Full Length Research Paper

ABSTRACT
Mosquito larvae feed on particulate organic matter including microorganisms. This study was conducted to investigate the diversity and abundant bacteria of Anopheles mosquito larva breeding habitats and to evaluate the contribution of maize pollen as source of nutrients for bacterial growth. The nutrient composition (COD, NH3-N and TP) of the larva breeding habitats water samples were measured by HACH 2010. Bacteria were isolated and enumerated from 18 water samples of larva habitats. The abundance of bacteria in the larva breeding habitats was significantly different. Bacteria were found to be grown abundant in habitats close to maize pollen sources (tasseled zone). This implies that maize pollen contributes to bacterial abundance. The Pearson Correlation showed that there was positive relationship between bacterial abundance and physicochemical characteristics of the water samples. The bacterial population in the habitat was dominated by species of Bacillus, Pseudomonas, Micrococcus and Serratia. The dominant bacteria were tested for their capability to grow on maize pollen medium. The growth kinetics of bacteria on maize pollen broth was performed to 18 h culture using JENWAY spectrophotometer at 600 nm wave length. The bacteria could show optimum growth on Maize Pollen broth at 15 g/l as of nutrient broth. The release of maize pollen during anthesis in rainy season in habitats close to larva breeding pool and its nutriment quality support proliferation of large array of bacteria which results in increased larval nourishment. Increased malaria transmission in Asendabo could thus be caused as the bacteria serve as source of nutrients for mosquito larva.
Key words: Maize pollen, microbial flora, mosquito larva habitat, maize pollen broth, bacterial abundance.
INTRODUCTION
Mosquitoes are vectors of pathogenic protozoan, viruses, and nematodes which cause the disease malaria, yellow fever, and lymphatic filariasis respectively to humans (Okogun et al., 2003; Maekawa et al., 2011) to humans and related diseases in domestic animals. Anopheles are medically important mosquito, transmit malaria disease. The genus Anopheles comprises of 421 species, which are distributed in the world (Kettle, 1995). They breed in many types of temporary water bodies, but differ in preferences to particular types of water body (Mwangangi et al., 2007; Machault et al., 2009).
Anopheles gambiae is one of the most important vectors of malaria and that is widely distributed through-out Afro tropical region. It breeds in small, sun exposed, fresh water pools which are often free of vegetation (Muirhead-Thomson, 1958). The larvae of Anopheles gambiae are also found at the coast in intertidal, salt water swamps (Ketlle, 1995).
The larval stages of A. gambiae complex are frequently found in transient water bodies, where suspended solid particles are abundant to provide turbidity. The other Anopheline species such as Anopheles quadrimaculatus develop in relatively clear water by exploiting bacteria-rich surface environment of permanent marshes through their interfacial feeding strategy (Yemane et al., 2000).
Bacteria (Cyanobacteria) are widely distributed in mosquito breeding habitats and used as food for mosquito larvae. The size of adult mosquito population is largely dependent on the presence of plankton (Mala and Irungu, 2011), the nutritional role of organic matter, and microbial fauna in the mosquito larval habitats (Okech et al., 2007). Larval food consists of a variety of living and non-living materials, such as dust, bacteria (Cyanobacteria), unicellular algae (zoo flagellates) and filamentous algae, fungi and small metazoans (rotifers and crustaceans), spores and in-sect scales. Bacteria are the most important microbial constituents of mosquito larvae food and the mosquito can grow on culture made only of bacteria (Merritt et al., 1992).
The amount of organic matter and nutrient in the water is important factors in temporary small water bodies of the habitats of many Anopheline mosquitoes. In these shallow habitats there is a close association between the organic matter in the water and the organic content of the underlying soil (Muirhead-Thomson, 1958).
Maize (Zea mays L., Poaceae) is the most important cereal in the world after wheat and rice with regard to cultivation areas and total production (Abdulrahaman and Kolawole, 2006). The name maize is derived from the South American Indian Arawak-Carib word mahiz. It was introduced into Ethiopia probably in the 16th century by the Portuguese (Phillip, 1995).
Maize is the second most important cereal crop grown by Ethiopian farmers (Adugna and Melaku, 2001), with major production regions located in the southern, western, southwestern, and eastern high lands and used predominantly for human consumption in the country (Getahun et al., 2000). It is estimated that the crop covers 1.3 m ha of cultivated land. Maize is used predominantly as human food consumption in the country (Adugna and Melaku, 2001). In general, the area coverage of maize is high in the Oromia region followed by SNNPR and lower in Amhara region (Berhanu et al., 2007). Maize pollen provides important sources of nutrients for adult and larvae of An. arabiensis in Ethiopia (Yemane et al., 2003). The mosquito adult developed more in maize pollen supplemented medium than algal supplemented medium (Yemane et al., 2000). Maize pollen contains various diffusible, water-soluble as well as non-soluble proteins and other nutrients. The nutrient analysis of BH660 Hybrid maize pollen contains 19% protein, 48% carbohydrate, 2% fat, 2% ash and 9% moisture contents Bezawit Eshetu (2007). This maize plant releases its pollen and provides more nutrients to the development of mosquito larvae (Yemane et al., 2003b).
While the microbial biomass associated with different larva aquatic habitats is well documented, the taxonomic composition of these microbes and their response to common environmental stressors is poorly understood (Muturi et al., 2013).
The goal of this research were to study the microbial constituents of Anopheles mosquito larva breeding habitats and evaluate the nutriment contribution of maize pollen to the growth of bacteria, and to investigate the correlation of the larva breeding water micronutrient to the abundance of bacterial isolates.
The hypotheses tested were that maize pollen support different quantities of abundant taxonomic groups of bacteria and exposure of microorganisms to maize pollen source can alter bacterial abundance in mosquito aquatic habitats and its contribution to malaria transmission.
MATERIALS AND METHODS
The nutrient contents of larva breeding water samples were measured and correlated with the abundance of bacteria and hence to the proliferation of mosquito larva. The abundant bacteria were isolated, characterized and identified, and the bacterial abundance was correlated to the availability of maize pollen.
Sample collection
Water samples were collected from eighteen different study sites in Asendabo, Waktola Asendabo (longitude: 37.23", latitude: 7.73" and altitude: 1870 m). Triplicate water samples were collected by using sterile jar, vials, glass bottles and sterile plastic bag. Soon after collection the samples were transported to AAU for microbiological studies and investigations. The maize pollen was collected from Arbaminch, Mirab Abaya (irrigation fed area) (Latitude: 06°18’, longitude: 37°47’), where maize shed the pollen lately. The pollen collected was kept in sterile glass bottle at room temperature until the time of its evaluation as a nutrient source for microbial growth.
Habitat characterization
Sample sites were described in terms of vegetation, physical water condition, exposure of the site to sunlight (Herrel et al., 2001; Barros et al., 2011; Abebe Animut et al., 2012; Afrane et al., 2012).
The study sampling sites were divided in to three zones with regard to the availability of maize pollen source: tasseled zone was a zone with its male maize fluorescence remains attached and pollen was allowed to pollinate by natural process (wind), the detasseled zone was a site from where the male fluorescence was removed and shaken to pollinate and the buffer zone was where the pollen source was of a different form other than maize pollen (taro, teff and green pepper).
Isolation of bacteria from mosquito larva habitats
One milliliter of each water sample was diluted with 9 ml of sterile distilled water in screw caped test tubes up to 10-5 series of dilution. From first to four series of dilution, 0.1 ml aliquots of water sample was spread in to nutrient agar plates and incubated for 48 h at 25°C. Nutrient agar (Oxoid) contains (g/l): ‘Lam-Lemco’ powder 1.0, Yeast extract 2.0, Peptone 5.0, Sodium Chloride 5.0 and Agar 15.0. Each colony was observed on a plate and checked for its purity and the impure colony was transferred to nutrient broth and streaked into nutrient agar plates to re- isolate and purify the bacteria. Following growth, bacterial colonies were further purified and preserved on nutrient agar slants at 4°C for further studies (Fry and Zia, 1982). The total counts of bacterial isolates were enumerated using the formula (number of CFU = number of colonies x Dilution factor of the plate counted). The bacterial isolates were designated as AS6B, AS6C, AS11A, AS12B, AS15B, AS16D, AS17D, AS17E, AS19B and AS20C.
Identification of bacterial isolates
The bacteria isolates were identified by using cultural, morphological, biochemical and physiological characteristics according to Bergey’s Manual of Systematic Bacteriology (1984).
Cultural characteristics of bacterial isolates
The cultural characteristics of bacterial isolates such as colony color, colony elevation, colony margin and colony surface were inspected visually in a plate (Aneja, 2005).
Morphological characteristics of bacterial isolates
Morphological characteristics of bacterial isolates such as spore stain, gram staining, shape and motility tests were determined by light microscopy (Bisen and Verma, 1994; Aneja, 2005).
Biochemical characteristics of bacterial isolates
Biochemical characteristics of isolates, such as Oxidation-Fermentation (OF) test (Bisen and Verma, 1994), Catalase activity (Chester, 1979), urea hydrolysis test, SIM test (Aneja, 2003), MR-VP test, oxidase test and citrate utilization (Bisen and Verma, 1994), carbohydrate fermentation test (Aneja, 2003), hydrolysis of gelatin (Bisen and Verma, 1994; Aneja, 2003), and starch hydrolysis (Aneja, 2003; Mondal et al., 2015) were characterized.
Physiological characteristics of bacterial isolates
For the physiological tests the bacterial isolates were streak inoculated in to nutrient agar medium. The bacterial isolates were inoculated in to the medium with different pH (5, 7, and 10). For salt tolerance tested the bacterial isolates were inoculated into the medium with different salt concentration (2% NaCl, 5% NaCl, 7% NaCl, 10% NaCl and 12% NaCl). The temperature optima was tested by inoculating the isolates at 4, 10, 15c, 40, 45, 50, 55 and 65°C (Aneja, 2003; Bisen and Verma, 1994; Mondal et al., 2015).
Measurement of chemical parameters of water samples
The determination of chemical concentration of the water samples were carried out.
Measurement of ammonia nitrogen
The ammonia nitrogen concentration was measured by Nesslers method. The spectrophotometer was programmed at 380 with wavelength 425 nm. Twenty five ml of water sample was filled in to a 30 ml glass vial (sample cell). To this sample, 1 ml of Nessslers reagent was added, mixed by shaking and allowed to stand for one minute to complete the re-action. All the samples were analyzed for ammonia nitrogen species by taking the reading at HACH 2010 spectrophotometer methods (APHA, 1998).
Measurement of total phosphorous
Total phosphorus (TP) was determined by acid per-sulfate digestion method (APHA, 1998). Five milliliter water sample was added in to total acid hydrolyzable test vial. Potassium per sulfate powder pillow was added and dissolved by shaking. Then, the sample was digested for 30 min at 150°C on COD reactor. After digestion, the digested samples were allowed to cool at room temperature. Two ml of 1.54 normal NAOH was added, mixed and the outside of the tube cleaned with a towel. From this point each sample was treated at a time, zero each vial in HACH 2010 spectrophotometer. One phosver 3-reagent powder pillow was added, shaken for 10 to 15 s. The timer was shift for 2 min; the outside of tube was cleaned with towel and read by using HACH 2010 spectrophotometer (APHA, 1998).
Measurement of COD
The COD reactor was adjusted to 150°C .Two milliliter sample was added to COD vial tube plus one blank. The content of vials was mixed by inverting the vials. The outside of the vial was cleaned with towel and digested for two hours. After two hours of sample digestion the reactor was turned off, the samples were removed from the reactor and allowed to cool at room temperature. The digested cool samples were read on HACH 2010 spectro-photometer programmed at 435 of wave length 620 nm (APHA, 1998).
Evaluation of growth of bacterial isolates in maize pollen
Maize pollen collection and processing
Maize pollen was collected by using hard cover post during anthesis of maize crop. The pollen was put and allowed to dry in open air. The dried pollen powder was shed by hand crushing from the pollen stalk, sieved and kept for future use as media.
Maize pollen broth media preparation
5, 10 and 15 g maize pollen powder was boiled in distilled water (1 L) to dissolve the nutrient contents. After boiling the preparation was allowed to settle. The supernatant decanted from the precipitate was sterilized at 121°C 15 lb pressure for 15 min. The pH of pollen medium was adjusted to 7.2 for bacterial isolate growth evaluation before sterilizing. For Maize pollen agar media preparation, 15 g agar was added to the same preparation as before on another test and was used for physical (visual) evaluation of bacterial growth.
Growth kinetics evaluation
The growth of microorganism on a maize pollen broth media was evaluated by inoculating the isolates on a medium prepared solely of maize pollen. From 18 h culture, 107 number of cells per ml was inoculated into the pollen broth medium (contain 5, 10 and 15 g/l). The optical density (OD) reading was recorded from zero hours of inoculation up to 8 h of incubation within one hour intervals using JENWAY spectrophotometer at 600 nm wave length. The growth of bacterial isolates on maize pollen agar (Pollen in g/l: 10, 15, 20, 25, 30 and 35) was also observed by streak inoculation. Additionally, the optical density reading of bacterial isolates was also measured in nutrient broth medium for comparison. OD reading was taken for the blank made of only maize and nutrient broth at con-centration of 5l, 10l and 15 g/l used as a control.
Data analysis
All the data generated was analyzed using SPSS version 20. One way ANOVA was computed for analyzing the variation of microbial abundance and habitat water chemistry. The Pearson correlation analysis was used to analyze the correlation between physicochemical and biological parameters of the water samples collected from larva habitats. The total count of bacteria (CFU) were determined by standard formula (CFU = Number of colonies × DF of sample plated).
RESULTS
Sample site description
Sample sites were covered by vegetation including algae, grasses. Water condition was clear, turbid, flowing and/or standing. Some of the site exposed directly to sunlight, partially shaded or shaded.
The sites were borrow pit formed by people when they dug to extract mud for their house construction, puddles and the hoof prints on cattle way and human foot prints. They were divided in to three zones with regard to the availability of maize pollen source: Tasseled zone was the zone where male maize fluorescence remains attached and pollen was allowed to pollinate by natural process (wind); the detasseled zone was the site where the male maize fluorescence was removed and pollinated by hand and the buffer zone was where the pollen source was different from maize pollen such as taro, teff and green pepper.
Isolation of bacteria from water samples of larva habitats
Twenty different bacterial colonies were isolated from eighteen water samples of larva breeding habitats. The ten selected bacterial isolates for further studies in terms of their abundance was designated as AS6B, AS6C, AS11A, AS12B, AS15B, AS16D, AS17D, AS17E, AS19B and AS20 C. The colonies isolated were white, yellow, red, opaque, cream and orange colored. The white colonies were rough surface with leafy margin, the cream colonies were mucoid, and the other colonies were smooth with entire margin. The designation of isolates was the same as for yeast isolate designation.
Enumeration of bacteria from water samples of larva breeding habitats
Bacteria was found to be abundant in water from site number 6 (6.24±0.32) followed by site 7 (6.18±0.39) and site 12 (6.12±0.24) (Table 4). Site 14 contained the least number of bacteria (4.39±0.39) (Table 1).
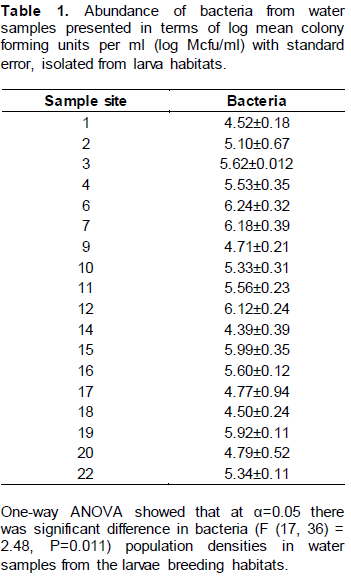
Evaluation of bacterial growth on maize pollen medium
Growth of bacterial isolates on maize pollen agar (PA)
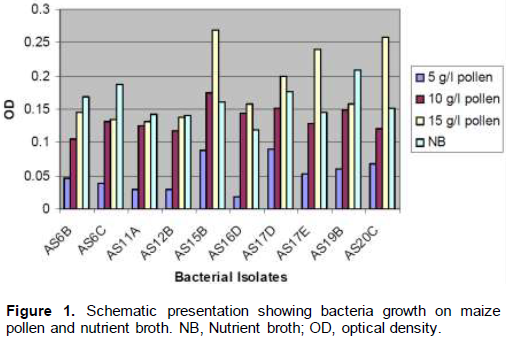
Among the ten most abundant bacteria selected from the isolates of water samples of the larva breeding sites, the bacterial isolate AS15B, AS17D, AS17E, AS19B and AS20C generally showed more growth on maize pollen agar than the other isolates (AS6B, AS6C, AS11A, AS12B and AS16D). As few as 5 g of maize pollen per liter was found to be enough to support the minimum growth of bacterial isolates.
Evaluation of bacterial isolates on maize pollen broth
The minimum growth of bacterial isolates was observed at 5 g/l of maize pollen (Figure 1). At concentration of 10 g/l of maize pollen, the bacteria showed appreciable growth. At 15 g/l of maize pollen concentration, the growth observed for isolates AS6B, AS6C, AS11A, AS12B and AS16D were almost similar but lower than the growth observed by the isolates AS15B, AS17D, AS17E, AS19B and AS20C. From the mean value of OD measured, isolates AS15B, AS17D, AS17E, AS19B and AS20 showed even better growth at 15 g/l of maize pollen concentration than in nutrient broth.
Measurement of nutrient contents of water samples from larva habitats (mean ± SEM)
The COD concentration measured from the water ample showed that it was not significantly different between the sample sites (p>0.05) at α=0.05. Sample site 10 showed the highest COD (247.67+103.22) concentration. Sample site 6 and 9 were the second in concentration of COD (200+83.34). Sample site 14 showed the least concentration of COD (11.0+4.58).
There was no significance difference between the amounts of ammonia nitrogen concentration measured from the water sample (P>0.05, α=0.05). The highest NH3-N was measured from water sample 9 (7.62±3.22) (Table 2) and the lowest was measured from water sample 16 (0.77±0.21). The highest NH4+ concentration was measured from water sample 9 (9.81±4.27) and the lowest from water sample (0.99±0.27).
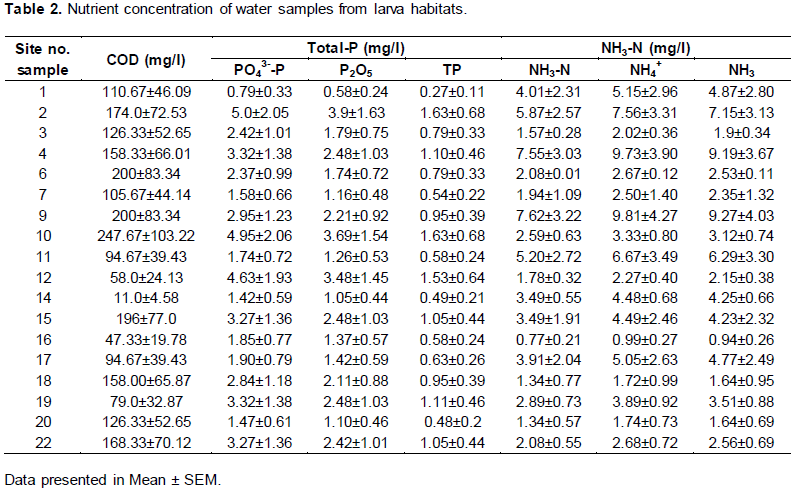
The concentration of reactive soluble phosphate- phosphorous (SRP-P) or PO43- was the highest in water sample 2 (5.0±2.05) and the lowest in water sample 1 (0.79±0.33). The concentration of Total phosphorous (TP) was measured the highest from water sample 2 and 10 (1.63±0.68) and the least from water sample 1 (0.27±0.11)
Correlation of biological and physicochemical parameters
Pearson’s correlations were calculated to test for relationships between bacterial abundance and environ-mental variables for bacterial communities in the larval breeding pool. The results indicated that bacterial abundance was positively but weakly correlated with COD (r2=0.096). The concentration of total phosphorous in a breeding site was positively but weakly correlated with the abundance of bacteria (r2=0.179). On the other hand the abundance of bacteria was negatively and weakly correlated with NH4+ (r2= -0.064) and NH3-N (r2= -0.063) (Table 3).
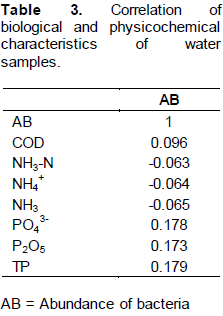
Identification of bacterial isolates
Based on the abundance in water samples ten bacterial isolates were selected for studies. The bacteria were identified to the genus level on the basis of their cultural and morphological (Table 3), biochemical (Table 4) and physiological characteristics.
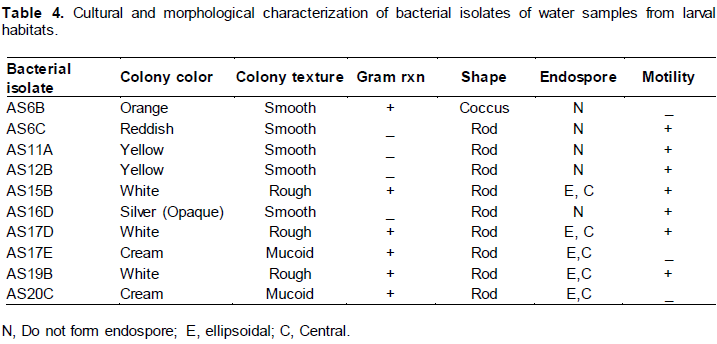
Cultural and morphological characterization of bacterial isolates
Ten bacterial isolates were selected in terms of their abundance in the water samples. The ten bacterial isolates produced white, cream, yellow, red, silver (opaque) and orange colony color on nutrient agar medium (Table 4). The colony surface was smooth, rough and mucoid (Table 4). The gram reaction and KOH test showed that 5 isolates were gram positive, rod shaped, 4 isolates were gram negative, rod shaped and one isolate was gram positive coccus. The endo spore stain showed that 5 isolates formed central and ellipsoidal endospore (Table 4).
Biochemical characterization of bacterial isolates
Seven isolates showed positive test for gelatin liquefaction and the remaining three showed negative result (Table 5). For starch hydrolysis test 4 isolates showed positive and six isolates showed negative result.
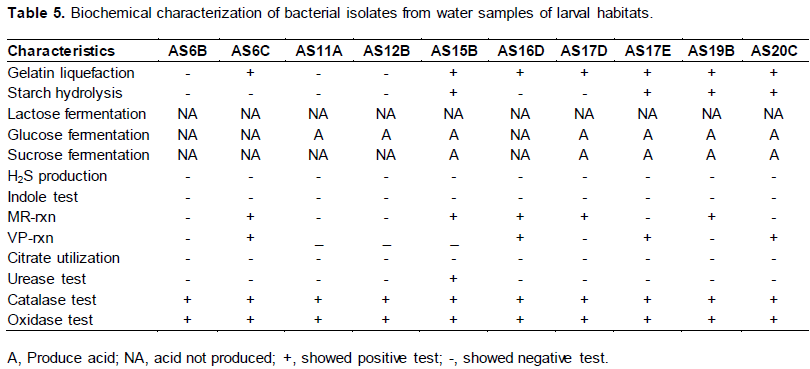
Carbohydrate fermentation test showed that none of the isolates fermented lactose, 7 isolates produced acid from glucose with no gas and 5 isolates produced acid from sucrose with no gas (Table 5). All the ten isolates neither produced H2S nor indole in SIM medium. Five isolates showed positive test result for MR and four isolate showed positive test result for VP test (Table 5). All isolates showed positive test for catalase and oxidase test. None of the isolates used citrate as the sole carbon source for growth. Only one isolate showed positive test result for urease test result. Seven isolate showed positive motility test result in SIM medium (Table 5).
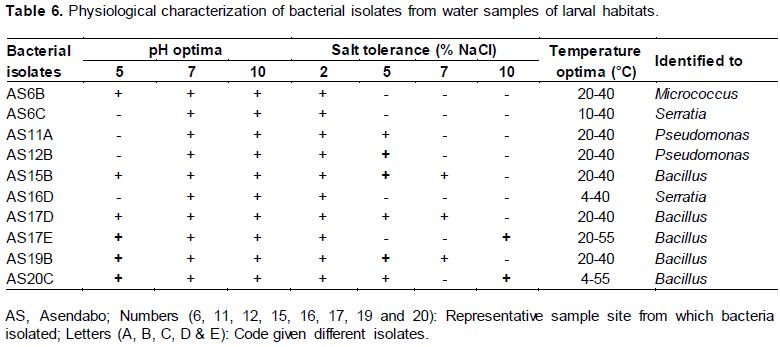
Physiological characterization of bacterial isolates
The optimum temperature for all bacterial isolates was between 20 and 40°C. Only two out of ten isolates rough and mucoid (Table 4). The gram reaction and KOH rough and mucoid (Table 4). The gram reaction and KOH showed growth at 4°C (AS16D and AS20C), 50 and 55°C (AS17E and AS20). All isolates were capable of growing at 2% NaCl in nutrient agar medium. None of the isolates showed growth at 12% NaCl concentration. Six isolates showed growth at 5% NaCl, three isolates grown at 7% NaCl and two isolates showed growth at 10% NaCl in nutrient agar solution. The optimum salt tolerance for the isolate was variable. The optimum pH for the isolate was 7 to 10 in which all isolates showed growth. Only three isolate showed growth at pH 5.
Eventually consulting the Bergey’s Manual of Systematic Bacteriology confirmed that the bacterial isolates were identified as Bacillus, Micrococcus, Pseudomonas and Serratia species (Table 6).
Anopheline larvae usually feed on microorganisms (Muturi et al., 2013) and detritus (Merritt et al., 1992). The microorganisms are able to thrive on maize pollen, which in turn serve as Anopheline larvae nutriment. The maize pollen contribute to the larval nutriment directly as the mosquito larvae use water soluble nutrient content of pollen (Yemane et al., 2003) and indirectly support the larval diet by allowing the proliferation of micro-organisms to comprise the microlayer of water bodies.
The population densities of bacteria are larger in tasseled zone than the detasseled and buffered zone. This is, may be, due to the nutritional enrichment of habitat of tasseled zone by maize pollen, which supplies additional nutrient to the microorganisms. Maize pollen dispersed about 60 m by wind and land on the larval breeding site nearby (Yemane et al., 2000). The bacteria in sites close to maize benefited more from the pollen, proliferate and become abundant.
The concentration of nutrient in the water sample of larva habitat drained and accumulated from surface runoff fertilized and irrigated soil was low. The concentration of ammonia nitrogen was lower and this might be the result of the transfer of volatile and semi volatile compounds (ammonia) from water bodies to the atmosphere (Schindler, 2001). The amount of ammonium concentration was low as compared to finding report by Kudom (2015). The small amount of ammonium concentration in the habitat of larvae suggests that larval excretion of NH4+was being balanced by its removal through microbial metabolism. However, without measurement of specific rates of ammonia oxidation, nitrification, and product consumption, it is difficult to assess the mechanism of larval impact on the process (Kaufman et al., 199). According to EEPA (2002), the standard for free ammonia is 5 g/l. In comparison to this EEPA (2002) standard, the concentrations of ammonia nitrogen is higher at sample site 2, 4 and 9 than the minimum standard and lower than the standard for the other study sites.
The concentration of SRP-P in the site was measured from 0.79 in site 1, which was the least concentration compared to 5.0 g/l in site 2 and 10 , which was the highest concentration measured in this study. This was smaller in comparison to the SRP-P measured from surface water (about 2-5 g/l) by Swift (1981) in marsh site, but higher from common types of A. gambiae breeding habitats (Imam and Deeni, 2014).
The relatively high levels of ammonia nitrogen relative to TP encountered in this microhabitat suggest phosphorous limitation (Kling et al., 2007). The smaller reactive soluble phosphate-phosphorous (RSP-P) in site 1 may have been the result of uptake by probably more abundant bacteria in the breeding pool. Because of its high reactivity, phosphorous may have lost due from the microhabitat by precipitation or adsorption on to inorganic particles which were continuously suspended as a result of mixing (Rediat, 2008).
The nutrient concentration of larval breeding habitat varies among the habitat; it can significantly influence the development and body size of emerging An. gambiae. Larval development duration and adult body size decrease but pupation rate in-creases when the organic content of the soil substrate increases. This supports the relation between larval habitat quality and mosquito response in terms of development time and body size of adult mosquitoes. The soil type of mosquito aquatic habitat is critical for larval development (Pfaehler et al., 2006). Maize pollen releases its nutrient content in to the larvae breeding site, in which the microorganism thrive on and facilitate the nutrient geochemical recycling, which create conducive microhabitat for larvae to flourish.
Furthermore Garros et al. (2008) suggested that algae and microorganisms are the main food source of A. quadrimaculatus larvae, and mosquito larvae are not discriminatory in the type of food they ingest. Analysis of the biodiversity present in the larval habitats was carried out and this study suggests that there is wide range of microorganisms available in natural habitats. This study can be used to improve our understanding of the larval ecology of African malaria vectors and to facilitate the development of new mosquito control strategy through bringing the changes on the larva habitats.
Even though there was no statistically significant difference between the concentrations of ammonia nitrogen in the breeding site, the sites differed in the abundance and diversity of microorganism. This study results showed that bacterial abundance in fresh waters can be stimulated by inorganic nutrients (Le et al., 1994).This might be due to the supplementary nutriment of bacteria by the maize pollen, which had provided soluble nutritional material for microbes. Nutrient released by the decomposition of maize pollen is equally available and important for proliferation of bacteria and other microorganism as well. The water sample nutrient concentration in larval habitats of anopheline probably has crucial roles in the resource ecology of these mosquitoes (Mala and Irungu, 2011).
There were more bacteria in water sampling sites with pollen access (sample site 6, 7, 11, 12 and 15) than sites of low pollen access. This confirmed that maize pollen is an important nutrient to bacteria in a natural setting. Therefore, it requests additional investigation on using maize cultivar, which could with stand drought and produce crop in a dry season than during rainy season. This will minimize the availability of maize pollen to microorganism as well as to mosquito larvae and hence, results in vector control and reduction of malaria transmission.
The accumulations of organic matter under the surface film of water bodies form the surface micro-layers. Heterotrophic microorganisms benefited from this organic matter for growth, and they, in turn, are fed upon by Anopheles mosquito larvae. From laboratory experiments by Wotton et al., 1997 mosquito larvae, An. gambiae and An. quadrimaculatus, grow most rapidly where surface microlayers are present and, especially, where labile dissolved organic matter (may be contributed by the maize pollen shedding in our case) is added to promote growth of microorganisms, which are the components of the larval diet that most affect growth (Wotton et al., 1997).
CONCLUSION
The results of the present study have revealed that the chemical parameter of water samples from larva habitats of Anopheles mosquito is correlated to the abundance of microorganisms. The microbial flora of the water sample was dominated by the Bacteria species, Bacillus, Micrococcus, Pseudomonas and Serratia.
The abundance of bacteria in the breeding site is seen to be largely a function of maize pollen concentration. Bacteria, fungi and yeasts are the microflora which constitutes the feeding surface microlayer of Anopheline larva habitat in the natural habitat, from which the larva obtain their feed. The larvae in natural habitat feed indiscriminately on particulate organic matter and microorganisms and hence, the bacteria isolated and identified from the water samples of larva breeding habitats could also be good nutrient source for larva. The microorganism of larva breeding habitat, close to maize pollen source obtain more nutrient from this nutritious plant in addition to nutrient from substratum, become more abundant and enrich the water surface microlayer with microbial flora.
RECOMMENDATION
Based on this study the need to verify the types and numbers of microorganisms in the gut of Anopheles mosquito larvae in relation to their habitats (water bodies) is necessary.
CONFLICTS OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Abdulrahaman A, Kolawole AOM (2006). Traditional preparations and uses of maize in Nigeria. Ethnobotanical Leaflets 10:219-227. |
|
|
Ayele AA, Gebre-Michael T, Balkew M, Lindtjørn B (2012). Abundance and dynamics of anopheline larvae in a highland malarious area of south-central Ethiopia. Parasites & Vectors 5(117):1-9 |
|
|
Adugna N, Melaku A (2001). Hybrid maize seed production and commercialization: The experience of pioneer hybrid seeds in Ethiopia. Second national maize workshop of Ethiopia. Pioneer Hybrid Seeds PLC, Addis Ababa. |
|
|
Afrane A, Lawson BW, Brenya R, Kruppa T, Yan G (2012). The ecology of mosquitoes in an irrigated vegetable farm in Kumasi, Ghana: abundance, productivity and survivorship. Parasit. Vectors 5(233):1-7. |
|
|
Aneja KR (2005). Experiments in Microbiolo-gy, Plantpathology and Biotechnology, 4th edn. New Age International PLtd. Publishers, New Delhi pp. 245-275. |
|
|
APHA (1998). Standard Methods for the Examination of Water and Waste Water, 20th edn. American Public Health Association, Wshington, DC. |
|
|
Barros FS, Arruda ME, Gurgel HC, Honório NA (2011). Spatial clustering and longitudinal variation of Anopheles darlingi (Diptera:Culicidae) larvae in a river of the Amazon:the importance of the forest fringe and of obstructions to flow in frontier malaria. Cambridge University Press, Bullet. Entomol. Res. 101:643-658. |
|
|
Bergey's Manual of Systematic Bacteriology (1984). Eds. Krieg, N.R. and Holt, J.G. Williams and Wilkins, Baltimore, USA. Vol 1. |
|
|
Berhanu G, Fernandez-Rivera S, Mo-hammed H, Mwangi W, Seid A (2007). Maize and livestock: their inter-linked roles in meeting human needs in Ethiopia, Pp.1-87. Research report 6 .ILRI, Nirobi. |
|
|
Bezawit E (2007). Nutrient Composition of Maize Pollen and its Microbial Degradation. M. Sc. Thesis, Addis Ababa University, Addis Ababa. |
|
|
Bisen PS, Verma K (1994). Hand Book of Microbiology. CBS Publishers, New Delhi, pp. 47-65. |
|
|
Chester B (1979). Semi quantitative catalase test as an aid in identification of oxidative and non saccharolytic gramnegative bacteria. J. Clin. Microbiol. 10:525-528. |
|
|
EEPA (2002). General standards for all other industrial effluents: effluent discharges to inland water. Ethiopian Environment Protection Authority, Addis Ababa, pp. 20-23. |
|
|
Fry JC, Zia T (1982). Viability of heterotrophic bacteria in freshwater. J. General Microbiol. 128:2841-2850. |
|
|
Garros C, Ngugi N, Githeko AE, Tuno N, Yang G (2008). Gut content identification of larvae of the Anopheles gambiae complex in Western Kenya using a barcoding approach. Mol. Ecol. Res. 8:512-518. |
|
|
Getahun D, Mwangi W, Verkuijl H, Abdishekur W (2000). An assessment of the adoption of seed and fertilizer packages and the role of credit in smallholder maize production in Sidama and North Omo Zone, Ethiopia. Mexico, D.F.: International Maize and Wheat Improvement Center (CIMMYT) and Ethiopian Agricultural Research Organization (EARO), pp. 12-22. |
|
|
Herrel N, Amerasinghe FP, Ensink J, Mukhtar M, Vanderhoek W, Konradsen F (2001). Breeding of anopheles mosquitoes in irrigated areas of South Punjab, Pakistan. Blackwell Science Ltd, Med. Veterinary Entomol. 15:236-248. |
|
|
Imam AA, Deeni Y (2014). Common Types of Anopheles gambiae Breeding Habitats in North Western Nigeria. J. Innov. Res. Eng. Sci. 4(4):496-504. |
|
|
Kaufman MG, Walker ED, Smith TW, Merritt RW, Klug MJ (1999). Effects of larval mosquitoes (Aedes triseriatus) and stem flow on microbial community dynamics in container habitats. Appl. Environ. Microbiol. 65:2661-2673. |
|
|
Kettle DS (1995). Medical and Veterinary Entomology, 2nd edn. CABI Publisher pp. 109-151. |
|
|
Kling LJ, Juliano SA, Yee DA (2007). Larval mosquito communities in discarded vehicle tires in a forested and unforested site: detritus type, amount, and water nutrient differences. J. Vector Ecol. 32:207-217. |
|
|
Kudom AA (2015). Larval ecology of Anopheles coluzzii in Cape Coast, Ghana: water quality, nature of habitat and implication for larval control. Malaria J. 14(447):1-13. |
|
|
Le J, Wehr JD Campbell L (1994). Uncoupling of bacterioplankton and phytoplankton production in fresh Waters is affected by inorganic nutrient limitation. Appl. Environ. Microbiol. 60:2086-2093. |
|
|
Machault V, Gadiaga L, Vignolles C, Jarja-val F, Bouzid S, Sokhna C, Lacaux, J, Trape J, Rogier C Pages F (2009). Highly focused Anopheline breeding sites and malaria transmission in Dakar. Malaria J. 8:138. |
|
|
Maekawa E, Aonuma H, Nelson B, Yoshimura A, Tokunaga F, Fukumoto S, Kanuka H (2011). The role of proboscis of the malaria vector mosquito Anopheles stephensis in host seeking behavior. Parasites Vectors 4(1):1-10. |
|
|
Mala AO, Irungu LW (2011). Factors influencing differential larval habitat productivity of Anopheles gambiae complex mosquitoes in a western Kenyan village. J. Vector Borne Dis. 48:52-57. |
|
|
Merritt RW, Dadd RH, Walker ED (1992). Feeding behavior, natural food, and nutritional relationships of larval mosquitoes. Ann. Rev. Entomol. 37:349-376. |
|
|
Mondal R, Devi NP, Jauhari RK (2015). Bacterial characterization in natural breeding habitats of Aedes mosquitoes and their role on ovipositional response. Int. J. Mosq. Res. 2(3):175-181. |
|
|
Muirhead-Thomson RC (1958). The ecology of vector snail habitats and mosquito breeding- places: The experimental approach to basic problems. Bull. org. Mond. Sante 19:637-659. |
|
|
Muturi EJ, Orindi BO, Kim CH (2013). Effect of Leaf Type and Pesticide Exposure on Abundance of Bacterial Taxa in Mosquito Larval Habitats. PLoS/ONE 8(8):1-8. |
|
|
Mwangangi JM, Mbogo CM, Muturi EJ, Nzovu JG, Githure JI, Yan G, Minakawa N, Novak R, Beier JC (2007). Spatial distribution and habitat characterization of Anopheles larvae along the Kenyan coast. J. Vect. Borne Dis. 44:44-51. |
|
|
Okech BA, Gouagna LC, Yan G, Githure JI, Beier JC (2007). Larval habitats of Anopheles gambiae s.s. (Diptera: Culicidae) influences vector competence to plasmodium falciparum parasites. Malaria J. 6:50. |
|
|
Okogun GRA, Nwoke BEB, Okere ANJ, Anosike JC, Esekhegbe AC (2003). Epidemiological implications of preferences of breeding sites of mosquito species in mid-western Nigeria. Ann. Agric. Environ. Med. 10:217-222. |
|
|
Pereira EDS, Sarquis MID, Ferriera-Keppler RL, Hamada N, Alencar Y (2009). Filamentous fungi associated with mosquito larvae (Dipteria: Culicidae) in municipalities of the Brazilian Amazon. Neotrop. Entomol. 38:352-359. |
|
|
Pfaehler O, Oulo DO, Guoagna LC, Githure J, Guerin PM (2006). Influence of soil quality in the larval habitat on development of Anopheles gambie giles. J. Vector Ecol. 31:400-405. |
|
|
Phillips S (1995). Poaceae (Gramineae). In: Flora of Ethiopia and Eritrea, 7:365-368 (Hedberg, I. and Edwards S, eds). Addis Ababa and Uppsala. |
|
|
Rediat A (2008). Seasonal Studies on Phyto-plankton in Relation to some Biological and Physicochemical Factors in lake HoraKilole, Ethiopia. M. Sc. Thesis, Addis Ababa University, Addis Ababa. |
|
|
Schindler D (2001). The cumulative effects of climate warming and other human stresses on Canadian freshwaters in the new millennium. Can. J. Fish. Aquat. Sci. 58:18-29. |
|
|
Swift DR (1981). Preliminary investigations of periphyton and water quality relation-ships in the everglades water conservation areas. South Florida Water Management District Technical Publication 81-5. West Palm Beach, Florida, 83 p. |
|
|
Tchioffo MT, Boissie`re A, Churcher TS, Abate L, Gimonneau G, Nsango SE, Awono-Ambe’ne´ PH, Christen R, Berry A, Morlais I (2013). Modulation of Malaria Infection in Anopheles gambiae Mosquitoes Exposed to Natural Midgut Bacteria. PLoS ONE 8(12):1-9. |
|
|
Wotton RS, Chaloner DT, Yardley TCA, Merrittt RW (1997). Growth of Anopheles mosquito larvae on dietary microbiota in aquatic surface microlayers. Med. Vet. Entomol. 11:65-70. |
|
|
Yemane Ye-ebiyo, Pollack RJ, Spielman A (2000). Enhanced development in nature of larval Anopheles arabiensis mosquitoes feeding on maize pollen. Am.J.Med.Hyg.63: 90-93. |
|
|
Yemane Ye-ebiyo, Pollack RJ, Kiszewski A, Spielman A (2003a). Enhancement of development of larval Anopheles arabiensis by proximity to flowering maize (Zea mays) in turbid water and when crowded. Am. J. Trop. Med. Hyg. 68:748-752. |
|
|
Yemane Ye-ebiyo, Pollack RJ, Kiszewski A, Spielman A (2003b). A component of maize pollen that stimulates larval mosquitoes (Dipteria: Culicidae) to feed and increases toxicity of microbial larvicides. J. Med. Entomol. 40:860-864. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


