Full Length Research Paper

ABSTRACT
Ten Trichoderma isolates were isolated from different locations in Egypt. Amplification and sequencing of internal transcribed spacers (ITS) was employed to identify Trichoderma isolates that exhibited from 99 to 100% identity with three species of Trichoderma: Trichoderma harzianum, Trichoderma asperellum and Trichoderma longibrachiatum. The biocontrol activity of Trichoderma isolates against Macrophomina phaseolina, Aspergillus niger and Meloidogyne incognita was tested in vitro and under greenhouse conditions. The results show that the isolate Th2 (T. harzianum) gave the best antagonism against M. phaseolina and A. niger with inhibition rates of 72.85 and 64.28%, respectively. Moreover, the isolate Ta1 (T. asperellum) was the best efficient isolate in reduction of each second stage juveniles (J2), number of galls, egg masses and females per root system with 90.33, 90.59, 90.06 and 85.50%, respectively. Treatment with Trichoderma isolates improved tomato growth parameters (root length, plant height, roots and shoots fresh weight and shoots dry weight).
Key words: Trichoderma, internal transcribed spacers (ITS), antagonism, Macrophomina phaseolina, Aspergillus niger, Meloidogyne incognita.
INTRODUCTION
MATERIALS AND METHODS
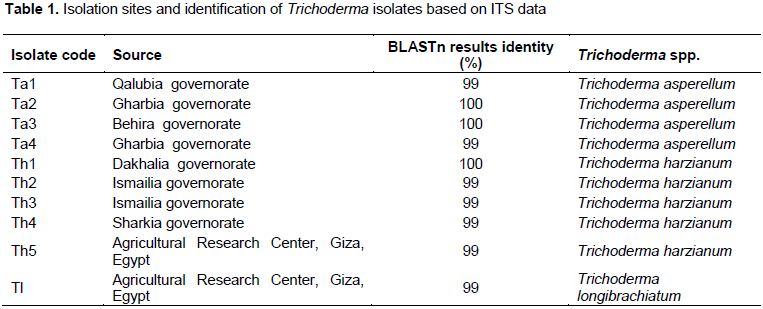
RESULTS AND DISCUSSION

CONCLUSION
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Abdel-lateif K, El-Zanaty AF, Helwa M (2017). Genetic diversity of Trichoderma isolates and their antagonism against Rhizoctonia Solani and Phythium aphnidermatum. Menoufia Journal of Agriculture Biotechnology 2:63-73. |
|
|
Agrawal T, Kotasthane AS (2012). Chitinolytic assay of indigenous Trichoderma isolates collected from different geographical locations of Chhattisgarh in Central India. Springer Plus 1:73-82. |
|
|
Al-Hazmi AS, Javeed MT (2016). Effects of different inoculum densities of Trichoderma harzianum and Trichoderma viride against Meloidogyne javanicaon tomato. Saudi Journal of Biological Sciences 23:288-292 |
|
|
Al-Samarrai TH, Schmid J (2000). A simple method for extraction of fungal genomic DNA. Letters in Applied Microbiology 30: 53-56. |
|
|
Aly AA, Abdel-Sattar MA, Omar MR, Abd-Elsalam KA (2007). Differential antagonism of Trichoderma sp. against Macrophomina phaseolina. Journal of Plant Protection Research 47(2). |
|
|
Athira K (2017). Efficacy of fungicide and bio-control agents against root rot of black Gram (Vigna mungo L.) caused by Macrophomina phaseolina (Tassi) Goid. International Journal of Current Microbiology and Applied Sciences 6(10):2601-2607. |
|
|
Bakr RA, Mahdy ME, Mousa EM (2011). A survey of root-knot and citrus nematodes in some new reclaimed lands in Egypt. Pakistan Journal of Nematology 29:165-170. |
|
|
Benítez T, Rincón AM, Limón MC, Codón AC (2004). Biocontrol mechanisms of Trichoderma strains. International Microbiology 7:249-260 |
|
|
Bhale UN, Wagh PM, Rajkonda JN (2013). Antagonistic confrontation of Trichoderma spp against fruit rot pathogens on Sapodilla (Manilka razapota L.). Journal of Yeast and Fungal Research 4(1):5-11 |
|
|
Chen S, Dickson DW (2004). Biological control of nematodes by fungal antagonists. In: Nematology Advances and Perspectives. Nematode Management and Utilization, vol. II. (Chen ZX, Chen SY, Dickson DW, eds).Wallingford: CAB International, UK pp. 977-1039. |
|
|
Daykin ME, Hussey RS (1985). Staining and histopathological techniques in nematology. pp. 39-48. In: K. R. Barker, C. C. Carter, J. N. Sasser, eds. An Advanced Treatise on Meloidogyne: Volume II. Raleigh, NC: North Carolina State University Graphics. |
|
|
Duncan B (1955). Multiple range and Multiple F test. Biometrex 11:1-42. |
|
|
Elad Y, Hadar Y, Chet I, Henis Y (1981). Biological control of Rhizoctonia solani by Trichoderma harzianum in carnation. Plant Disease 65:675- 677. |
|
|
El-Hassan SA, Gowen SR, Pembroke B (2013). Use of Trichoderma hamatum for biocontrol of lentil vascular wilt disease: efficacy, mechanisms of interaction and future prospects. Journal of Plant Protection Research 53(1). |
|
|
Epstein E (1972). Mineral Nutrition of Plants: Principles and Perspectives. John Wiley, New York. |
|
|
Fahmi AI, Eissa RA, El-Halfawi KA, Hamza HA, Helwa MS (2016). Identification of Trichoderma spp. by DNA Barcode and Screening for Cellulolytic Activity. Journal of Microbial and Biochemical Technology 8:202-209. |
|
|
Gajera H, Kalu K, Vakharia D (2011). Bioefficacy of Trichoderma isolates against Aspergillus niger Van Tieghem inciting collar rot in groundnut (Arachis Hypogaea L.). Journal of Plant Protection Research 51(3). |
|
|
Gajera HP, Bambharolia RP, Patel SV, Khatrani TJ, Goalkiya BA (2012). Antagonism of Trichoderma spp. against Macrophomina phaseolina: Evaluation of coiling and cell wall degrading enzymatic Activities. Journal of Plant Pathology and Microbiology 3:149. |
|
|
Goody JB (1957). Laboratory method for work with plant and soil nematodes. Ministry of Agric., Fisheries and Food: Tech. Bull. 2 London, England 44 p. |
|
|
Haran S, Schickler H, Oppenheim A, Chet I (1995). New components of the chitinolytic system of Trichoderma harzianum. Mycological Research 99:441-446. |
|
|
Hartman K, Sasser JN (1985). Identification of Meloidogyne species on the basis of differential hosts test and perineal pattern morphology. In: Barker KR, Carter CC, Sasser JN, editors. An advanced treatise on Meloidogyne. Volume II. North Carolina State University Graphics, Raleigh, North Carolina: Methodology. A cooperative publication of the Department of Plant Pathology and the United States Agency for International Development pp. 69-77. |
|
|
Hassan M, EL-fiky ZA, Said MA, Ahmed NF (2015). Chitinase and cellulase genes sequencing for some Egyptian Trichoderma species isolated from rhizosphere and assay of their activity. Egyptian Journal of Genetics and Cytology 44:15-30. |
|
|
Hussey RS, Barker KR (1973). Comparison of methods for collecting inocula of Meloidogyne spp., including a new technique. Plant Disease Report 57:1025-1028. |
|
|
Ike M, Nagamatsu K, Shioya A, Nogawa M, Ogasawara W, Okada H, Morikawa Y (2006). Purification, characterization, and gene cloning of 46 kDa chitinase (Chi 46) from Trichoderma reesei PC-3-7 and its expression in Escherichia coli. Applied Microbiology and Biotechnology 71(3): 294-303. |
|
|
Izuogu NB, AbiriTO (2015). Efficacy of Trichoderma harzianum T22 as a biocontrol agent against root- knot nematode (Meloidogyne incognita) on some soybean varieties. Croatian Journal of Food Science and Technology 7(2):47-51. |
|
|
Jansson HB, Jeyaprakash A, Zuckerman BM (1985). Control of root-knot nematodes on tomato by the endoparasitic fungus Meriaconiospora. Journal of Nematology 17:327-329. |
|
|
Jiang H, Zhang L, Zhang J, Ojaghian MR, Hyde KD (2016). Antagonistic interaction between Trichoderma asperellum and Phytophthora capsici in vitro. Journal of Zhejiang University-Science B 17(4): 271-281. |
|
|
Junaid JM, Dar NA, Bhat TA, Bhat AH, Bhat MA (2013). Commercial biocontrol agents and their mechanism of action in the management of plant pathogens. International Journal of Modern Plant and Animal Sciences 1(2): 39-57. |
|
|
Karssen G, Moens M (2006). Root-knot nematodes. In: Perry, R.N. and Moens, M. (Eds). Plant Nematology. CABI publishing pp. 59-90 |
|
|
Katooli N, Moghadam EM, Taheri A, Nasrollahnejad S (2010). Management of root knot nematode (Meloidogyne incognita) on cucumber with the extract and oily of nematicidal plants. International Journal of Agricultural Research 5:582-586. |
|
|
Khaledi N, Taheri P (2016). Biocontrol mechanisms of Trichoderma harzianum against soybean charcoal rot caused by Macrophomina phaseolina. Journal of Plant Protection Research 56(1). |
|
|
Khalili E, Javed MA, Huyop F, Rayatpanah S, Jamshidi S, Abdul Wahab R (2016). Evaluation of Trichoderma isolates as potential biological control agent against soybean charcoal rot disease caused by Macrophomina phaseolina. Biotechnology and Biotechnological Equipment 30(3):479-488 |
|
|
Khokhar I, Haider MS, Mukhtar I, Mushtaq S (2012). Biological control of Aspergillus niger, the cause of Black-rot disease of Allium cepa L. (onion), by Penicillium species. Journal of Agriculture and Biology 29(1):23-28. |
|
|
Krishnamurthy YL, Shashikala J (2006) Inhibition of aflatoxin B1 production of Aspergillus flavus, isolated from soybean seeds by certain natural plant products. Letters in Applied Microbiology 43:469-474. |
|
|
Loc NH, Quang HT, Hung NB, Huy ND, Phuong TT, Ha TTT (2011). Trichoderma asperellum Chi42 genes encode chitinase. Mycobiology 39(3): 182-186 |
|
|
Maqbool MA, Kerry B (1997). Plant Nematodes Problems and their Control in the Near East Region. Proc. Expert Consultation of Plant Nematode Problems and Their Control in the Near East Region Karachi, Pakistan 315 p. |
|
|
Mascarin GM, Junior MFB, Filho JVdeA (2012). Trichoderma harzianum reduces population of Meloidogyne incognita in cucumber plants under greenhouse conditions. Journal of Entomology and Nematology 4:54-57. |
|
|
Mendoza JLH, Pérez MIS, Prieto JMG, Velásquez JDQ, Olivares JGG, Langarica HRG (2015). Antibiosis of Trichoderma spp strains native to northeastern Mexico against the pathogenic fungus Macrophomina phaseolina. Brazilian Journal of Microbiology 46(4):1093-1101 |
|
|
Njoki LM, Okoth SA, Wachira PM (2017). Effects of medicinal plant extracts and photosensitization on aflatoxin producing Aspergillus flavus (Raper and Fennell). International Journal of Microbiology 5:1-9. |
|
|
Oskiera M, Szczech M, Bartoszewski G (2015). Molecular identification of Trichoderma strains collected to develop plant growth-promoting and biocontrol agents. Journal of Horticulture Research 23(1): 75-86. |
|
|
Pal KK, Gardener BM (2006). Biological Control of Plant Pathogens. The Plant Health Instructor. |
|
|
Pandya JR, Sabalpara AN, Chawda SK (2011). Trichoderma: a particular weapon for biological control of Phytopathogens. Journal of Agricultural Technology 7(5): 1187-1191. |
|
|
Prasetyawan S, Sulistyowati L, Aulanni'am (2018). Glucanase and chitinase from some isolates of endophytic fungus Trichoderma spp. IOP Conference Series: Materials Science and Engineering 299:012026 |
|
|
Rao KLNM, Raju KS, Ravisankar H (2015). Cultural conditions on the production of extracellular enzymes by Trichoderma isolates from tobacco rhizosphere. Brazilian Journal of Microbiology 47(1):25-32 |
|
|
Rayatpanah S, Nanagulyan SG, Alav SV, Razavi M, Ghanbari-Malidarreh A (2011). Pathogenic and genetic diversity among Iranian isolates of Macrophomina phaseolina. Chilean Journal of Agricultural Research 72:40-44. |
|
|
Savitha MJ, Sriram S (2015). Morphological and molecular identification of Trichoderma isolates with biocontrol potential against Phytophthora blight in red pepper. Pest Managment in Horticulture Ecosystems 21(2):194-202. |
|
|
Seidl-Seiboth V, Ihrmark K, Druzhinina I, Karlsson M (2014). Molecular evolution of Trichoderma chitinases. Biotechnology and Biology of Trichoderma 2014:67-78. |
|
|
Sharma RA (2012). Brief review on mechanism of Trichoderma fungus use as biological control agents. International Journal of Innovations in Biological Sciences 2:200-210. |
|
|
Sharon E, Chet I, Spiegel Y (2011). Trichoderma as a biological control agent. In: Davies, K., Spiegel, Y. (Eds.), Biological Control of Plant-Parasitic Nematodes: Building Coherence between Microbial Ecology and Molecular Mechanisms, Progress in Biological Control. Springer, Netherlands pp. 183-201. |
|
|
Shoaib A, Munir M, Javaid A, Awan ZA, Rafiq M (2018). Anti-mycotic potential of Trichoderma spp. and leaf biomass of Azadira chtaindica against the charcoal rot pathogen, Macrophomina phaseolina (Tassi) Goid in cowpea. Egyptian Journal of Biological Pest Control 28:26. |
|
|
Sikora RA, Fernandez E (2005). Nematode parasites of vegetables, In: Luc, M., Sikora, R.A, Bridge, J. (Eds.). Plant Parasitic Nematodes in Subtropical and Tropical Agriculture, CABI Publishing, Wallingford, UK pp. 319-392. |
|
|
Spiegel Y, Sharon E, Bar-Eyal M (2007). Evaluation and mode of action of Trichoderma isolates as biocontrol agents against plant-parasitic nematodes. IOBC. WPRS. Bulletin 30:129-133. |
|
|
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S (2013). MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Molecular Biology and Evolution 30:2725-2729. |
|
|
Woo SL, Ruocco M, Vinale F, Nigro M, Marra R, Lombardi N, et al (2014). Trichoderma-based Products and their Widespread Use in Agriculture. The Open Mycology Journal 8(1). |
|
|
Wu Q, Sun R, Ni M, Yu J, Li Y, Yu C, Dou K, Ren J, Chen J (2017). Identification of a novel fungus, Trichoderma asperellum GDFS1009, and comprehensive evaluation of its biocontrol efficacy. PLOS ONE 12(6):e0179957. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


