Full Length Research Paper

ABSTRACT
The fungus-cultivating termites (Macrotermitinae) form part of diverse termite fauna in Africa, but information on their fungal symbionts is inadequate and poorly understood. In this study, the fungal communities and structure between termite gut, mound and surrounding soil were determined using the 454 pyrosequencing-based analysis of the internal transcribed spacer (ITS) gene sequences. Genomic DNA was extracted and purified from the guts of three termites (Odontotermes sp., Macrotermes michaelseni and Microtermes sp.), mound and surrounding soil samples for high-throughput sequencing. A total of 15,256 sequences were obtained and individual samples contained between 4 and 133 operational taxonomic units (OTUs). Termite gut had the least fungal diversity, dominated by members of the Basidiomycota (> 98%). More than 98% of the gut sequences were of the genus Termitomyces, while < 2% were related to the genera Chaetomium, Fusarium, Eupenicillium, Cladosporium, Curreya and Phaeosphaeria with between 95 and 98% pair-wise sequence identities. Members of Ascomycota (> 94%) were the most abundant in the mound and soil, but significantly differed (P value of 0.04; R value = 0.909) between the mound and soil environments. The results confirm that the genus Termitomyces exist in a tight association with their hosts and that Termitomyces species are scarcely present in the mound and soil. In addition, by altering soil properties; the fungus-cultivating termites modify the fungal community composition and structure in the mound and surrounding soil environments.
Key words: 454-pyrosequencing, microtermitinae, mutualism, tropical mycology.
INTRODUCTION
MATERIALS AND METHODS
RESULTS
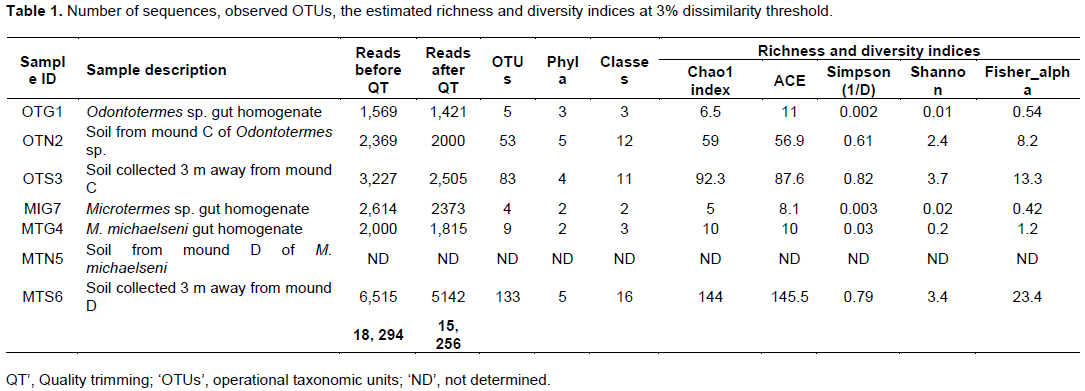
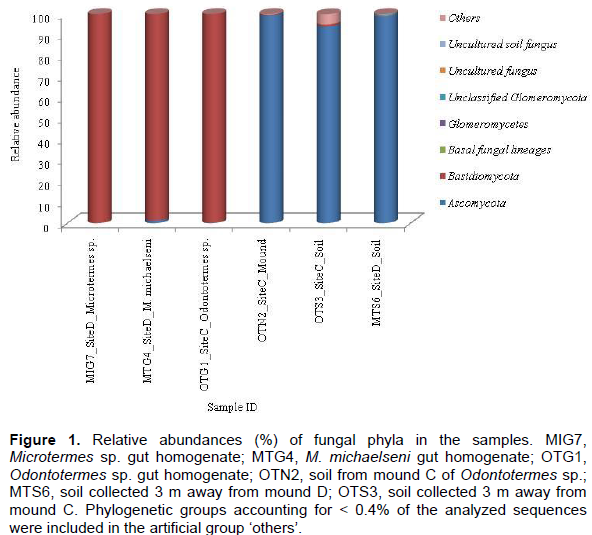
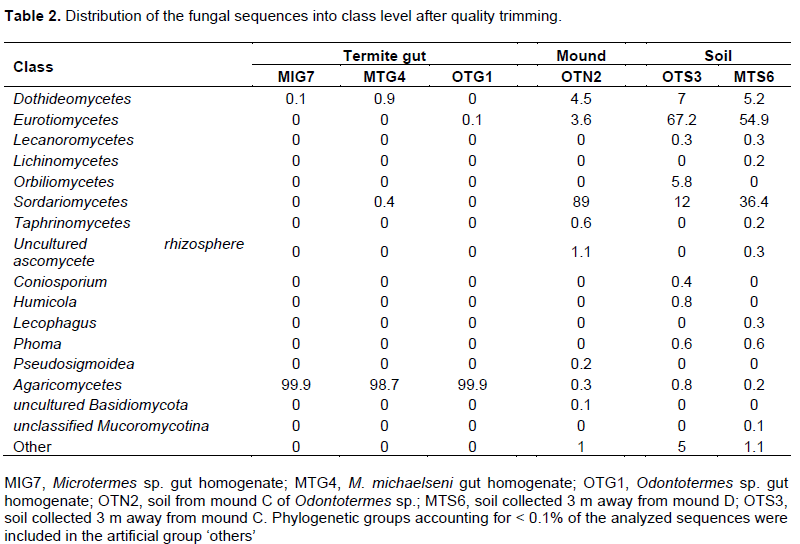
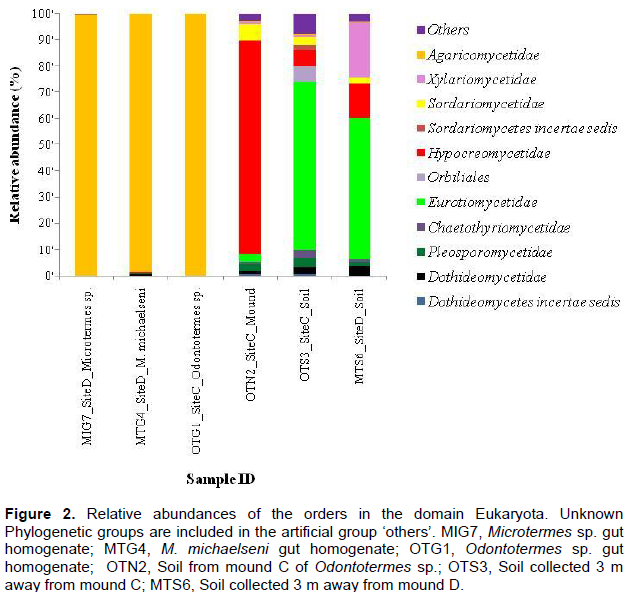
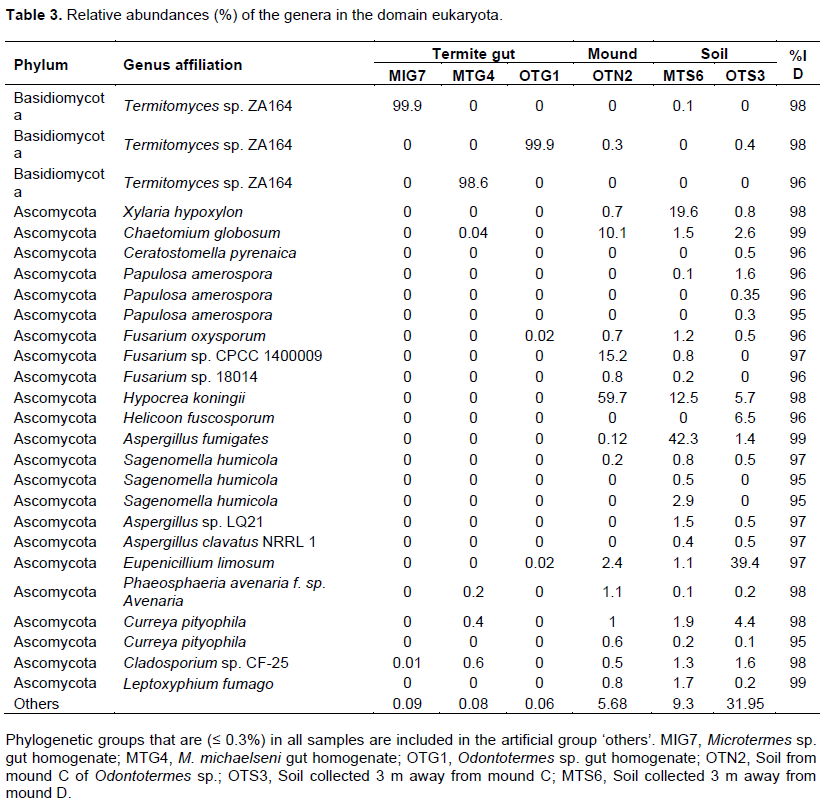
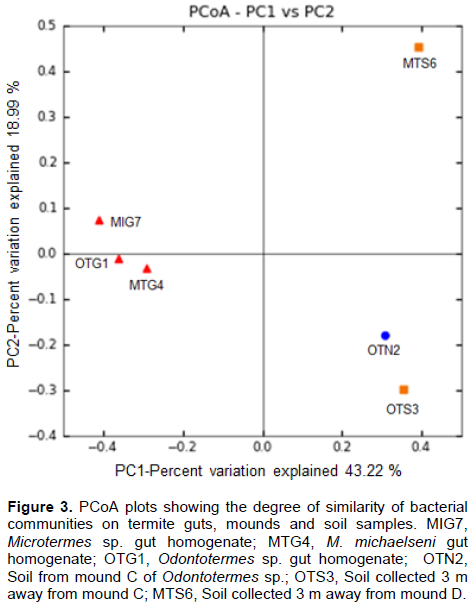
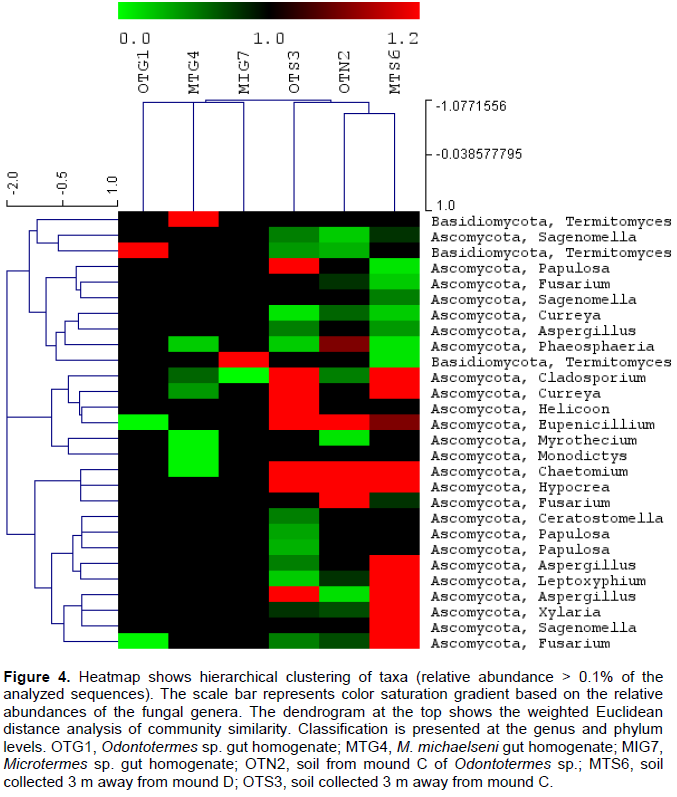
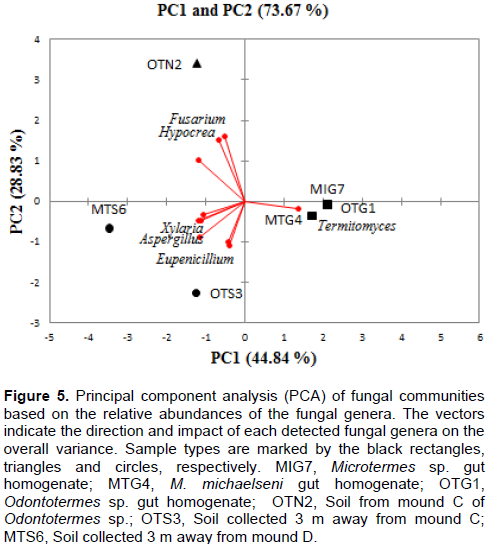
DISCUSSION
CONCLUSION
CONFLICT OF INTERESTS
REFERENCES
|
Aanen DK, de Fine Licht HH, Debets AJM, Kerstes NAG, Hoekstra RF, Boomsma JJ (2009). High symbiont relatedness stabilizes mutualistic cooperation in fungus growing termites. Science. 326:1103-1106. |
|
|
Aanen DK, Ros VID, De Fine Licht HH, Mitchell J, de Beer ZW, Slippers B, Rouland-LeFevre C, Boomsma JJ (2009). Patterns of interaction specificity of fungus-growing termites and Termitomyces symbionts in South Africa. BMC Evol. Biol. 7(115):1-11. |
|
|
Ahmed BM, Nkunika POY, Sileshi WG, French JRJ, Nyeko P, Jain S (2011). Potential impact of climate change on termite distribution in Africa. Br. J. Environ. Clim. Change. 1:172-189. |
|
|
Austin JW, Szalanski AL, Cabrera BJ (2004). Phylogenetic analysis of the subterranean termite family Rhinotermitidae (Isoptera) by using the mitochondrial cytochrome oxidase II gene. Ann. Entomol. Soc. Am. 97:548-555. |
|
|
Barnett HL, Hunter BB (1972). Illustrated Genera of lmperfect Fungi. 3rd Ed. Burgess, Min- neapolis. P 241. |
|
|
Bridge P, Spooner B (2001). Soil fungi: diversity and detection. Plant Soil. 232:147-154. |
|
|
Caporaso JG, Kuczynski J, Stombaugh J, Bittinger K, Bushman FD, Costello EK, Fierer N, Pe-a AG, Goodrich JK, Gordon JI, Huttley GA, Kelley ST, Knights D, Koenig JE, Ley RE, Lozupone CA, McDonald D, Muegge BD, Pirrung M, Reeder J, Sevinsky JR, Turnbaugh PJ, Walters WA, Widmann J, Yatsunenko T, Zaneveld J, Knight R (2010). QIIME allows analysis of high-throughput community sequencing data. Nat. Meth. 7:335-336. |
|
|
Chen DM, Cairney JWG (2002). Investigation of the influence of prescribed burning on ITS profiles of ectomycorrhizal and other soil fungi at three Australian sclerophyll forest sites. Mycol. Res. 106:532-540. |
|
|
Clarke KR (1993). Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 18:117-143. |
|
|
Collins NM (1983). The utilization of nitrogen sources by termites (Isoptera). In: Nitrogen as an Ecological Factor. Eds. Lee JA, McNeill S, Rorison H. 22nd Symposium of British Ecological Society, Oxford. pp. 381-412. |
|
|
De Bruyn LLA, Conacher AJ (1990). The role of termites and ants in soil modiï¬cation: A review. Aust. J. Soil. Res. 28:55-93. |
|
|
Edgar RC (2010). Search and clustering orders of magnitude faster than BLAST. Bioinformatics 26:2460-2461. |
|
|
Fierer N, Lauber CL, Zhou N, McDonald D, Costello EK, Knight R (2010). Forensic identification using skin bacterial communities. Proc. Natl. Acad. Sci. USA. 107(14):6477-6481. |
|
|
Garnier-Sillam E, Toutain F, Renoux J (1988). Comparison of the action of two termite colonies (soil feeding and fungus growing species) on the structural stability of tropical forest soils. Pedobiology 32:89-97. |
|
|
Guedegbe HJ, Miambi E, Pando A, Houngnandan P, Rouland-Lefevre C (2009). Molecular diversity and host specificity of termite-associated Xylaria. Mycologia. 101: 686-691. |
|
|
Harry M, Jusseaume N, Gambier B, Garnier-Sillam E (2001). Use of RAPD markers for the study of microbial community similarity from termite mounds and tropical soils. Soil. Biol. Biochem. 33:417-427. |
|
|
Hawksworth DL (2001). The magnitude of fungal diversity: the 1.5 million species estimate revisited. Mycol. Res. 105:1422-1432. |
|
|
Holt JA, Lepage M (2000). Termites and soil properties. In Termites: Evolution, Sociality, Symbioses, Ecology. Eds. T. Abe T, Bignell DE, Higashi M. Kluwer Academic Publishers Dordrecht. pp. 25-51. |
|
|
Huse SM, Huber JA, Morrison HG, Sogin ML, Welch DM (2007). Accuracy and quality of massively parallel DNA pyrosequencing. Genome Biol. 8(7):R143. |
|
|
Hyodo F, Inoue T, Azuma JI, Tayasu I, Abe T (2000). Role of the mutualistic fungus in lignin degradation in the fungus-growing termite Macrotermes gilvus (Isoptera; Macrotermitinae). Soil. Biol. Biochem. 32:653-658. |
|
|
Hyodo F, Tayasu I, Inoue T, Azuma JI, Kudo T, Abe T (2003). Differential role of symbiotic fungi in lignin degradation and food provision for fungus-growing termites (Macrotermitinae: Isoptera). Func. Ecol. 17:186-193. |
|
|
Inward DJG, Vogler AP, Eggleton P (2007). A comprehensive phylogenetic analysis of termites (Isoptera) illuminates key aspects of their evolutionary biology. Mol. Phylogenet. Evol. 44:953-967. |
|
|
Jones CG, Lawton JH, Shachak M (1997). Positive and negative effects of organisms as physical ecosystem engineers. Ecology 78:1946-1957. |
|
|
Ju YM, Hsieh HM (2007). Xylaria species associated with nests of Odontotermes formosanus in Taiwan. Mycologia. 99:936-957. |
|
|
Lamberty M, Zachary D, Lanot R, Bordereau C, Robert A, Hoffmann JA, Bulet P (2001). Insect immunity: Constitutive expression of a cysteine-rich antifungal and a linear antibacterial peptide in a termite insect. J. Biol. Chem. 276:4085-4092. |
|
|
Landeweert R, Leeflang P, Kuyper TW, Hoffland E, Rosling A, Wernars K, Smit E (2003). Molecular identification of ectomycorrhizal mycelium in soil horizons. Appl. Environ. Microbiol. 69:327-333. |
|
|
Lavelle P, Bignell D, Lepage M, Wolters V, Roger P, Ineson P, Heal OW, Dhillion S (1997). Soil function in a changing world: the role of invertebrate ecosystem engineers. Eur. J. Soil. Biol. 33:159-193. |
|
|
Lozupone C, Knight R (2005). UniFrac: a new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 71:8228-8235. |
|
|
Lozupone CA, Hamady M, Kelley ST, Knight R (2007). Quantitative and qualitative (beta) diversity measures lead to different insights into factors that structure microbial communities. Appl. Environ. Microbiol. 73(5):1576-1585. |
|
|
Makonde HM, Boga HI, Osiemo Z, Mwirichia R, Stielow JB, Göker M, Klenk HP (2013). Diversity of Termitomyces Associated with Fungus-Farming Termites Assessed by Cultural and Culture-Independent Methods. PLoS ONE. 8(2):e56464. |
|
|
Manuwa SI (2009). Physico-chemical and dynamic properties of termite mound soil Relevant in sustainable food production. Afr. Crop Sci. Soc. 9:356-369. |
|
|
Martin MM, Martin JS (1978). Cellulose digestion in the midgut of the fungus-growing termites Macrotermes natalensis: The role of acquired digestive enzymes. Science 199:1453-1455. |
|
|
Mathew GM, Ju Y-M, Lai C-Y, Mathew DC, Huang CC (2012). Microbial community analysis in the termite gut and fungus comb of Odontotermes formosanus: the implication of Bacillus as mutualists. FEMS Microbiol. Ecol. 79:504-517. |
|
|
McLean MA, Parkinson D (2000). Field evidence of the effects of the epigeic earthworm Dendrobaena octaedra on the microfungal community in pine forest floor. Soil Biol. Biochem. 32:351-360. |
|
|
Mohindra P, Mukerji KG (1982). Fungal ecology of termite mounds. Rev. Ecol. Biol. Sol. 19:351-361. |
|
|
Moriya S, Inoue T, Ohkuma M, Yaovapa T, Johjima T, Suwanarit P, Sangwani U, Vongkaluang, C, Noparatnaraporn N, Kudo T (2005). Fungal Community Analysis of Fungus gardens in termite nests. Microbes Environ. 20:243-252. |
|
|
Muwawa EM, Budambula NLM, Osiemo ZL, Boga HI, Makonde HM (2016). Isolation and characterization of some gut microbial symbionts from fungus-cultivating termites (Macrotermes and Odontotermes spp.). Afr.J. Microbiol. Res. 10(26):994-1004. |
|
|
Muwawa EM, Makonde HM, Budambula NLM, Osiemo LZ, Boga HI (2014). Chemical properties associated with guts, soil and nest materials of Odontotermes and Macrotermes species from Kenya. J. Biodiv. Environ. Sci. 4(2):253-263. |
|
|
Nobre T, Eggleton P, Aanen DK (2010). Vertical transmission as the key to the colonization of Madagascar by fungus growing termites? Proceedings of the Royal Society B: Biol. Sci. 277:359-365. |
|
|
Nobre T, Fernandes C, Boomsma JJ, Korb J, Aanen DK (2011). Farming termites determine the genetic population structure of Termitomyces fungal symbionts. Mol. Ecol. 20:2023-2033. |
|
|
Okane I, Nakagiri A (2007). Taxonomy of an anamorphic xylariaceous fungus from a termite nest found together with Xylaria angulosa. Mycoscience 48:240-249. |
|
|
Osiemo Z, Marten A, Kaib M, Gitonga L, Boga H, Brandl R (2010). Open relationships in the castles of clay: high diversity and low host specificity of Termitomyces fungi associated with fungus-growing termites in Africa. Insect Soc. 57:351-363. |
|
|
Otani S, Hansen LH, Sørensen SJ, Poulsen M (2015). Bacterial communities in termite fungus combs are comprised of consistent gut deposits and contributions from the environment. Microb. Ecol. 71(1):207-20. |
|
|
Otani S, Mikaelyan A, Nobre T, Hansen LH, Koné NGA, Sørensen SJ, Aanen DK, Boomsma JJ, Brune A, Poulsen M (2014). Identifying the core microbial community in the gut of fungus-growing termites. Mol. Ecol. 23:4631-4644. |
|
|
Quast C, Pruesse E, Yilmaz P, Gerken J, Schweer T, Yarza P, Peplies J, Glöckner FO (2013). The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucl. Acids. Res. 41(DI):D590-D596. |
|
|
R Core Team (2012). R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. Vienna, Austria. URL http://www.R-project.org/. |
|
|
Rideout JR, He Y, Navas-Molina JA, Walters WA, Ursell LK, Gibbons SM, Chase J, McDonald D, Gonzalez A, Robbins-Pianka A, Clemente JC, Gilbert JA, Huse SM, Zhou H, Knight R, Caporaso JG (2014). Subsampled open-refer-ence clustering creates consistent, comprehensive OTU definitions and scales to billions of sequences. Peer J. 2:e545. |
|
|
Roose-Amsaleg C, Brygoo Y, Harry M (2004). Ascomycete diversity in soil-feed-ing termite nests and soils from a tropical rainforest. Environ. Microbiol. 6:462-469. |
|
|
Rouland-Lefévre C (2000). Symbiosis with fungi. In: Termites: evolution, society, symbioses, ecology. Eds. Abe T, Bignell DE, Higashi M. Kluwer Academic Publishers, Dordrecht, The Netherlands. pp. 289-306. |
|
|
Steinhaus EA (1949). Principles of Insect Pathology. McGraw-Hill, New York. P 757. |
|
|
Traniello JFA, Rosengaus RB, Savoie K (2002). The development of immunity in a social insect: Evidence for the group facilitation of disease resistance. Proc. Natl. Acad. Sci. USA. 99:6838-6842. |
|
|
Visser AA, Nobre T, Currie CR, Aanen DK, Poulsen M (2012). Exploring the potential for actinobacteria as defensive symbionts in fungus-growing termites. Microb. Ecol. 63: 975-985. |
|
|
Visser AA, Ros VID, De Beer ZW, Debets AJM, Hartog E, Kuyper TW, Laessøe T, Slippers B, Aanen DK (2009). Levels of specificity of Xylaria species associated with fungus-growing termites: A phylogenetic approach. Mol. Ecol. 18:553-567. |
|
|
Waid JS (1999). Does soil biodiversity depend upon metabiotic activity and influences? Appl. Soil Ecol. 13:151-158. |
|
|
White TJ, Bruns TD, Lee S, Taylor JW (1990). Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics In: Innis MA, Gelfand DH, Sninsky JJ, White TJ (eds) PCR Protocols: a guide to methods and applications. New York: Academic Press. pp. 315-322. |
|
|
William AS (1969). The association of termite and fungi In: Kummer K, Frances MW (eds) Biology of the termite. Academic Press, New York, USA, pp. 495-524. |
|
|
Zoberi MH, Grace JK (1990). Fungi associated with the subterranean termite Reticulitermes flavipes in Ontario. Mycologia. 82:289-294. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


