ABSTRACT
This study evaluated the biotechnological potential of the endophytic fungus Diaporthe species, which is isolated from Brazilian mangroves. In vitro studies have investigated the antimicrobial activity of Diaporthe spp. against human pathogens, such as Escherichia coli, Staphylococcus aureus, Salmonella enteritidis, and Candida albicans, as well as phytopathogenic fungi, such as Colletotrichum species, Ceratocystis paradoxa, Fusarium oxysporum, Phythophthora sojae and Rhizopus microspores. Additionally, the enzyme production and leishmanicidal and cytotoxic activities of Diaporthe spp. were tested. Minimal inhibitory concentrations (MICs) of the crude extract of Diaporthe spp. 94 ranged from 756 to 949 µg.mL-1 (MIC50) and from 3.094 to 7.082 μg.mL-1 (MIC90); the antagonism index (AI) ranged from 20 to 62% with type A interactions (“deadlock” with mycelial contact). An assay to detect enzyme production showed that this endophytic fungus produced the enzyme cellulase. The crude extract of Diaporthe spp. also demonstrated activity against the promastigote form of Leishmania infantum chagasi, exhibiting 90% cell death at 6.000 and 10.000 µg.mL-1 and 80% cell death at 4.000 µg.mL-1. Cytotoxicity tests on cultured human skin fibroblasts (the HFF-1 cell line) indicated cell viability of between 73 and 55% at 4.000 to 10.000 μg.mL-1 at 24 h; between 69 and 56% at 4.000 to 8.000 μg.mL-1 for 48 h, and 59% at 4.000 to 6.000 μg.mL-1 for 72 h.
Key words: Diaporthe species, endophytic fungus, leishmanicidal activities, cytotoxic activities, antimicrobial activities, enzymatic activities.
Mangroves are ecosystems located in transition areas between terrestrial and marine environments, which has forced them to adapt to such conditions as high salinity, tidal flooding, high temperatures, anaerobic soil, and large amounts of sludge. Thus, mangroves represent a useful ecosystem for identifying plants and microorganisms that have ecological, morphological, biological, and physiological adaptations and survive under these particular conditions (Jalgaonwala et al., 2011; Sebastianes et al., 2013). Endophytes are fungi or bacteria that colonize plants from the inside in symbiotic relationships, causing no visible harm to the host (Strobel, 2018). However, depending on external factors, such as environmental conditions and the host, endophytes may switch between pathogenic and commensal or mutualistic lifestyles (Bacon et al., 2008; Fesel and Zuccaro, 2016). Since the identification of the first endophytic fungus, significant attention has been devoted to the identification of new bioactive compounds that can be synthesized by them (Sebastianes et al., 2012; Sebastianes et al., 2013; Strobel, 2018). This research provides an alternative to plant exploitation, thereby facilitating the conservation of flora diversity, which is increasingly under threat worldwide (Zheng et al., 2015; Gurgel et al., 2020), as well as reducing the market value of these biomolecules (Strobel and Daisy, 2003; Sebastianes et al., 2012; Bibi et al., 2020). The broad spectrum of biological activity exhibited by endophytic fungal isolates enables, among other things, the exploration of substances with potential for antimicrobial and enzyme production, which can be achieved on a large scale through fermentation and presents considerable potential for industrial applications (Dezam et al., 2017; Marques et al., 2018; Yan et al., 2018; Gurgel et al., 2020). Several studies have described endophytic fungi with antimicrobial activities (Ratnaweera et al., 2015; Bezerra et al., 2015; Campos et al., 2015; Chen et al., 2016, 2019; Khan et al., 2016; Deshmukh et al., 2020) and enzymatic activities in several plant species and mangroves from different regions (Bezerra et al., 2015; Martinho et al., 2019).
Secondary metabolites produced by endophytic fungi also have promising applications in phytopathogen control (Macías-Rubalcava and Sánchez-Fernández, 2017), as demonstrated by studies employing antagonism tests.
Endophytes may present activity against human parasites that are commonly related to neglected diseases that infect millions of people worldwide, especially in developing countries. Among these parasites, Leishmania infantum chagasi, the aetiological agent of visceral leishmaniasis, causes high morbidity and mortality (Maran et al., 2016).
The most common form of leishmaniasis is the cutaneous form. Cutaneous leishmaniasis is caused by several species of Leishmania, including Leishmania braziliensis and Leishmania amazonensis. However, this leishmaniasis is the least severe and lethal. A number of studies have demonstrated activities against this type of leishmaniasis (Santiago et al., 2011; Cota et al., 2018). To date, there have been no studies investigating the role played by related endophytic fungi from mangroves in producing secondary metabolites with leishmanicidal activity against L. infantum chagasi.
Fungi from marine environments grow under particular conditions that can contribute to the synthesis of new compounds (Sebastianes et al., 2012). In this context, endophytic fungi associated with plants that inhabit mangrove ecosystems may represent a promising source of secondary metabolites that have not been fully elucidated to date. The present study evaluated the in vitro antimicrobial, antagonistic and leishmanicidal activities, enzyme production, and cytotoxicity activities of secondary metabolites produced by the endophytic fungus Diaporthe species 94 strain isolated from Brazilian mangroves.
Microorganisms
The endophytic fungus Diaporthe spp. FS-94(4) strain (https://www.ncbi.nlm.nih.gov/nuccore/HQ022906.1) was isolated from Avicennia nitida branches in mangrove areas of São Paulo State, Brazil (Sebastianes et al., 2013). The phytopathogens Colletotrichum species, Ceratocystis paradoxa, Fusarium oxysporum, Phythophthora sojae, and Rhizopus microsporus were selected for antagonistic assays. The human pathogen strains Escherichia coli (ATCC 25922), Salmonella enteritidis (ATCC 19196), Staphylococcus aureus (ATCC 6538) and Candida albicans (ATCC 10231) were selected from the American Type Culture Collection (ATCC) for antimicrobial assays. All microorganisms were deposited in the Microbiology and Biomolecules Laboratory (LaMiB), Microorganism Collection, Morphology and Pathology Department, Federal University of São Carlos, Brazil.
L. infantum chagasi (MHOM/BR/1972/LD), deposited in the collection of Inflammation and Infectious Diseases Laboratory (LIDI), Federal University of São Paulo, Brazil, and human skin fibroblasts from a cell line (HFF-1) deposited in the collection of the National Institute of Physics of Optics and Photonics Science and Technology, University of São Paulo, Brazil, were tested via cytotoxicity assays.
Culture conditions and obtaining crude extracts
To obtain crude extracts (CE), the Diaporthe spp. 94 strain was grown in Petri dishes containing medium potato dextrose agar (PDA) (KASVITM, Brazil) for 7 days at 28°C. Eight discs (5 mm diameter) of mycelia from these cultures were taken from the plates and inoculated into 500-mL Erlenmeyer flasks containing 200 mL of potato dextrose liquid medium (PD) (KASVITM, Brazil) followed by incubation for 7 days at 28°C in the absence of light and static conditions. The culture medium containing metabolites was separated from the mycelium by a 1-µm paper filter and subsequently utilized in biological assays.
Antimicrobial activity assays
Broth microdilution techniques were employed in 96-well microplates. The inoculum was standardized at 0.5 on the scale of McFarland (1.5 × 108) (McFarland, 1907) and carried out a dilution of 1:10 in Mueller Hinton (MH) broth (KASVITM, Brazil), obtaining a bacterial suspension of 1.0 × 107 colony forming units (CFU/mL). Serial dilutions of the CE were prepared in MH broth at concentrations of 10.000 to 1.000 μg.mL-1. Penicillin and streptomycin (GIBCO, USA) were employed as antimicrobial agent standards at a concentration of 2.500 µg.mL-1 was added to 100 µL of bacterial cells adjusted to the concentration 0.5 on the McFarland scale and 100 µL of the crude extract of each concentration, obtaining a final volume of 200 µl per well. The microplates were incubated at 37°C for 24 h. After incubation, 20 µl of Alamar Blue® (Sigma-Aldrich, USA) was added to each well for a final concentration of 0.01%. Subsequently, the microplates were read at 570 nm in a fluorescence spectrophotometer (SPECTRAMAX M3).
Antagonism assays
A mycelial disc (5 mm in diameter) taken from a 7-day culture of the Diaporthe 94 (4) strain on PDA was placed at one edge of a 15-cm Petri dish containing PDA (KASVITM, Brazil). On the same day, an inoculum of each phytopathogen to be tested (5-mm-diameter mycelial disc) was added to another edge of the plate at a distance of 2 cm from the margin. The plates were incubated at 28°C and observed for 10 consecutive days. Plates inoculated with phytopathogens alone were employed as controls. The tests were performed in triplicate, and the analysis of endophytic-phytopathogen interactions was based on the scale of Badalyan et al. (2004) according to 3 types (A, B and C) and 4 subtypes (CA1, CA2, CB1, and CB2), where A means deadlock with mycelial contact, B means deadlock at a distance, C means replacement and overgrowth without initial deadlock, CA1 means partial and complete replacement after initial deadlock with mycelial contact, CA2 means complete replacement after initial deadlock, and CB1 and CB2 mean partial and complete replacement after initial deadlock at a distance, respectively. The antagonism index (AI) was calculated after 10 days of incubation using the formula of Campanile et al. (2007): AI = (RM – rm) / RM × 100, where rm is the ray of the colony towards the antagonist, and RM is the average of the three rays of the colony in the other directions.
Enzymatic activity
The Diaporthe 94 (4) strain was evaluated for enzymatic activity using a qualitative method. The following enzymes were analysed: amylase (Buzzini and Martini, 2002), cellulase (Teather and Wood, 1982), protease (Castro et al., 2014), lipase and esterase (Sierra, 1957). For these tests, the endophytic strain was previously grown on a 15-cm Petri dish containing PDA at 28°C for 8 to 12 days, and discs of mycelium (5 mm in diameter) were inoculated onto the centre of plates containing specific medium for each enzyme. The plates were incubated at 28°C, and the enzymatic activity was observed based on the presence or absence of halos surrounding mycelial discs. The tests were performed in triplicate.
Leishmanicidal activity
L. infantum chagasi in its promastigote form was cultured in 25-cm2 T25 flasks (KASVI, BRA) containing medium 199 (SIGMA-ALDRICH, USA) supplemented with foetal bovine serum (FBS) (LGC, BRA), pen/strep antibiotics (GIBCO, USA) and L-glutamine (SIGMA-ALDRICH, USA). In the assay, each well contained 100 µL of a cell suspension previously adjusted to 1 × 105 and 100 µL of the CE diluted into four different concentrations: 10.000, 8.000, 6.000 and 4.000 µg.mL-1, with 200 µL being the final volume of each well. Amphotericin B (GIBCO, USA) was used as a leishmanicidal agent standard at a concentration of 0.1 mM. The plates were incubated at 23°C for 24 and 48 h. For the cell viability test, the colorimetric Alamar Blue® (Sigma-Aldrich, USA) was employed. After each incubation period, 1.5 mM Alamar Blue® was added and incubated in the incubator, with CO2 levels being maintained between 5 and 10%. Absorbance was read in a microplate reader (TP-READER, NM-THERMO PLATE) with a wavelength of 550 nm.
Cytotoxicity assay
For cytotoxicity tests, HFF1 cells were cultured in 75-cm2 flasks (GREINER, USA) containing Dulbecco’s Modified Eagle’s Medium (DMEM) (SIGMA-ALDRICH, USA) supplemented with foetal bovine serum (FBS) (HYCLONE, USA), penicillin/streptomycin antibiotics (GIBCO, USA), L-glutamine, HEPES and bicarbonate (SIGMA-ALDRICH, USA). In 96-well plates (KASVI, BRA), 100 µL of medium was added containing cells at a concentration of 1×105, and 100 µL of EC at concentrations of 20.000, 10.000, 8.000, 6.000 and 4.000 µg.mL-1, was added next, obtaining a final volume of 200 µL per well. After inoculation, cultures were kept in an incubator at 37°C and 5% CO2 for 24, 48, and 72 h. For the assessment of cell viability, a colorimetric method was employed, with 3-(4,5-dimethyl thiazole 2-yl) 2,5-diphenyl bromide tetrazolium (MTT) (Sigma-Aldrich, USA) serving as the standard.
The absorbance was read in a microplate reader (Thermo Scientific®, USA) at a 570-nm wavelength.
Statistical analysis
Data were submitted to the Shapiro-Wilk normality test and Levene’s test of homogeneity. Analysis of variance (ANOVA) and Tukey analysis were used to indicate the differences between the experimental groups. The values were considered to be significant when P-values ≤ 0.05 were obtained using GraphPad Prism software (Version 7.04 for Windows). Origin Pro version 9 software was employed to produce graphics and calculate the MIC50 and MIC90 (slope of linear regression) values.
Antimicrobial activity assays
The MIC50 and MIC90 were the minimum concentrations of CE required to inhibit 50 and 90%, respectively, of the pathogens tested. The data showed that CE was effective against the following pathogens: E. coli, MIC50 = 767 µg.mL-1 and MIC90 = 7.820 μg.mL-1; S. enteritidis, MIC50 = 949 µg.mL-1 and MIC90 = 3.980 µg.mL-1; and S. aureus, MIC50 = 756 μg.mL-1 and MIC90 = 3.940 µg.mL-1 (Table 1).
Antagonism assays
Diaporthe spp. 94 (4) strain inhibited the mycelial growth of Colletotrichum spp. by 33%, and the endophyte-phytopathogen interaction observed was of type CA1. For F. oxysporum, the mycelial inhibition was 50%, and the endophyte-phytopathogen interaction observed was of type CA1. For P. sojae, the mycelial inhibition was 20%, and the endophyte-phytopathogen interaction observed was of type A. Finally, for R. microsporus, the mycelia inhibition was 62%, and the endophyte-phytopathogen interaction observed was of type A. There was no activity against C. paradoxa. Type A interaction occurs when there is a deadlock with mycelial contact, and type CA1 interaction occurs when there is a partial replacement after the initial deadlock with mycelial contact (Table 2).
Enzymatic activity
Qualitative enzymatic assays showed that the endophytic Diaporthe spp. 94 (4) strain produced the enzyme cellulase. The assays did not indicate production of the enzymes amylase, protease, lipase or esterase.
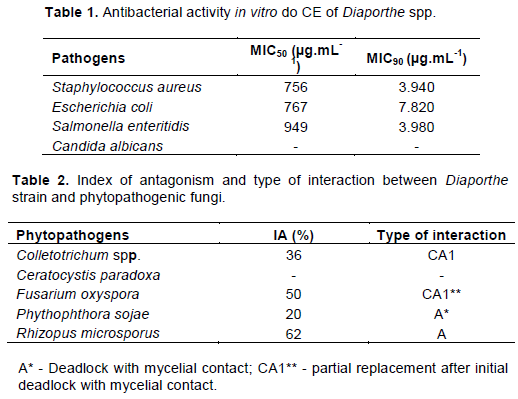
Leishmanicidal assays
The CE produced by Diaporthe strain 94 (4) was tested against the promastigote forms of L. infantum chagasi in the periods of 24 and 48 h at 4.000, 6.000, 8.000 and 10.000 µg.mL-1. In the 24-h period, the concentration of 4.000 µg.mL-1 had a mortality greater than 80%, and in the concentrations of 6.000, 8.000 and 10.000 µg.mL-1, there was mortality of at least 95% of the cells. In the 48-h period, the concentration of 4.000 µg.mL-1 had a mortality greater than 75%, and in the concentrations of 6.000, 8.000 and 10.000 µg.mL-1, there was mortality of at least 95% of the cells. (Figure 1). In the 24 and 48-h periods, the concentration of 4.000 µg.mL-1 was significantly different from the concentrations of 6.000, 8.000 and 10.000 µg.mL-1; the concentrations of 6.000 and 8.000 µg.mL-1 were significantly different from the concentration of 10.000 µg.mL-1.
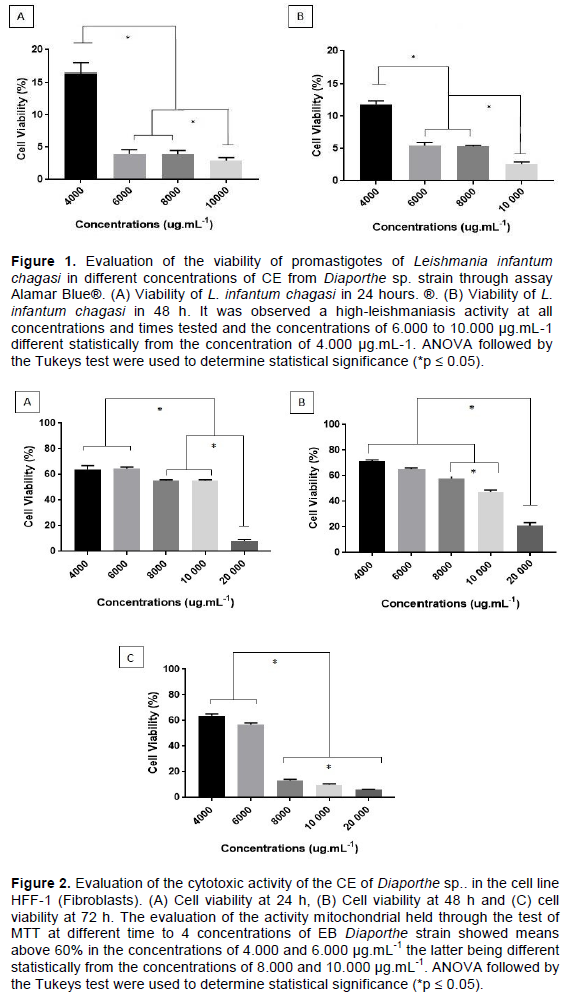
Cytotoxicity
Assays for cytotoxicity showed cell viability above 60% at concentrations of 4.000 and 6.000 µg.mL-1, above 50% at concentrations of 8.000 and 10.000 µg.mL-1, and less than 10% at a concentration of 20.000 µg.mL-1 in the 24-h period. In the 48-h period, cell viability was above 60% at concentrations of 4.000 and 6.000 µg.mL-1, above 50% at a concentration of 8.000 µg.mL-1, above 40% at a concentration of 10.000 µg.mL-1, and above 15% at a concentration of 20.000 µg.mL-1. In the 72-h period, the cell viability was above 60% at a concentration of 4.000 µg.mL-1, above 50% at a concentration of 6.000 µg.mL-1 and up to 20% at concentrations of 8.000, 10.000 and 20.000 µg.mL-1 (Figure 2). In the 24-h period, the concentrations of 4.000 and 6.000 µg.mL-1 were significantly different from the concentrations of 8.000, 10.000 and 20.000, and the concentration of 8.000 µg.mL-1 was significantly different from the concentration of 10.000 µg.mL-1. In the 48-h period, the concentrations of 4.000, 6.000, 8.000 and 10.000 µg.mL-1 were significantly different from the concentration of 20.000, and the concentration of 8.000 µg.mL-1 was significantly different from the concentration of 10.000 µg.mL-1. In the 72-h period, the concentrations of 4.000 and 6.000 µg.mL-1 were significantly different from the concentrations of 8.000, 10.000 and 20.000, and the concentrations of 8.000, 10.000 and 20.000 µg.mL-1 were significantly different from each other.
Endophytic fungi isolated from mangrove ecosystems, including fungi belonging to the genus Diaporthe, have been described as rich sources of biologically active compounds with a variety of applications (Lin et al., 2005; Huang et al., 2008; Sebastianes et al., 2012; Brissow et al., 2017; de Medeiros et al., 2018; Huang et al., 2019), opening new avenues for biotechnological research. In this study, we evaluated the biotechnological potential of crude extract from Diaporthe spp. 94 isolates that were isolated from Brazilian mangroves for applications in the fields of medicine, agriculture and industry. Thus, evaluating the various effects of these compounds may enable researchers to identify an antimicrobial or antiparasitic effect to control different diseases.
The emergence of multidrug-resistant microorganisms has generated considerable attention and stimulated the search for new drugs. In this context, studies on the antimicrobial activity of endophytic mangrove-derived fungi against resistant human pathogens have been demonstrated (Prihanto et al., 2011; Sebastianes et al., 2012; Handayani et al., 2017; de Medeiros et al., 2018; Huang et al., 2019).
Bezerra et al. (2015) tested 32 endophytic fungi isolated from Bauhinia forficata against 10 pathogenic bacteria in disc diffusion assays. Of these 32 isolates, 11 presented antibacterial activity against one or more bacteria among S. aureus (UFPEDA02), Streptococcus pyogenes (UFPEDA07), Mycobacterium smegmatis (UFPEDA71), Bacillus subtilis (UFPEDA86), Enterococcus faecalis (UFPEDA138), Salmonella Typhi (UFPEDA478), Pseudomonas aeruginosa (UFPEDA735), Enterobacter aerogenes (UFPEDA739), Proteus vulgaris (UFPEDA740), and E. coli (UFPEDA224). Among the bacteria tested in common with this present study are E. coli, where six endophytic fungi presented antimicrobial action, S. aureus and the genus Salmonella, where for each, an endophytic fungus showed activity. Therefore, our results show promise, since the dose exhibiting effectiveness against pathogenic bacteria is ten times lower than the dose showing cytotoxicity in mammalian cells. Thus, our data suggest that this compound could be evaluated in a future study seeking new biomolecules with promising activity for future antibiotics.
Campos et al. (2015) isolated 82 endophytic fungi from the Caesalpinia echinata plant and tested their ethyl acetate extracts. Three extracts presented activity against E. coli and S. aureus, where MIC values varied between 32 and 64 µg/mL, and one presented activity against Salmonella bacteria, where MIC was 64 µg/mL. The MIC values were lower than those in the present study, probably due to the purification of the extract.
However, the fact that our compound has a ten- or twelve-fold greater effect on fibroblast cytotoxicity makes it a promising biomolecule for future studies attempting to achieve microbial control.
Ratnaweera et al. (2015) isolated eight endophytic fungi of Opuntia dillenii. The eight strains showed antibacterial activity against at least one of the tested bacteria, including E. coli and S. aureus. Additionally, Khan et al. (2016) isolated the endophytic fungus Cladosporium species of Rauwolfia serpentina, and its extract exhibited antibacterial activity against E. coli and S. aureus among other bacteria. The endophytic Diaporthe presented promising antimicrobial activity against S. aureus, E. coli and S. enteritidis. When comparing our results with those of other authors, we must evaluate the antimicrobial effects and the dose-dependent effect in killing the studied bacteria. Our data suggest that this activity is dose-dependent and may favour the regulation of the dose used, given the evaluated antimicrobial effect. We suggest that in future tests, after producing these compounds in greater purity, we can amplify this effect and achieve greater bacterial control. Endophytic fungi provide novel opportunities to control phytopathogens (Campanile et al., 2007). The antagonistic activity displayed by Diaporthe spp. 94 in our study could be explained by their secreting diffusible antifungal compounds into the culture medium, thereby inhibiting mycelial growth of Colletotrichum spp., F. oxysporum, P. sojae and R. microsporus. This inhibition could also be due to competing for space and/or nutrient depletion in the medium in the dual-culture assay. Competitive interactions of type A (deadlock with mycelial contact) and type CA1 (partial replacement after initial deadlock with mycelial contact), which were observed in the antagonistic assays, indicate different modes of action of Diaporthe spp. on phytopathogens, supporting our hypotheses regarding the possible mechanisms of control employed by endophytes.
Lytic enzymes secreted by endophytes, including cellulases, are another important mechanism by which plant-associated fungi control pathogens (Tripathi et al., 2008). These enzymes can suppress phytopathogens from breaking down cell walls, thereby restricting their growth (Gao et al., 2010). In this study, Diaporthe spp. 94 secreted cellulase into the culture medium. This potential for extracellular enzyme production by endophytic fungi from mangroves has also been described by other studies (Maria et al., 2005; Thatoi et al., 2013; Bezerra et al., 2015; Maroldi et al., 2018). These microorganisms naturally degrade lignocellulosic plant biomass of sediments in mangrove environments (Behera et al., 2017) and thus also suggest other applications, such as the processing of fruit juices in the textile, food and paper industries (Gao et al., 2008).
Leishmaniasis is a neglected tropical disease, and its prevalence in the poorest populations is not a major stimulus for the pharmaceutical industry. Thus, the search for new medicines to treat leishmaniasis has not been heavily funded. Therefore, contributing studies that may lead to the discovery of a compound or compounds with anti-leishmaniasis activity is of great importance for the control of these diseases. Similar works tested endophytic fungi isolated from plants from different Brazilian biomes, and they obtained leishmanicides that exhibited promise against other species of Leishmania that cause integumentary leishmaniasis, which is less deadly than visceral leishmaniasis (VL) (Marinho et al., 2005; Campos et al., 2008; Rosa et al., 2010; Brissow et al., 2017; Cota et al., 2018). Little is known about the anti-leishmaniasis activity of these isolates against the species that cause VL (Leishmania donovani and L. infantum chagasi). The results obtained for the control of L. chagasi demonstrate that the compound has leishmanicidal activity at a dose similar to the IC50 observed in fibroblasts. However, most of the drugs available and recommended to treat visceral leishmaniasis are highly toxic. Thus, in future research, we will evaluate in vivo whether the toxic effect is capable of compromising homeostasis in a murine model. Thus, we will be able to evaluate these data more globally. However, these results are promising for future studies seeking a new compound for the control of visceral leishmaniasis.
In Brazil, the aetiological agent of VL is L. infantum chagasi, our choice for the testing of inhibition. Our data demonstrated a high mortality rate at a dose of 4.000 µg.mL-1 in different periods of exposure (24 and 48 h) (Figure 1). These data are promising compared with those found in the literature, where tests for less aggressive leishmaniasis were performed (Marinho et al., 2005; Campos et al., 2008; Santiago et al., 2011; Campos et al., 2015; Brissow et al., 2017).
The search for compounds with biological activity has great importance for research correlating new therapies for existing diseases. However, to continue the studies, it is of great relevance for these studies to demonstrate that for basal cells, the toxicity is considerably lower compared to the toxicity observed on the pathogenic microorganisms (Figure 2). Thus, we evaluated fibroblast cytotoxicity, an important model to establish these parameters. Thus, our study leads us to new bioprospects from these compounds, since the cytotoxicity in fibroblasts was considerably lower than that found in the pathogens studied. Thus, we suggest that these biomolecules are promising for future studies in which we are searching for new compounds with biological activity.
This study demonstrated the biotechnological potential of the endophytic fungus Diaporthe spp. 94 isolated from Brazilian mangroves. We described for the first time the leishmanicidal activity of the secondary metabolites of this fungus on the promastigote forms of L. infantum chagasi and the cytotoxic effects of metabolites on HFF-1 cells. Our results may provide a new biological source of novel drugs.
The results of this study indicate that the tested CE from Diaporthe spp. 94 has antimicrobial action and anti-leishmaniasis activity at the two lower concentrations and has low toxicity at those same concentrations. These results suggest a that is fungus has strong biotechnological potential for future applications, as it is important to identify new drugs and antimicrobials because the rapid resistance that microorganisms have acquired is not compatible with new drug production. It is also important to develop new leishmanicidal drugs with lower cost and toxicity and greater effectiveness.
The authors declare that there are no conflicts of interest.
This project was supported by grants from Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP 2014/07331-5 and 2016/13423-5) and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - Brasil (CAPES) (Proc. n° 88882.426505/2019-01; Finance Code 001).
REFERENCES
|
Bacon CW, Glenn AE, Yates IE (2008). Fusarium verticillioides: managing the endophytic association with maize for reduced fumonisins accumulation. Toxin Reviews 27:411-446.
Crossref
|
|
|
|
Badalyan SM, Inocennti G, Garibyan G (2004). Interaction between xylotrophic mushrooms and mycoparasitic fungi in dual culture experiment. Phytopathologia Mediterranea 43(1):44-48.
|
|
|
|
|
Behera BC, Sethi BK, Mishra RR, Dutta SK, Thatoi HN (2017). Microbial cellulases - Diversity & biotechnology with reference to mangrove environment: A review. Journal of Genetic Engineering and Biotechnology 15(1):197-210.
Crossref
|
|
|
|
|
Bezerra JD, Nascimento CC, Barbosa R do N, da Silva DC, Svedese VM, Silva-Noueira EB, Gomes BS, Paiva LM, Souza-Motta CM (2015). Endophytic fungi from medicinal plant Bauhinia forficata: Diversity and biotechnological potential. Brazilian Journal of Microbiology 46(1):49-57.
Crossref
|
|
|
|
|
Bibi SN, Gokhan Z, Rajesh J, Mahomoodally MF (2020). Fungal endophytes associated with mangroves Chemistry and biopharmaceutical potential. South African Journal of Botany.
Crossref
|
|
|
|
|
Brissow ER, da Silva IP, de Siqueira KA, Senabio JA, Pimenta LP, Januário AH, Magalhães LG, Furtado RA, Tavares DC, Junior PAS, Santos JL, Soares MA (2017). 18-Des-hydroxy Cytochalasin: an antiparasitic compound of Diaporthe phaseolorum-92C, an endophytic fungus isolated from Combretum lanceolatum Pohl ex Eichler. Parasitology Research 116(7):1823-1830.
Crossref
|
|
|
|
|
Buzzini P, Martini A (2002). Extracellular enzymatic activity profiles in yeast and yeast-like strains isolated from tropical environments. Journal of Applied Microbiology 93(6):1020-1025.
Crossref
|
|
|
|
|
Campanile G, Ruscelli A, Luisi N (2007). Antagonistic activity of endophytic fungi towards Diplodia corticola assessed by in vitro and in planta tests. European Journal of Plant Pathology 17(3):237-246.
Crossref
|
|
|
|
|
Campos FF, Junior, PAS, Romanha AJ, Araújo MSS, Siqueira EP, Resende JM, Alves TMA, Martins-Filho OA, dos Santos VL, Rosa CA, Zani CL, Cota BB (2015). Bioactive endophytic fungi isolated from Caesalpinia echinata Lam. (Brazilwood) and identification of beauvericin as a trypanocidal metabolite from Fusarium sp.. Memórias Do Instituto Oswaldo Cruz 110(1):65-74.
Crossref
|
|
|
|
|
Campos FF, Rosa LA, Cota BB, Caligiorne RB, Rabello ALT, Alvez TMA, Rosa CA, Zani CL (2008). Leishmanicidal metabolites from Cochliobolus sp., an endophytic fungus isolated from Piptadenia adiantoides (Fabaceae). Plos Neglected Tropical Diseases 3(1).
Crossref
|
|
|
|
|
Castro RA, Quecine MC, Lacava PT, Batista BD, Luvizotto DM, Marcon J, Ferreira A, Melo IS, Azevedo JL (2014). Isolation and enzyme bioprospection of endophytic bacteria associated with plants of brazilian mangrove ecosystem. Springer PLUS 3:382.
Crossref
|
|
|
|
|
Chen S, Chen D, Cai R, Cui H, Long Y, Lu Y, Li C, She Z (2016). Cytotoxic and antibacterial preussomerins from the mangrove endophytic fungus Lasiodiplodia theobromae ZJ-HQ1. Journal of Natural Products 79(9):2397-2402.
Crossref
|
|
|
|
|
Chen Y, Yang W, Zou G, Chen S, Pang J, She Z (2019). Bioactive polyketides from the mangrove endophytic fungi Phoma sp. SYSU-SK-7. Fitoterapia 139:104369.
Crossref
|
|
|
|
|
Cota BB, Tunes LG, Maia DNB, Ramos JP, de Oliveira DM, Kohlhoff M, Alves TMA, Souza-Fagundes EM, Campos FF, Zani CL (2018). Leishmanicidal compounds of Nectria pseudotrichia, an endophytic fungus isolated from the plant Caesalpinia echinata (Brazilwood). Memórias Do Instituto Oswaldo Cruz 113(2):102-110.
Crossref
|
|
|
|
|
de Medeiros AG, Savi DC, Mitra P, Shaaban KA, Jha AK, Thorson JS, Rohr J, Glienke C (2018). Bioprospecting of Diaporthe terebinthifolii LGMF907 for antimicrobial compounds. Folia Microbiologica 63(4):499-505.
Crossref
|
|
|
|
|
Deshmukh SK, Agrawal S, Prakash V, Gupta MK, Reddy MS (2020). Anti-infectives from mangrove endophytic fungi. South African Journal of Botany.
Crossref
|
|
|
|
|
Dezam APG, Vasconcellos VM, Lacava PT, Farinas CS (2017). Microbial production of organic acids by endophytic fungi. Biocatalysis and Agricultural Biotechnology 11:282-287.
Crossref
|
|
|
|
|
Fesel PH, Zuccaro A (2016). Dissecting endophytic lifestyle along the parasitism/mutualismo continuum in Arabidopsis. Current Opinion in Microbiology 32:103-112.
Crossref
|
|
|
|
|
Gao FK, Dai CC, Liu XZ (2010). Mechanism of fungal endophytes in plant protection against pathogen. African Journal of Microbiology Research 4(13):1346-1351.
|
|
|
|
|
Gao J, Weng H, Zhu D, Yuan M, Guan F, XI Y (2008). Production and characterization of cellulolytic enzymes from the thermoacidophilic fungal Aspergillus terreus M11 under solid-state cultivation of corn stover. Bioresource Technology 99(6):7623-7629.
Crossref
|
|
|
|
|
Gurgel RS, Rodrigues JGC, Matias RR, Batista BN, Oliveira RL, Albuquerque PM (2020). Biological activity and production of metabolites from Amazon endophytic fungi. African Journal of Microbiology Research 14(2):85-93.
Crossref
|
|
|
|
|
Handayani D, Rivai H, Hutabarat M, Rasyid R (2017). Antibacterial activity of endophytic fungi isolated from mangrove plant Sonneratia griffithii Kurz. Journal of Applied Pharmaceutical Science 7(4):209-212.
|
|
|
|
|
Huang Z, Cai X, Shao C, She Z, Xia X, Chen Y, Yang J, Zhou S, Lin Y (2008). Chemistry and weak antimicrobial activities of Phomopsis produced by mangrove endophytic fungus Phomopsis sp. ZSU-H76. Phytochemistry 69(7):1604-1608.
Crossref
|
|
|
|
|
Huang X, Zhou D, Liang Y, Liu X, Cao F, Qin Y, Mo T, Xu Z, Li J, Yang R (2019). Cytochalasins from endophytic Diaporthe sp. GDG-118. Natural Product Research 13:1-8.
Crossref
|
|
|
|
|
Jalgaonwala RE, Mohite BV, Mahajan RT (2011). A review: Natural products from plant associated endophytic fungi. Journal of Microbiology and Biotechnology 1(2):21-32.
|
|
|
|
|
Khan IH, Sohrab MH, Rony SR, Tareq FS, Hasan CM, Mazid A (2016). Cytotoxic and antibacterial naphthoquinones from an endophytic fungus, Cladosporium sp. Toxicology Reports 3:861-865.
Crossref
|
|
|
|
|
Lin X, Huang Y, Fang M, Wang J, Zheng Z, Su W (2005). Cytotoxic and antimicrobial metabolites from marine lignicolous fungi, Diaporthe sp..FEMS Microbiology Letters 251(1):53-58.
Crossref
|
|
|
|
|
Macías-Rubalcava ML, Sánchez-Fernández RE (2017). Secondary metabolites of endophytic Xylaria species with potential applications in medicine and agriculture. World Journal of Microbiology and Biotechnology 33(15).
Crossref
|
|
|
|
|
Maran N, Gomes PS, Freire-de-Lima L, Freitas EO, Freire-de-Lima CG, Morrot A (2016). Host resistance to visceral leishmaniasis: Prevalence and prevention. Expert Review of Anti-infective Therapy 14(4):435-442.
Crossref
|
|
|
|
|
Maria GL, Sridhar KR, Raviraja NS (2005). Antimicrobial and enzyme activity of mangrove endophytic fungi of southwest coast of India. Journal of Agricultural Technology 1:67-80.
|
|
|
|
|
Marinho AMR, Rodrigues-Filho E, Moitinho MLRM, Santos LS (2005). Biologically active polyketides produced by Penicillium janthinellum isolated as an endophytic fungus from fruits of Melia azedarach. Journal of the Brazilian Chemical Society 16(2):280-283.
Crossref
|
|
|
|
|
Maroldi MMC, Vasconcellos VM, Lacava PT, Farinas CS (2018). Potential of mangrove-associated endophytic fungi for production of carbohydrolases with high saccharification efficiency. Applied Biochemistry and Biotechnology 184(3):806-820.
Crossref
|
|
|
|
|
Marques NP, de Cassia Pereira J, Gomes E, da Silva R, Araújo AR, Fereira H, Rodrigues A, Dussán KJ, Bocchini DA (2018). Cellulases and xylanases production by endophytic fungi by solid state fermentation using lignocellulosic substrates and enzymatic saccharification of pretreated sugarcane bagasse. Industrial Crops and Products 122:66-75.
Crossref
|
|
|
|
|
Martinho V, Lima LMS, Barros CA, Ferrari VB, Passarini MRZ, Santos LA, Sebastianes FLS, Lacava PT, Vasconcellos SP (2019). Enzymatic potential and biosurfactant production by endophytic fungi from mangrove forest in Southeastern Brazil. AMB Express 9:130.
Crossref
|
|
|
|
|
McFarland J (1907). Nephelometer: an instrument for estimating the number of bacteria in suspensions used for calculating the opsonic index and for vaccines. Journal of the American Medical Association 14:1176-1178.
Crossref
|
|
|
|
|
Prihanto AA, Firdaus M, Nurdiani R (2011). Endophytic fungi isolated from mangrove (Rhyzopora mucronata) and its antibacterial activity on Staphylococcus aureus and Escherichia coli. Journal of Food Science and Engineering 1:386-389.
|
|
|
|
|
Ratnaweera PB, de Silva ED, Williams DE, Andersen RJ (2015). Antimicrobial activities of endophytic fungi obtained from the arid zone invasive plant Opuntia dillenii and the isolation of equisetin, from endophytic Fusarium sp.. BMC Complementary and Alternative Medicine 15:220.
Crossref
|
|
|
|
|
Rosa LH, Gonçalves VN, Caligiorne RB, Alves TMA, Rabello A, Sales PA, Romanha AJ, Sobral MEG, Rosa CA, Zani CL (2010). Leishmanicidal, trypanocidal and cytotoxic activities of endophytic fungi associated with bioactive plants in Brazil. Brazilian Journal of Microbiology 41(2):420-430.
Crossref
|
|
|
|
|
Santiago IF, Alves TMA, Rabello A, Junior PAS, Romanha AJ, Zani CL, Rosa CA, Rosa LH (2011). Leishmanicidal and antitumoral activities of endophytic fungi associated with the Antarctic angiosperms Deschampsia antarctica Desv and Colobanthus quitensis (Kunth) Bartl. Extremophiles 16(1):95-103.
Crossref
|
|
|
|
|
Sebastianes FLS, Cabedo N, El Aouad N, Valente AM, Lacava PT, Azevedo JL, Pizzirani-Kleiner AA, Cortes D (2012). 3-Hydroxypropionic acid as an antibacterial agent from endophytic fungi Diaporthe phaseolorum. Current Microbiology 65:622-632.
Crossref
|
|
|
|
|
Sebastianes FLS, Romão-Dumaresq AS, Lacava PT, Harakava R, Azevedo JL, de Melo IS, Pizzinari-Kleiner AA (2013). Species diversity of culturable endophytic fungi from Brazilian mangrove forests. Current Genetic 59:153-166.
Crossref
|
|
|
|
|
Sierra GA (1957). A simple method for the detection of lypolitic activity of microorganisms and observations on the influence of the contact between cells and fatty substracts. Antonie van Leeuwenhoeck 23:15-22.
Crossref
|
|
|
|
|
Strobel G (2018). The emergence of endophytic microbes and their biological promise. Journal of Fungi 4:5.
Crossref
|
|
|
|
|
Strobel G, Daisy B (2003). Bioprospecting for microbial endophytes and their natural products. Microbiology and Molecular Biology Reviews 67:491-502.
Crossref
|
|
|
|
|
Teather RM, Wood PJ (1982). Use of Congo red-polysaccaride interactions inenumeration and characterization of cellulolytic bacteria from bovine rumen. Applied and environmental microbiology 43:777-780.
Crossref
|
|
|
|
|
Thatoi H, Behera BC, Mishra RR (2013). Ecological role and biotechnological potential of mangrove fungi: a review. Mycology 4:54-71.
|
|
|
|
|
Tripathi S, Kamal S, Sheramati R, Oelmuller R, Varma A (2008). Mycorrhizal fungi and other root endophytes as biocontrol agents against root pathogens. Mycorrhiza 3:281-306.
Crossref
|
|
|
|
|
Yan L, Zhao H, Zhao X, Xu X, Di Y, Jiang C, Shi J, Shao D, Huang Q, Yang H, Jin M (2018). Production of bioproducts by endophytic fungi: chemical ecology, biotechnological applications, bottlenecks, and solutions. Applied Microbiology and Biotechnology 102:6279-6298.
Crossref
|
|
|
|
|
Zheng Y, Qiao X, Miao C, Liu K, Chen Y, Xu L, Zhao L (2015). Diversity, distribution and biotechnological potential of endophytic fungi. Annals of Microbiology 66:529-542.
Crossref
|
|