected at 0 to 20 cm depth from 45 sites in 14 local government areas (LGAs) spread across three agro-ecological zones of Nigeria (Table 1). Figure 1 shows a map of the sampling distribution. The samples were bulked, air dried, crushed and sieved through a 4 mm mesh (for planting) and 2 mm and 0.5 mm mesh (for physicochemical analysis). Plastic pots of about 16.5 cm in diameter and 16.5 cm height with a capacity of 3.5 litres and drainage holes were filled with 4 kg of soil samples.
The trial was made up of 3 treatments. The first treatment had mineral nitrogen otherwise known as the reference; the second treatment had a known commercial inoculant (Legumefix) which contained Bradyrhizobium japonicum USDA 532c while the third treatment was an uninoculated control with no N applied. All treatments were replicated three times and arranged in Randomized Complete Block Design (RCBD) due to shading effect of trees around the screenhouse.
The following fertilizer treatments were applied to all pots to ensure adequate nutrient supply:
ZnSO4 (0.079 mg/pot), MgSO4 (0.085 mg/pot), TSP (0.507 mg/pot), NaMoO4.2H2O (0.004 mg/pot), KCl (0.117mg/pot) and Urea (0.263 mg in each of the relevant pots).
The urea was applied in split form: the first dose at planting and the second dose 14 days after planting. The test crop used was a promiscuous soybean cultivar (TGx1448-2E). The seeds were surface sterilized to rid them of contamination especially from rhizobia as described by Somasegaran and Hoben (1985). Five seeds were planted per pot and then later thinned to two plants per pot at two weeks after planting. The plants were irrigated once or twice daily depending on the moisture content of the soil.
Soil analysis
Particle size distribution was determined by the hydrometer method, as described by Gee and Bauder (1986). The soil texture was determined using the USDA soil textural triangle. Soil pH was measured in both water and 0.01 M CaCl2 solution using a 1:2.5 soil to solution ratio (Hendershot et al., 1993).
Organic carbon was determined by the Walkey-Black (chromic acid) method as described by Nelson and Sommers (1982). Total nitrogen was determined by micro-Kjeldahl digestion method (Bremner and Mulvaney, 1982). Available phosphorus was determined by the Bray 1 method (Olson and Sommers, 1982). Exchangeable bases were extracted with 1N ammonium acetate at pH 7.0 buffer (Chapman, 1965). Exchangeable K and Na was determined using flame photometer while exchangeable Ca and Mg was determined using Atomic Adsorption Spectrophotometer (AAS) (Jackson, 1958). The soil CEC was determined by 1N ammonium acetate saturation method (Anderson and Ingram, 1993) (Table 1 and Figure 1).
At eight weeks after planting, the plants were harvested and the following parameters were assessed: nodule number, nodule fresh weight, shoot dry weight and root dry weight. The data used for the soil analysis were subjected to descriptive statistics. The total dry matter yield (TDMY) was used to calculate the relative effectiveness index which gives an indication of the relative efficiency (RE) and inoculant response (IR) for each of the agro-ecological zones.
Relative Efficiency (RE) = 100a/b
Inoculant Response (IR) = 100 (c-a)/a
Where:
a = dry matter weight in control treatment
b = dry matter weight in plus N treatment
c = dry matter weight of inoculated treatment
RESULTS AND DISCUSSION
Physicochemical properties of soil samples
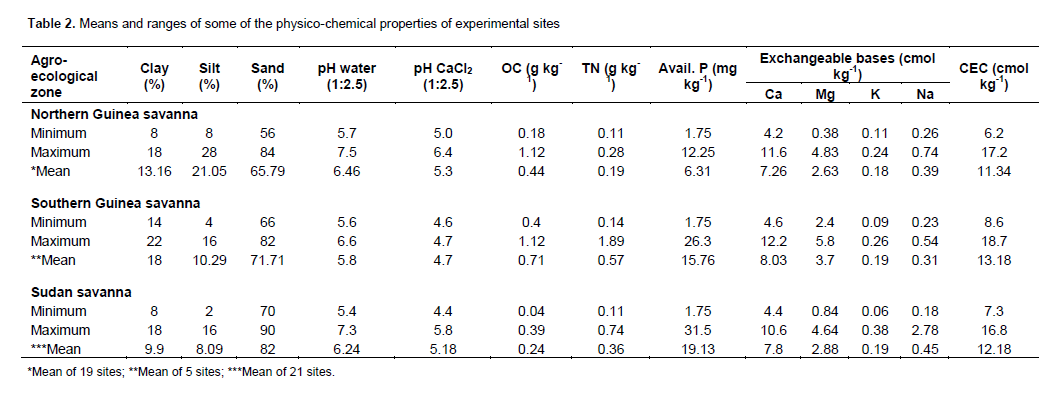
The soils of the experimental areas are generally slightly acidic to moderately acidic (Table 2). This is a common feature of savanna soils (Jones and Wild, 1975). The pH value falls within the normal range of 5.5-7.0 reported to be optimum for the release of plant nutrients (Sharu et al., 2013). The organic carbon total nitrogen contents were generally low for all the agro-ecologies. Jones and Wild (1975) have earlier reported low values of available P in addition to organic carbon and total nitrogen in Nigerian Savanna soils. These ratings are according to FMANR (1990).
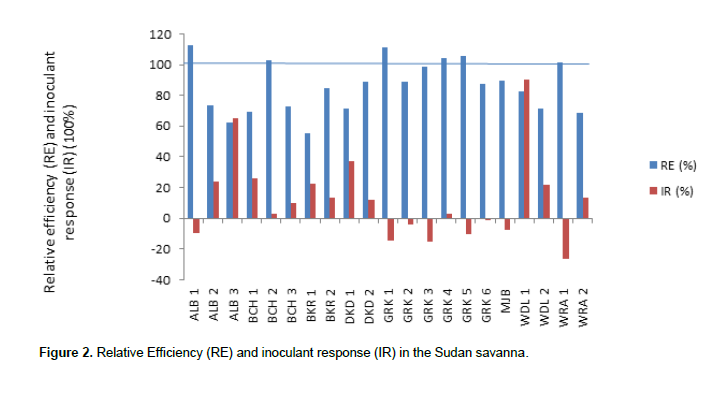
Relative efficiency (RE) and inoculant response (IR) in the three agro-ecological zones
In the Sudan Savanna (Figure 2), the IR ranged from -26.15 to 90.5%, while the RE ranged from 55.06 to 112.5%. The figure shows that WDL 1 has the highest response to inoculation (90.5%), followed by ALB 3 (65.11%); WRA1 has the lowest negative response to inoculation by -26.15%. ALB 1 has the highest relative efficiency of 112.5% followed by GRK 1 which has RE of 111.11% while BKR 1 has the lowest RE of 55.06%.
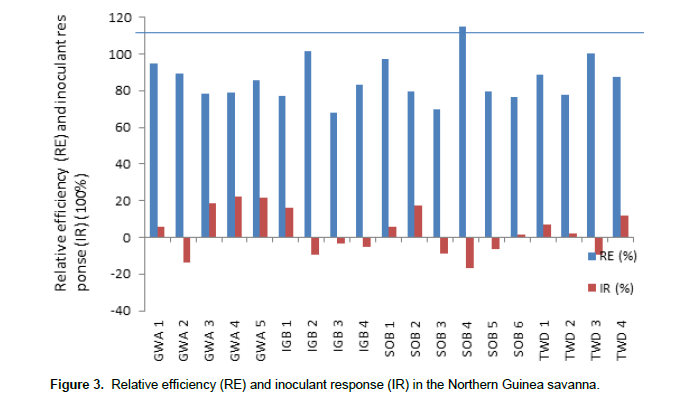
In the Northern Guinea Savanna (Figure 3), the IR ranged from -16.67 to 22.2% while the RE ranged from 67.74 to 114.7%. The highest relative efficiency was observed for SOB 4 with RE of 114.7% followed by IGB 2 with RE of 101.4%; the lowest was IGB 3 (67.74%). The highest response to inoculation was in GWA 4 (22.2%), followed by GWA 5 (21.8%); while the lowest was SOB 4 (-16.67).
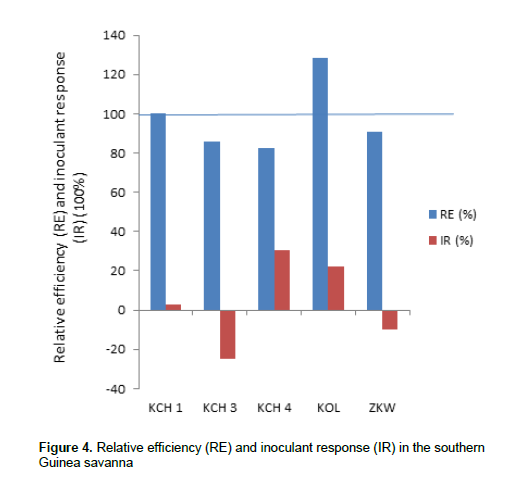
In the Southern Guinea Savanna (Figure 4), the IR ranged from -25 to 30.3% while RE ranged from 82.5 to 128.6%. The highest IR was in KCH 4 (30.3%) followed by KOL (22.22%); while the lowest was in KCH 3 (-25%). The highest RE was observed for KOL (128.6%) followed by KCH 1 (100%); while the lowest RE was in KCH 4 (82.5%). However, samples were collected from relatively few sites, which is not sufficient to provide information that will confidently allow for suitable recommendation. The high relative efficiency observed in some locations could be as a result of the interactions of the population of the native rhizobia in those locations. It is suggested that successful nitrogen fixation depends on the interaction between legume genotype, rhizobium strain and environment (Giller, 2001).
Relative efficiency greater than 100% implies that the indigenous population of rhizobia was superior to the application of mineral N. The strains isolated from these location(s) have very high potential for use as inoculant. Hence, it is not surprising that very few such cases occurred. Notable among such locations are BCH 2 (102.9%), GRK1 (R111.1%), GRK 4 (104.35%), GRK 5 (105.3%), WRA 1 (101.6%) in the Sudan Savanna. In the Northern Guinea Savanna, there is IGB 2 (101.4%), SOB 4 (114.7%).
In the Southern Guinea Savanna, there is KOL (128.6%). Locations with IR greater than 100% show that inoculation with effective rhizobia will improve soybean biomass. The higher inoculant response observed in the Sudan Savanna could be as a result of high competitiveness of the introduced strains. The presence of large population density of compatible rhizobia does not preclude the possibility that responses to inoculation can be obtained if competitive and highly effective strains are introduced (Giller, 2001).
Similarly, the population of indigenous rhizobia could have been very low in the Sudan Savanna. In a study conducted by Aliyu et al. (2013) in Shanono which lies in the Sudan Savanna, it was reported that the native rhizobia population was 1.10 x 102 which was quite low, suggesting that the soils have little or no N fixing ability for soybean. Hence, introduction of a commercial inoculant would have resulted in high response. Kumaga and Etu-Bonde (2000) demonstrated that nodulation and N2 fixation of promiscuous soybean may be increased by inoculation with effective Bradyrhizobia.
The negative response observed in certain locations could be as a result of the competitiveness of the indigenous rhizobia that were more efficient than the introduced strain. It was reported by Meade et al. (1985) that successful competition for nodule sites by native rhizobia is one reason for the failure to achieve a response to inoculation with elite rhizobial strains.
In the Northern Guinea Savanna, SOB 4 interestingly had the highest RE and the lowest IR. This suggests that the population of the indigenous rhizobia may have been highly competitive for nodule occupancy and highly effective in their nitrogen fixing ability. Therefore, there will be no need for inoculation or mineral N application. It is generally argued that indigenous populations are highly adapted to their local soil environments and may form more effective symbioses than commercial inoculants isolated from a distant and un-related soil environment (Gandee et al., 1999).
Similarly, this AEZ has a long history of soybean cultivation and inoculant use (Sanginga et al., 1995, 2003; Okogun et al., 2004; Yusuf et al., 2012). The previously introduced strains might have replaced the indigenous population, thus response to inoculation may be very minimal. This situation can be circumvented if superior strains are present in inoculants to be used in the zone.
CONCLUSION
The results show variation in the response to inoculation of rhizobia with respect to plant biomass in the various agro-ecological zones. The results showed better response to inoculation in the Sudan Savanna compared to the Northern and Southern Guinea savannas. This is not surprising as response to inoculation is usually lower in areas where soybean has been extensively grown. Of all the three agro-ecological zones, Sudan Savanna will benefit more from inoculation. Also, locations with high relative efficiency would produce promising strains if explored further. While the results show response to inoculation and high relative efficiency of indigenous strains, further field studies need to be carried out in order to improve their use as inoculants.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENT
Special thanks to the N2 Africa Project “Putting Nitrogen Fixation to work for small holder farmers in Africa” for providing the grant to conduct this trial.
REFERENCES
|
Anderson JM, Ingram JS (1993). Tropical Soil Biology and Fertility. A Handbook of Methods (2nd ed.), information Press, UK, 221 p. |
|
|
Bokhtiar SM, Sakurai K (2005). Effects of organic manure andchemical fertilizer on soil fertility and productivity of plant and ratoon crops of sugarcane. Arch. Agron. Soil Sci. 51:325-334. |
|
|
Bremner JM, Mulvaney CS (1982). Total nitrogen. In: Page AL, Miller RH, Keeney DR. (eds). Methods of soil analysis. Part 2, Am. Soc. Agron. pp. 595-624. |
|
|
Chapman HD (1965). Cation exchange capacity. In: Black, C. A. (eds), Methods of soil analysis. Part 2, Am. Soc. Agron. 9:702-904. |
|
|
FMANR (Federal Ministry of Agriculture and Natural Resources) (1990). Literature Review on Soil Fertility Investigations in Nigeria (in five volumes). Federal ministry of Agriculture and Natural Resources, Abuja, p281. |
|
|
Gee GW, Bauder JW (1986). Particle size analysis. In : A. Klute (ed) Methods of Soil Analysis 2nd . no 0 Am. Soc. Agron. pp. 383-409. |
|
|
Gandee CM, Harrison SP, Davies WP (1999). Genetic characterization of naturally occurring Rhizobium meliloti populations and their potential to form effective symbiosis with Lucerne. J. Appl. Microbiol. 23:169-174. |
|
|
Giller KE (2001) Nitrogen fixation in tropical cropping systems. 2nd edn. CAB International, Wallingford, UK. 423 p. |
|
|
Jackson ML (1958). Soil chemical analysis. Prentice-Hall Incorporated, Englewood Cliff. New Jersey. 498 p. |
|
|
Jones MJ, Wild A (1975). Soils of the west african savanna.Technical Community No. 55 Commonwealth Bureau of Soils. Harpenden, UK. P 246. |
|
|
Hendershot WH, Laland H, Duquette M (1993). Soil reaction and exchangeable acidity. In: Soil sampling and methods of analysis. Carter, M. R. (ed). Can. Soc. Soil Sci. Lewis publishers London. pp. 141-145. |
|
|
Kolawole GO, Diels J, Manyong VM, Ugbabe O, Wallays K, Iwuafor ENO, Falaki AM, Merckx R, Deckers J, Tollens E, Vanlauwe B, Sanginga N (2007). Balanced Nutrient Management System Technologies in the northern Guinea savanna of Nigeria: Validation and Perspective: In Bationo A, Waswa B, Kihara J, Kimetu J. (Eds), Advances in Integrated Soil Fertility Management in sub-Saharan Africa: Challenges and Opportunities, Springer Verlag, Heidelberg, Germany. pp. 669-678. |
|
|
Kumaga FK, Etu-Bonde K (2000). Response of two promiscuous soybean genotypes to bradyrhizobial inoculation in two Ghanaian soils. J. Ghana Sci. Assoc. 2:99-104. |
|
|
Maingi JM, Gitonga NM, Shisanya C A, Hornetz B, Muluvi GM (2006). Population Levels of Indigenous Bradyrhizobia Nodulating Promiscuous Soybean in two Kenyan Soils of the Semi-arid and Semi-humid Agroecological Zones. J. Agric. Rural Dev. Trop. Subtrop. 107(2):149-159. |
|
|
Manyong VM, Smith GK, Weber SS, Jagtab SS, Oyewole B (1995). Macro characterization of agricultural system in West Africa: An overview. Resource and Crop Management Research Monograph No. 21, International Institute of Tropical Agriculture (IITA), Ibadan, 66 p. |
|
|
Meade J, Higgins P, O'Gara F (1985). Studies on the inoculation and competitiveness of a Rhizobium leguminosarum strain in soils containing indigenous rhizobia. Appl. Environ. Microbiol. 49:899-903. |
|
|
Nelson DW, Sommers LE (1982). Total carbon, organic carbon and organic matter. In: Page AL, Miller RH and Keeney DR (eds). Methods of soil analysis, part 2. American Society of Agronomy. Madison, Wisconsin. pp. 539-579. |
|
|
Ojanuga AG (2006). Agro-ecological Zones of Nigeria Manual. FAO/NSPFS, Federal Ministry of Agriculture and Rural development, Abuja Nigeria. 124 p. |
|
|
Okereke GU, and Eaglesham ARJ (1993). Nodulation and Nitrogen fixation by 79 promiscuous soyabean genotypes in soil in eastern Nigeria. Agron. Afr. 5(2):113-122. |
|
|
Okereke GU, Onochie CC, Onukwo AU, Onyeagba E, Ekejindu GO (2000). Response of introduced Bradyrhizobium strains infecting a promiscuous soybean Cultivar. World J. Microbiol. Biotech. 16:43-48. |
|
|
Okogun JA, Otuyemi BT, Sanginga N (2004). Soybean yield determinants and rhizobial inoculation in an on-farm trial in northern guinea savanna of Nigeria. Agronomy Department University of Ibadan, Nigeria. W. Afr. J. Appl. Ecol. 6:30-39. |
|
|
Olson SR, Sommers LE (1982). Phosphorus. In: Page, A. L., Miller, R. H. and Keeney. D. R. (eds). Methods of soil analysis. Part 2, Am. Soc. Agron. pp. 403-430. |
|
|
Pulver EL, Brockman F, Wein HC (1982). Nodulation of soybean cultivars with Rhizobium spp. and their response to inoculation with R. japonicum. Crop Sci. 22:1070-1085. |
|
|
Sanginga N (2003). Role of biological nitrogen fixation in legume-based cropping systems; a case study of West Africa farming systems. Plant Soil. 252(1):25-39. |
|
|
Sanginga N, Abaidoo R, Dashiell K, Carsky RJ, Okogun A (1995). Persistence and effectiveness of rhizobia nodulating promiscuous soybeans in moist savanna zones of Nigeria. Appl. Soil Ecol. 3:216-224. |
|
|
Sanginga PC, Adesina AA, Manyong VM, Otite O, Dashiell KE (1999). Social impact of soybean in Nigeria's southern Guinea savanna. Int. Institute Trop. Agric. pp. 4-6. |
|
|
Sharu M, Yakubu M, Noma SS, Tsafe, AI (2013). Characterization and Classification of Soils on an Agricultural landscape in Dingyadi District, Sokoto State, Nigeria. Nig. J. Basic Appl. Sci. 21 (2):137-147. |
|
|
Shehu BM, Jibrin JM, Samndi AM (2015). Fertility Status of Selected Soils in the Sudan Savanna Biome of Northern Nigeria. Int. J. Soil Sci. 10:74-83. |
|
|
Sowunmi FA, Akintola JO (2010). Effect of Climate Variability on Maize Production in Nigeria. Res. J. Environ. Earth Sci. 2(1):19-30 |
|
|
Wall JRD (1979). Land resources of the central Nigeria agricultural development possibilities.The Kaduna Plain. Vol. 5. In wall J.R.D. (Ed.), Land Resources Development Center, Ministry of Overseas Development, Tolworth Tower Surbiton, KT6 7DY, Surrey, England, land resource study 29. |
|
|
Wasike VW, Lesueur D, Wachira FN, Mungai NW, Mumera LM, Sanginga N, Mburu HN, Wango P, Vanlauwe B (2009). Genetic diversity of rhizobia nodulating promiscuous soyabean varieties: Impact of phosphorus. Plant Soil 322:151-163. |
|
|
Yusuf AA, Iwuafor ENO, Olufajo OO, Abaidoo RC, Sanginga N (2006). Genotype effects of cowpea and soybean on nodulation, N2-fixation and N balance in the northern Guinea savanna of Nigeria. Proceedings of the 31st Annual Conference of the Soil Science Society of Nigeria (SSSN) held between 13th and 17th at ABU, Zaria, Nigeria, pp. 147-154. |
|
|
Yusuf AA, Jemo M, Nwoke OC, Abaidoo RC (2012). Evaluation of commercial and laboratory rhizobium inoculants on nodulation and yield of promiscuous soybean in the Nigerian savanna. In: M.K.A. Adeboye, A.J. Odofin, A.O. Osunde, A. Bala and S.O. Ojeniyi (eds). Soil resources management, gobal climate change and food security. Proceedings of the 35th Annual Conference of the Soil Science of Nigeria. pp. 199-214. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


