Review

ABSTRACT
Biological control by antagonistic organisms is a potential nonchemical tool for crop protection against phytopathogenic fungi. Trichoderma spp. are the most commonly used biocontrol agent that are being used against many soil borne pathogens. The biocontrol mechanism of Trichoderma involves mycoparasitism, antibiosis and competition for key nutrients. Chitinase, xylanase and β-glucanase are the major cell wall-degrading enzymes (CWDEs) that are involved in the mycoparasitic action. CWDEs improve the biocontrol activity of these strains. Different carbon sources are supplemented in the growth medium for the induction of these CWDEs.
Key words: Cell wall-degrading enzymes (CWDEs), mycoparasitic, phytopathogenic, Trichoderma.
INTRODUCTION
Traditional methods of crop protection include use of chemical pesticides. But the use of chemical pesticides has drastic effects on environment and consumers. Chemical pesticides are not beneficial for long term use as they pollute the environment, leave harmful residues in the soil and they also lead to the development of resistant strains. Use of biocontrol agents (BCAs) is the solution for this problem. The term biocontrol agent includes use of benifical microbes and their products against pathogens. In present era there are number of BCAs that are available and commercialized in agriculture field against plant pathogens. To date, a number of BCAs are available as commercial products, as Agrobacterium, Pseudomonas, Streptomyces and Bacillus, and fungal genera such as Gliocladium, Trichoderma, Ampelomyces, Candida and Coniothyrium. Trichoderma spp. are the most common soil fungi (Harman et al., 2004). These fungi are opportunistic, avirulent plant symbionts, and act as parasites and antagonists of many phytopathogenic fungi, such as Sclerotonia, Pythium etc. Thus, protecting plants from disease. Trichoderma spp. are among the most studied fungal BCAs and commercially marketed as biopesticides, biofertilizers and soil amendments (Harman, 2000; Harman et al., 2004; Lorito et al., 2004). Trichoderma provides numerous advantages: (i) colonization of the rhizosphere by the BCA (‘‘rhizosphere competence’’) allowing rapid establishment within the stable microbial communities in the rhizosphere; (ii) control of pathogenic and competitive/deleterious micro-flora by using a variety of mechanisms; (iii) improvement of the plant health and (iv) stimulation of root growth (Harman et al., 2004; Papavizas et al., 1985).
Several strains from the genus Trichoderma have been described as antagonistic fungi able to control a wide range of phytopathogenic fungi. The antifungal activity of Trichoderma involves production of antibiotics, including competition for key nutrients, and production of fungal cell wall-degrading enzymes (CWDEs) (Hjeljord and Tronsmo, 1998; Elad et al., 1982).
MYCOPARASITIC ACTION AND LYTIC ENZYMES
Process of mycoparasitic action includes several steps: Recognition of the host, attack and subsequent penetration and killing. During this process Trichoderma secretes CWDEs that hydrolyze the cell wall of the host fungus, subsequently releasing the cell wall oligomers (Kubicek et al., 2001; Howell, 2003; Woo et al., 2006).
The antifungal action of Trichoderma spp. includes a great variety of lytic enzymes such as proteases, xylanase, chitinase, glucanases (Lorito, 1998; Lorito et al., 1994a, 1996a), most of which play a great role in biocontrol (Harman and Kubicek, 1998; Baek et al., 1999; Carsolio et al., 1999; Woo et al., 1999; Zeilinger et al., 1999; Kullnig et al., 2000; Kubicek et al., 2001). These CWDEs isolated from the different strains of Trichoderma have been purified and characterized (Lorito, 1998). Interestingly, when tested alone or in combinations, the purified proteins showed antifungal activity against a variety of pathogens (that is species of Rhizoctonia, Fusarium, Alternaria, Ustilago, Venturia and Colletotrichum, as well as fungus like organisms such as the Oomycetes Pythium and Phytophthora which lack chitin in their cell walls (Tronsmo, 1991; Lorito et al., 1993, 1994a).
The direct application of these secreted CWDEs produced by fungal BCAs, instead of using the whole ‘‘live’’ organisms, has numerous advantages in industry and agriculture. The selective production of these enzymes active can be increased by modifying the growth conditions, that is type and composition of culture medium, carbon sources, temperature of incubation and pH, etc. (Lorito and Scala, 1999; Lorito et al., 2006). The presence of different carbon sources, such as mono- or polysaccharides, colloidal chitin, or fungal tissues, has been shown to increase the secretion of CWDEs (Mach et al., 1999). Moreover, purified mixture of CWDEs showed improved antifungal effects against various plant pathogens (Lorito et al., 1994b, 1996a; Baek et al., 1999; Carsolio et al., 1999).
Xylanases
Xylanases (β-1,4-xylan xylanohydrolase, E.C.3.2.1.8) (Figure 1) are the major components of xylanolytic enzymes and randomly cleave the β-1,4-glycosidic bond of xylan backbone. Xylan is the second most abundant natural polysaccharide. Xylan is a heteropolysaccharide consisting of β -1,4-linked D-xylose monomers in connection with side branches of arabinosyl, glucuronosyl, acetyl, uronyl, and mannosyl residues (Tsujibo et al., 1997). Complete degradation of xylan structures requires the concerted and synergistic function of several enzymes including endo-β-1,4-xylanases (EC 3.2.1.8) (Jun et al., 2009). Due to the broad applications in biopulping and biobleaching in paper industry, xylanase has attracted the attentation of many researchers (Tsujibo et al., 1997).
.png)
Xylanase production
Solid state fermentation (SSF) was carried out at 30°C with 50 ml of Vogel?s medium with some modifications and 1% of birchwood xylan in 250 ml Erlenmeyer flasks in a BOD incubator. After eight days, the contents of the flasks were filtered through filter paper and the obtained culture broths were centrifuged 10 min at 3000 × g supernatants were then assayed for extracellular xylanase activity.
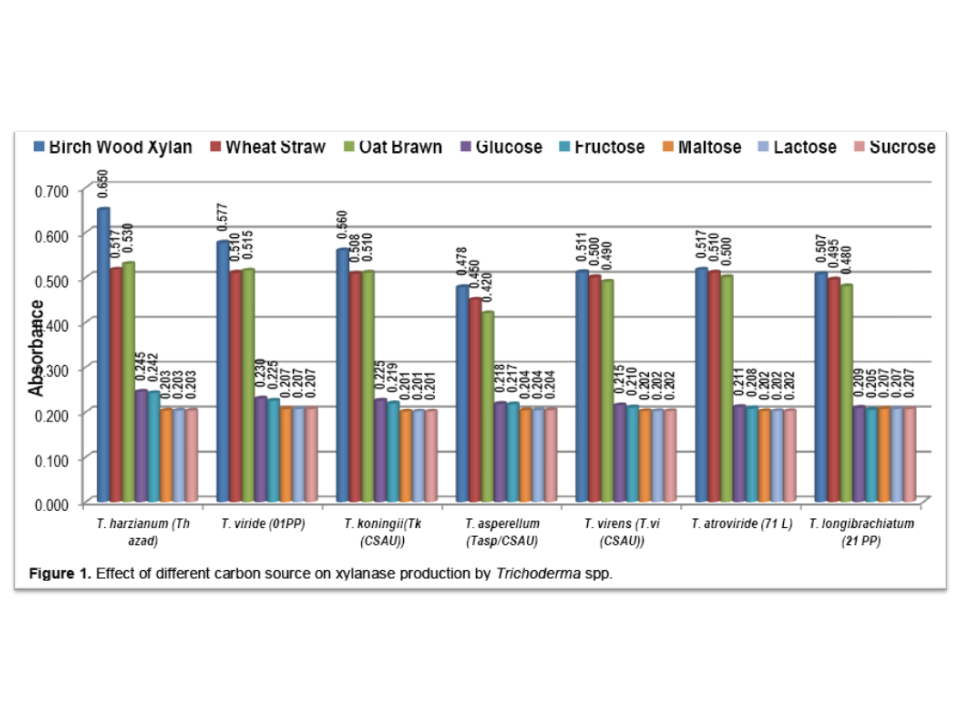
For maximum xylanase production of, various culture conditions viz., carbon and nitrogen source, inoculum size, pH, temperature and incubation period were optimized by conventional “one variable at a time” approach which involves varying a single independent variable at a time while maintaining the others at a constant level. Fungal isolates were grown in different media containing 1% birchwood xylan, at pH 6.0 for 192 h. Medium giving maximum growth was further used to standardize the inoculum size by inoculating different concentrations (1-5 ml) of inoculums. Similarly, the temperature and time of incubation were optimized by growing the fungal isolate at different temperatures (25-65°C) for different time periods (96-192 h). Different organic viz., yeast extract (YE), beef extract (BE), peptone (PPT), soybean residue (SR) and corn powder (CP) were supplemented separately to a final con-centration of 0.3% (w/v) to study the microbial growth and xylanase activity. Glucose, maltose and sucrose at a final concentration of 0.1% and wheat bran (WB), corn cob (CC) birchwood xylan (BW) and carboxy methyl cellulose (CMC) were used as the carbon source at a final concentration of 1.0% (Figure 2). After 7 days of growth, the fungal broth was filtered through filter paper and centrifuged at 3000 × g for 15 min at 4°C in a refrigerated centrifuge. The supernatant taken as enzyme extract containing extracellular xylanase was used to assay the enzyme activity. The highest xylanase activity was shown by Birch wood xylan (Sonika et al., 2014).
Applications of xylanases
Besides the use of xylanase enzymes in antagonistic mechanism, the potential applications of xylanases also include the bioconversion of lignocellulosic material and agro-wastes into fermentative products, the clarification of juices, the improvement of the consistency of beer and the digestibility of animal feed stocks. The incorporation of xylanase from Trichoderma longibrachiatum into the rye-based diet of broiler chickens reduced intestinal viscosity, thus, improving both the weight gain of the chicks and their feed conversion efficiency. One of the most important biotechnological applications of xylanase is its use in pulp bleaching.
Glucanase enzyme
β-1,3-glucanases are hydrolytic enzymes, widely distributed among bacteria, fungi and higher plants. β-1,3(1,6)-glucans are major components in cell wall of yeasts and fungi. In most yeast, especially Saccharomyces. Trichoderma spp. secretes β-1,3-glucanases when it is grown on polysaccharides, fungal cell walls.
Cellulose is the most abundant organic polymer on earth. Cellulases hydrolyze β-1,4- glycosidic bond in the cellulose structure and operate as a multicomponent enzyme system which consists of three classes of enzyme; Endoglucanases (endo-β-1,4-glucanase, CMCase, EC 3.2.1.4), cellobiohydrolases (exoglucanase, avicelase, EC 3.2.1.91) and β-glucosidase (cellobiase, EC 3.2.1.21) (Dincer and Telefoncu, 2006). The first two enzymes are called as “real cellulose” and produce cellobiose and glucose by acting directly on cellulose. The cellobiose is then hydrolyzed into glucose by β-glucosidase (Dincer and Telefoncu, 2006; Andersen, 2007). Endoglucanases cleave the internal glycosidic bonds of cellulose chains and act synergistically with exoglucanase and β-glucosidase during the hyrolysis of crystalline cellulose. Also, endoglucanases are used in increasing the yield of fruit juices, beer filtration, and oil extraction, improving the nutritive quality of bakery products and animal feed, and enhancing the brightness, smoothness, and over all quality of cellulosic garments. Therefore, new fungal sources and properties (optimal pH and temperature) of EG must be investigated (Naika et al., 2007).
Glucanase enzyme production
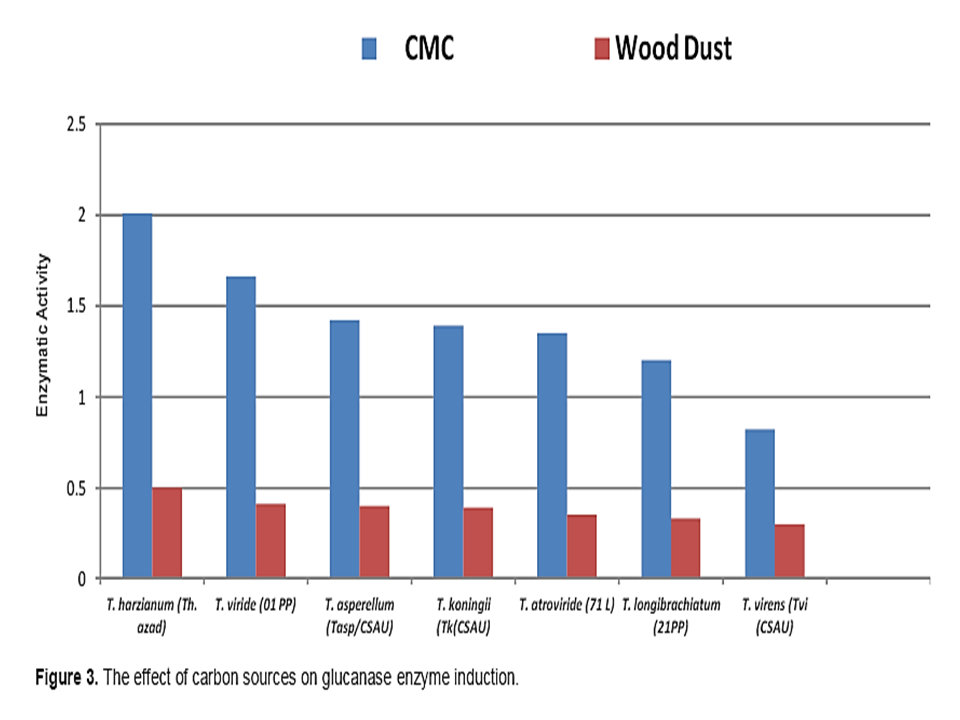
Trichoderma sp. previously isolated from the different states of Uttar Pradesh were cultivated on Czapek Dox Medium containing CMC and wood dust as sole carbon source (1%) (Figure 3). Cultures were incubated for 10-14 days on orbital shaker at 150 rpm. At the end of the incubation time contents of conical flasks were filtered and the filtrate was centrifuged at 5000 rpm for 10 min. The clear supernatant was considered as the source of crude enzymes. The clear supernatant used as a source of crude enzyme was purified by the slow addition of Ammonium Persulfate with continuous stirring till 80% saturation. The obtained precipitate was dissolved in citrate phosphate buffer pH 5.0 and used for enzyme activity determination (Pandey et al., 2014).
Chitinase enzyme
Trichoderma are well known producer of chitinolytic enzymes and used commercially as a source of these proteins. Due to the importance of chitinolytic enzymes in insect, nematode, and fungal growth and development, they are receiving attention in regard to their deve-lopment as biopesticides or chemical defense proteins in transgenic plants and microbial biocontrol agents. In this sense, biological control of some soil-borne fungal diseases has been correlated with chitinase production. Fungi and bacteria producing chitinases exhibit antagonism against fungi, and inhibition of fungal growth by plant chitinases has been demonstrated. Insect pathogenic fungi have considerable potential for the biological control of insect pests.
Chitin, a homopolymer of β-(1, 4)-N-acetylglucosamine (GlcNAc) is the second most abundant source of nutrients and energy after cellulose (Agrawal and Kotasthane, 2012; Lorito et al., 2004; Tronsmo, 1991; Dincer and Telefoncu, 2006). It is widely distributed in the nature as the integuments of insects and crustaceans and as a component of fungi and algae (Baek et al., 1999; Hjeljord and Tronsmo, 1998). Chitinases are a group of enzymes that decompose chitin into a variety of products that include the deacylated oligomer chitosan, the disaccharide chitobiose and the monomer N-acetyl glucosamine (Dincer and Telefoncu, 2006; Pandey et al., 2014). The present study was aimed for screening the different Trichoderma species to select potent isolate, production and purification of chitinase followed by determination of its molecular weight (Saraswathi and Jaya, 2014).
For the evaluation of chitinases activity, two different insoluble chitin sources (colloidal chitin derived from commercial chitin and Seashells) were used. Chitinase detection medium consisted of a basal medium comprising (per liter) 0.3 g of MgSO4.7H2O, 3.0 g of (NH4)2SO4, 2.0 g of KH2PO4, 1.0 g of citric acid monohydrate, 15 g of agar, 200 μl of Tween-80, 4.5 g of chitin source and 0.15 g of bromocresol purple; pH used was adjusted to 4.7. Lukewarm medium was poured in Petri plates and allowed to solidify. Fresh culture plugs of the isolates to be tested for chitinase activity were inoculated on the medium and incubated at 25±2°C and were observed for colored zone formation (Agrawal and Kotasthane, 2012). In our study, it was found that during the study the colloidal chitin derived from the commercial chitin was more potent in inducing chitinase enzyme activity as compared to the colloidal chitin derived from the sea- shells.
CONFLICT OF INTERESTS
The authors have declared that they have no conflict of interests.
ACKNOWLEDGEMENT
The authors are grateful for the financial support granted by the ICAR under the Niche Area of Excellence on “Exploration and Exploitation of Trichoderma as an antagonist against soil borne pathogens” running in Biocontrol Laboratory, Department of Plant Pathology, Chandra Shekhar Azad University of Agriculture and Technology, Kanpur, India.
REFERENCES
|
Agrawal T, Kotasthane AS (2012) Chitinolytic assay of indigenous Trichoderma isolates collected from different geographical locations of Chhattisgarh in Central India SpringerPlus. 1:73. |
|
|
Andersen N (2007). Enzymatic hydrolysis of cellulose. Ph.D. dissertation, Dept. Chemical Eng., Technical University of Denmark, Copenhagen, Denmark. |
|
|
Baek JM, Howell CR, Kenerley CM (1999). The role of an extracellular chitinase from Trichoderma virens Gv29-8 in the biocontrol of Rhizoctonia solani. Curr. Genet. 35:41-50. |
|
|
Carsolio C, Benhamou N, Haran S, Cortes C, Gutierrez A, Chet I, Herrera-Estrella A, (1999). Role of the Trichoderma harzianum endochitinase gene, ech42, in mycoparasitism. Appl. Environ. Microbiol. 65:929-935. |
|
|
Dincer A, Telefoncu A (2006). Improving the stability of cellulase by immobilization on modified polyvinyl alcohol coated chitosan beads. J. Mol. Catal. B Enzym. 45 (5): 10-14, |
|
|
Elad Y, Chet I, Henis Y (1982). Degradation of plant pathogenic fungi by Trichoderma harzianum. Can. J. Microbiol. 28:719-725. |
|
|
Harman GE (2000). Myths and dogmas of biocontrol: changes in perceptions derived from research on Trichoderma harzianum T-22. Plant Dis. 84:377-393. |
|
|
Harman GE, Howell CR, Viterbo A, Chet I, Lorito M (2004).Trichoderma species-opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2: 43-56. |
|
|
Harman GE, Kubicek CP (1998). Trichoderma and Gliocladium. Taylor & Francis, London, 278p. |
|
|
Hjeljord L, Tronsmo A (1998). Trichoderma and Gliocladium in biological control: an overview. in Trichoderma and Gliocladium: enzymes, biological control and commercial applications, eds Harman G. E., KubicekC. (Taylor and Francis, Ltd. London, United Kingdom) 2:131-151. |
|
|
Kubicek CP, Mach RL, Peterbauer CK, Lorito M (2001). Trichoderma: from genes to biocontrol. J. Plant Pathol. 83:11-23. |
|
|
Kullnig C, Mach RL, Lorito M, Kubicek CP (2000). Enzyme diffusion from Trichoderma atroviride (T. harzianum P1) to Rhizoctonia solani is a prerequisite for triggering of Trichoderma ech42 gene expression before mycoparasitic contact. Appl. Environ. Microbiol. 66:2232-2234. |
|
|
Lorito M, (1998). Chitinolytic enzymes and their genes. Taylor and Francis Ltd., London. pp. 2:73-99. |
|
|
Lorito M, Harman G.E, Hayes C.K, Broadway RM, Tronsmo A, Woo SL, Di Pietro A (1993). Chitinolytic enzymes produced by Trichoderma harzianum: antifungal activity of purified endochitinase and chitobiosidase. Phytopathology 83:302-307. |
|
|
Lorito M, Hayes, CK Di Pietro, A, Woo SL, Harman GE (1994a). Purification, characterization and synergistic activity of a glucan 1,3-bglucosidase and an N-acetyl-b-glucosaminidase from Trichoderma harzianum. Phytopathology 84:398-405. |
|
|
Lorito M, Woo SL, D'Ambrosio, M., Harman, GE, Hayes CK, Kubicek CP, Scala F, (1996a). Synergistic interaction between cell wall degrading enzymes and membrane affecting compounds. Molecular Plant-Microbe Interaction. 9: 206-213. |
|
|
Lorito M, Woo SL, Scala F, (2004). Le biotecnologie utili alla difesa sostenibile delle piante: i funghi. Agroindustria 3: 181-195. |
|
|
Lorito M, Woo SL, D'Ambrosio M, Harman GE, Hayes CK, Kubicek CP, Scala F (1996a). Synergistic interaction between cell wall degrading enzymes and membrane affecting compounds. Mol. Plant Microbe Interact. 9: 206-213. |
|
|
Lorito M, Woo SL, Scala F (2004). Le biotecnologie utili alla difesa sostenibile delle piante: i funghi. Agroindustria 3:181-195. |
|
|
Lorito M, Woo SL, Iaccarino M, Scala F (2006). Microrganismi antagonisti. In: Iaccarino, M. (Ed.), Microrganismi Benefici per le Piante. Idelson-Gnocchi s.r.l., Napoli, Italia, pp. 146-175. |
|
|
Mach RL, Peterbauer CK, Payer K, Jaksits S, Woo SL, Zeilinger S, Kullnig CM, Lorito M, Kubicek CP (1999). Expression of two major chitinase genes of Trichoderma atroviride (T. harzianum P1) is triggered by different regulatory signals. Appl. Environ. Microbiol. 65: 1858-1863. |
|
|
Naika GS, Kaul P, Prakash V (2007). Purification and characterization of a new endoglucanase from Aspergillus aculeatus. J. Agric. Food Chem. 55:7566-7572. |
|
|
Pandey S, Shahid M, Srivastava M, SharmaA, Singh A Kumar V, Srivastava Y (2014). Isolation and Optimized Production of Xylanase under Solid State Fermentation Condition from Trichoderma sp. Int. J. Adv. Res. 2 (3): 263-273. |
|
|
Pandey S, Shahid M, Srivastava M, Sharma A, Singh A, Kumar V (2014). Isolation, Purification and Characterization of Glucanase Enzyme from the Antagonistic Fungus Trichoderma. Int. J. Sci. Eng. Res. 5 (3): 2229-5518. |
|
|
Papavizas GC (1985). Trichoderma and Gliocladium: biology, ecology and potential for biocontrol. Annu. Rev. Phytopathol. 23:23-54. |
|
|
Saraswathi M, Jaya MR (2013). Production and purification of chitinase by trichoderma harzianum for control of sclerotium rolfsii international J. Appl. Nat. Sci. 2 (5): 65-72. |
|
|
Tronsmo A (1991). Biological and integrated controls of Botrytis cinerea on apple with Trichoderma harzianum. Biol. Control 1: 59-62. |
|
|
Tsujibo H, Ohtsuki T, Iio T, Yamazaki I, Miyamoto K, Sugiyama M, Inamori Y (1997). Cloning and sequence analysis of genes encoding xylanases and acetyl xylan esterase from Streptomyces thermomviolaceus OPC-520. Appl. Environ. Microbiol. 63: 661-664. |
|
|
Woo SL, Donzelli, B, Scala, F Mach R, Harman GE, Kubicek CP, Del Sorbo G, Lorito M (1999). Disruption of the ech42 (endochitinaseencoding) gene affect biocontrol activity in Trichoderma harzianum P1. Mol. Plant Microbe Interact. 12: 419-429. |
|
|
Woo SL, Scala F, Ruocco M, Lorito M (2006). The molecular biology of the interactions between Trichoderma spp., phytopathogenic fungi, and plants. Phytopathology 96:181-185. |
|
|
Zeilinger S, Galhaup C, Payer K, Woo SL, Mach RL, Fekete C, Lorito M, Kubicek CP (1999). Chitinase gene expression during mycoparasitic interaction of Trichoderma harzianum with its host. Fungal Genet. Biol. 26: 131-140. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


