ABSTRACT
The Magadi Lake located in the Southern Rift Valley in Kenya is a hypersaline and alkaline lake. The lake’s pH ranges from pH 9 to 11 whereas the salinity can reach saturation during the dry Season. Little studies have previously been done on Lake Magadi as compared to other soda lakes from Kenya. This study concentrated on isolating and characterizing haloalkaliphilic bacteria found in the hot springs of Lake Magadi. Collection of water, wet sediments and microbial mats was carried out from the springs in the main lake at a temperature of 45.1°C and from Little Magadi Nasikie Engida (temperature of 81 and 83.6°C). Seven bacterial isolates were identified using various media prepared with filter-sterilized water from the lake. These isolates were characterized and analyzed for production of extracellular enzymes and antimicrobial compounds. Bacteria recovered, thrived at pH ranging from 6 to 14, a temperature range of 30 to 60°C and sodium chloride range of 0 to 17.5%. The isolates produced amylases, lipases, proteases, cellulases, xylanases, esterases and also exhibited a range of inhibitory effects on various test organisms. Sequencing of the 16S rDNA of the isolates was done and sequences for 7 isolates were without ambiguities. BLAST results revealed that 90% of isolated microbes had an affiliation to the genus Bacillus while 10% was affiliated to genus Gammaproteobacteria. Strains from Bacillus family included; Bacillus siamensis, Bacillus licheniformis, Bacillus methylotrophicus, Bacillus amyloliquefaciens, Bacillus pumilus, and Bacillus sonorensis while one strain that belonged to Alcaligenes family was Alcaligenes feacalis.
Key words: Hypersaline, Little Magadi, alkaliphilic, hot springs, filter-sterilized.
Soda lakes and their arid counterparts, the soda deserts are extreme environments often found in remote locations of subtropical and continental interiors (Kambura et al., 2016). Some of the best-studied soda lakes are Mono Lake in U.S. A. Lake Wadi An-Natrun in Egypt and Lake Magadi in Kenya. pH values for these lakes is commonly greater than 10 reaching pH 12 occasionally with large amounts of Na2CO3, (usually as Na2CO3.10H2O or Na2CO3, NaHCO3.2H2O) that are depleted of Mg2+ and Ca2+ due to the insoluble carbonates under alkaline conditions (Duckworth et al., 1996; Sorokin et al., 2013; Vavourakis et al., 2016). Due to their location in closed basins, the rates of evaporation often exceed any inflow. The low pH lakes are the most productive aquatic environments in the world despite their extreme conditions like high temperatures and high salinity (Grant, 2006; Nyakeri et al., 2018).
Lake Magadi in Kenya for example is characterized by extensive salt flats and is the world’s second-largest source of sodium carbonate (Bell et al., 2005). Salinity levels go up to 30% w/v (Deshmukh et al., 2011). Extremophiles are inhabitants of these harsh environments since they are so well-adapted to readily grow and multiply in these environments as elucidated by previous studies (Sorokin et al., 2016). Some of the habitats combine high pH, temperature and alkalinity; organisms adapted to these conditions have been referred to as polyextremophiles (Tamura et al., 2007; Mesbah and Wiegel, 2012). From anthropogenic point of view prokaryotes grow and survive under conditions considered harsh and inhospitable (Mesbah and Wiegel, 2012). Detailed microbiological investigations have been carried out on the lakes over many years (Nyakeri et al., 2018). Lake Magadi supports the growth of a remarkable collection of these microbes that could be of economic and ecological importance (Antony et al., 2012).
Extremophiles possess useful physiological properties; for instance they are adapted to high alkalinity and possess heat stable enzyme. This makes them suitable for exothermic industrial processes (Horikoshi, 1999; Kambura et al., 2016). Physiological adaptation to temperatures and high pH together with the possible use in biotechnological applications is what drives the interest in alkalithermophiles (Grant and Heaphy, 2010). It has been hypothesized that microorganisms from soda lakes may be a source of enzymes that function at high salinity, pH and temperature (Grant and Heaphy 2010). Alkali-stable extracellular proteases, lipases, and cellulases have been used for the production of improved laundry detergents. These enzymes have an additional advantage because they are frequently used in bioremediation (Yang et al., 2013) and in the production of bioethanol from organic waste (Sorokin et al., 2014). The number of isolates recovered from these environments remains low whereas culture-independent studies point to a very high diversity of uncultured organisms (Kambura et al., 2012; Yang et al., 2013). The purpose of this study was to conduct screening, isolation, and characterization of new extremophilic microorganisms from the hot springs of the hyper saline Lake Magadi in Kenya.
Sample collection
Sterilized plastic containers of 500 ml were used whereby a sterilized hand shovel was used to collect wet sediments which were then put into sterile 50 ml Falcon tubes. The top 5 mm from every microbial mat forming on the hot spring water boundaries was collected into sterilized 500 ml plastic bottleneck jars. The collected samples were put in sterile bottles and preserved cool boxes then transported to the laboratory at Institute of Biotechnology Research in Jomo Kenyatta University of Agriculture and Technology (JKUAT), Kenya. At the laboratory, the samples were preserved at 80°C.
Enrichment and isolation
Malt extract-yeast extract agar (ISP medium 2) and International Streptomyces Project (ISP) medium 5 (starch casein agar) was used to perform bacterial culture isolation (Ellaiah et al., 1996). Kanamycin of 75 mg/ml was added to the media to inhibit fungal contamination and the pH of the media was adjusted to 9.4. The filtered lake water was used to replicate the lake conditions mainly salinity and pH. Fresh microbial mat was then pulverized and mixed with a spatula on a large piece of sterilized paper. Three samples of 0.5 gram of microbial mat were weighed on a sterile aluminum foil and transferred straight away into three universal bottles containing 10 ml lake water (Janssen et al., 2002). These preparations were mixed by approximate vortexing of 150 rpm for 1 min. Serial dilution of the microbial mat suspension with lake water was done in the ratio of 1 ml into 9 ml respectively. The dilutions were: 100, 10-1, 10-2 and 10-3. These preparations were then mixed by vortexing; 1 ml of aliquots was transferred to other 9 ml universal bottles. Aliquots of 200 μl from various dilutions were moved to petri dishes with the prepared media and laid on the surface using a sterilized spreading glass rod. Thereafter, incubation at 45oC for 24 to 72 h was done, followed by sub culturing of the bacterial colonies until a pure colony was obtained. Morphological features like pigmentation and size characterization was done on the colonies that had grown on the plates. Colonies that were well differentiated and isolated were transferred to 20% glycerol stocks and stored at -20°C.
Morphological, physiological and biochemical characterization
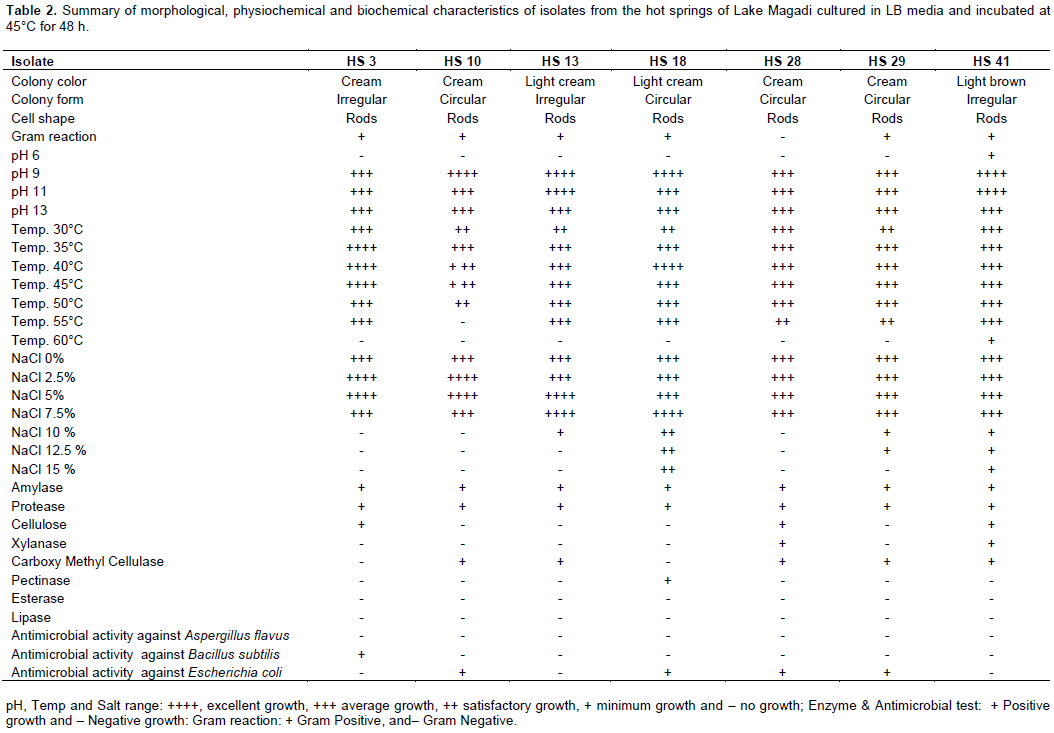
Cultural and morphological features were used in carrying out initial characterization where a compound and dissecting microscope were used to conduct morphological identification of the isolates that included characteristics of the cell, size and its shape after a classified staining of bacteria (Bartholomew, 1962). The isolates were screened by checking for their ability to grow on nutrient agar with different NaCl concentrations (0-17.5%), different pH levels (6-13) and growth at various temperatures (30-60ºC) according to Bergey’s Manual of Systematic Bacteriology (Holt et al., 1994). Culturing the isolates separately on basal media (1.0g K2HPO4, 0.1 g MgSO4, 7H2O, 0.05 g CaCl2.7H2O and 15.0 g of agar) supplemented with 5 g of cellulose, 1.25 g of xylan, 5.0 g of starch, 5 ml of olive oil and 1 % skim milk respectively was used to study the isolates’ ability to produce amylase, xylanase, cellulose, protease, lipase and esterase enzymes. For esterase production, basal media supplemented with previously sterilized 1% (v/v) Tween20 was used. All the plates inoculated with the substrates were incubated for 4 days at 45°C. A clear zone around the colony after 4 days of incubation was taken as evidence of proteolytic activity. The presence of lipase activity was demonstrated by a white halo due to the formation of precipitates of calcium laurate around the isolate while the presence of amylolytic activity on the plates was determined by flooding the plate with 0.6% KI solution. The presence of cellulases and xylanases was demonstrated by the formation of halos around the colony after flooding the isolates with Congo red dye for 2 min then pouring it out and subsequently rinsing the plates with NaCl (1 M) (Fatope et al., 2000).
In order to screen the isolates for antimicrobial activity, a cell based screening strategy was performed which showed the ability of each isolate to prevent growth of test organism. Incubation at 37°C in a shaker incubator was conducted for five days for each isolate that was cultured onto nutrient broth media allowing sufficient air circulation and preventing fermentation that could lead to acidity in the media. The test organisms were incubated at 45°C for 24 h and were also cultured in nutrient broth. There was a preparation of paper discs which were then impregnated with 10 ml of cell free broth isolate which were then dried under a fume chamber and placed on nutrient agar seeded with test organisms; Aspergillus flavus, Bacillus subtilis and Escherichia coli, which were incubated for 24 to 48 h at 37°C followed by recording of results.
Molecular characterization of pure bacterial colonies
Extraction of total bacterial DNA was done in accordance with the conventional phenolic-chloroform extraction and isopropyl alcohol precipitation procedures as described (Sambrook et al., 1989). Forward bacterial Primer pair 8F 5’-AG (A/G) GTTTGATCCTGGCT-3’) and reverse bacterial primer 1492R5’CGGCTACCTTGTTACGACTT-3’ were used to conduct PCR amplification resulting in 1500 bp of the PCR product. The total 50 ul PCR reaction volume contained 0.25 μl of high fidelity Taq, 1.25 μl (10 pmol) of 8F forward primer, 1.25 μl (10pmol) of 1492R reverse primer, 0.5 μl of template DNA, 0.5 ul of dNTPs mix (2.5mM), 4.0 10x PCR buffer (Genescript, China) and 36.25 μl of PCR water. The PCR cycling consisted of an initial activation of the enzyme at 94°C for 5 min followed by 35 cycles of a denaturation step at 94°C for 45 s, primer annealing step at 55°C for 60 s chain elongation step at 72°C for 2 min and a final extension at 72°C for 5 min (Embley and Stackebrandt, 1994). Presence of the PCR product was verified on 1.5% agarose gel and visualized under UV light. Purification of PCR products was done using the QIAquick PCR purification Kit protocol (Qiagen, Germany) according to the manufacturer’s instructions (Marmur, 1961). ABI prism big dye terminator was used to sequence purified products at Macrogen while viewing and editing of 16S rRNA gene sequences of the bacterial isolates was done by using Chromas software (www.technelysium.com.au). The sequences were aligned using CLUSTAL W 1.6 software and then run using the BLAST search algorithm found in the National Center for Biotechnology Information (NCBI) website (http://www.ncbi.nlm.nih.gov/) to determine closely related bacterial 16S rRNA gene sequences from public databases. Phylogenetic analyses were conducted in MEGA version 7 (Tamura et al., 2007). Neighbor-Joining technique was used to deduce evolutionary history (Saitou and Nei, 1987). The Jukes and Cantor (1969) technique was used to calculate the evolutionary distances, deduce and construct the phylogeny tree. The bootstrap consensus tree deduced from 10,000 replicates (Felsenstein, 1985) was taken to denote the evolutionary history of the already analyzed taxa. The evolutionary distances were computed through the Maximum Composite Likelihood technique (Robertson et al., 2009).
Sampling site physical characteristics
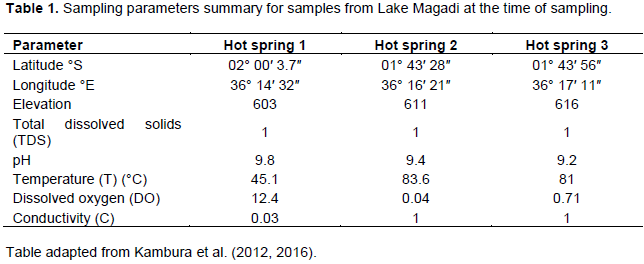
Table 1 presents the physical parameters of the lake.
Morphological, physiochemical and biochemical characterization of the Isolates
Classical macroscopic techniques of shape, form, color and elevation of pure colonies were the bases under which morphological characterization was conducted whereby most colonies managed to grow within 2-3 days at 45°C. The colony morphology of the bacterial isolates ranged from, flat and filamentous or branching (Table 2). The colony colors ranged from brown, cream and white while the margins were either rough or smooth. Growth pH optima of the bacterial isolates ranged from 9 to 11 and optimum salt (NaCl) concentration ranging from 2.5 to 7.5% w/v. Most of them were slight thermophiles with growth temperature optima ranging from 40 to 50°C (Table 2). Hydrolysis of starch, proteins, cellulose, pectin, CMC and xylan was used while testing the ability of the bacterial isolated to excrete extracellular enzymes thus showing the ability to produce the respective enzymes (Table 2 and Figure 1). The colonies also exhibited antimicrobial activity against Aspergillus flavus, Bacillus subtilis and Escherichia coli.
Molecular characterization
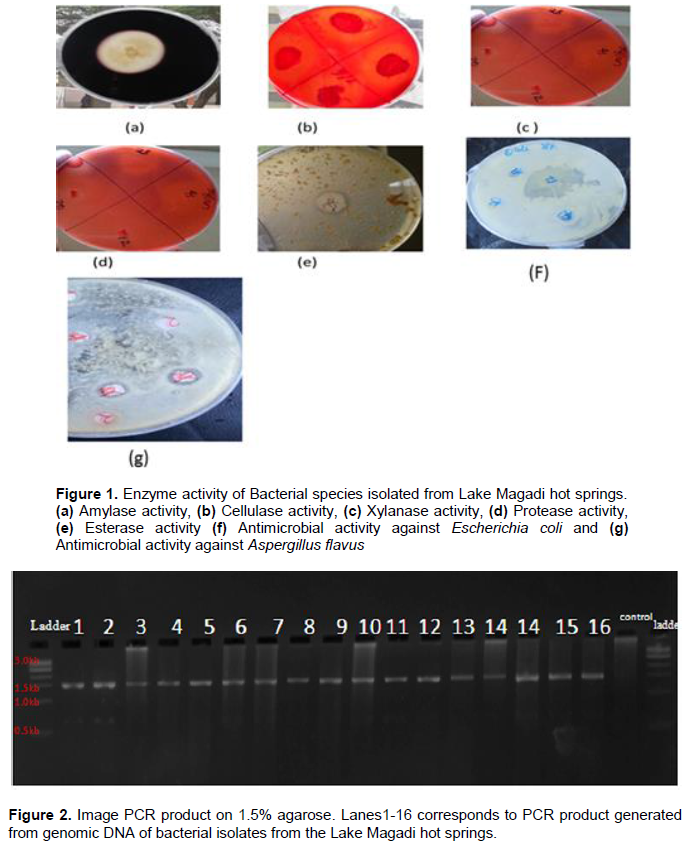
The amplification of the 16S rDNA region of the genomic DNA was conducted by the use of universal bacterial primers bac 8F and bac 1492R. The 16S rRNA gene amplification yielded an amplicon of approximately 1500 bp using the above primers (Figure 2). Blast analysis indicated that six isolates originated from the genus Bacillus within the Firmicutes phylum and had similarities between 86 and 98% while one isolate belonged to genus Alcaligenes in the phylum proteobacteria with a similarity of 96%. These were: Bacillus siamensis, Bacillus licheniformis, Bacillus methylotrophicus, Bacillus amyloliquefaciens, Bacillus pumilus, Bacillus sonorensis and Alcaligenes feacalis (Table 3 and Figure 3).
The existence of terrestrial hot springs on earth represents hotspots for unusual forms of life, genes, and metabolites. Several authors have carried out studies and characterized the microbiological inhabitants of the East African Rift Valley lakes (Vavourakis et al., 2016). The isolates subjected to phenotypic, chemotaxonomic and phylogenetic analysis have shown considerable phylogenetic diversity (Mwirichia et al., 2010). This research was conducted to isolate a large number of useful alkalithermophiles from Lake Magadi hot springs, then characterizing and identifying them using biochemical, morphological and molecular methods. Both Gram positive and Gram negative isolates were recovered in this study (Table 2). The gram staining results were confirmed by blast results that identified the majority of the isolates as gram positive isolates and related to genus Bacillus.
Physiochemical characterization of the isolates showed that the highest growth was observed between pH 9 to 12. There was an isolate that managed to grow at a pH 6 which showed that it was capable of tolerating the acidic conditions (Moreira and Siqueira, 2002; Kambura et al., 2012). An earlier study by Horikoshi (1999) indicated that a low pH of 5.7 - 9.0 was favorable for the growth of alkaliphiles and that a pH ranging 9.0 to 10 served as their selective optimum pH. Microorganisms living at extreme pH are able to do so by retaining their cytoplasm at the same pH as their mesophilic families, hence reducing the need for evolution of changed internal physiology (Kambura et al., 2016). pH in the neutral spectrum is achieved by several mechanisms. The acidic polymers of the cell walls play a role as a negatively charged matrix which reduces the pH value at the cell surface. Alternately, the plasma membranes use the Na+/H+-antiporter system and the ATPase driven hydrogen ion (H+) expulsion to maintain at least neutrality by lowering the internal pH (or below neutral cytoplasmic acidification) of the intracellular milieu (Mesbah and Wiegel, 2012). The cell membrane of alkaliphiles also contains a special composition for protecting them in the high alkaline conditions of the lake.
All the isolates were capable of growing within temperature ranging between 30 to 60°C. The optimal growth temperature range was recorded at 40 to 50°C. Good growth was also recorded even at temperature of 55°C. The temperatures recorded at sampling points ranged from 45.1 and 83.6°C. This study is therefore a confirmation of an earlier study on bacterial strains which had been obtained from Lake Magadi and that clustered with neighbors growing well at temperatures ranging between 10 to 55°C (Baumgarte, 2003; Mwirichia et al., 2010).
Alkalithermophiles isolated from the hot springs of Lake Magadi showed tolerance to salt concentrations as high as 15% with the optimal growth being observed between 2.5 to 7.5% sodium chloride concentration. In this research, growth of isolates improved with a rising salt concentration from 0 to7.5% sodium chloride concentrations. Optimum growth was observed at 7.5% salt concentration. Isolated microorganism’s capability to grow at 5% and above sodium chloride concentration is a proof of them being true halophiles. It can be concluded therefore that bacteria are able to survive for thousands of years in fluid inclusions of salt deposits and they do so by adapting to these potentially deadly environments (Akhwale et al., 2015). The biophysical properties of the lipid membranes of bacteria and archaea are crucial to their sustenance in a wide variety of physical environment. Comparative studies have implicated that the phospholipid and glycolipid membrane of halophiles are an important adaptation to salinity (Nyakeri et al., 2018). Generally, the capability of the isolated microorganisms in this research to grow rapidly and to the optimum in physicochemical and environmental parameters of high pH, high temperature and salinity not favorable for growth of other microbes makes the organisms from the hot springs of Lake Magadi good research models on adaptation mechanisms to harsh environmental conditions (Nyakeri et al., 2018).
The hot springs of Lake Magadi possess alkalithermophiles that have the potential of producing useful metabolites like enzymes and antibiotics. Various extracellular enzymes produced by the isolates include; lipases, cellulases, amylases, proteases, and xylanases. These results are in conjunction with previous studies which suggest that species of the genus Bacillus have biotechnological importance because of their ability to produce extracellular alkaline enzymes like amylases, proteases and pectinases that are resistant to adverse temperatures and high pH (Sokorin et al., 2014). Peptides have been shown to be the vastest nitrogenous compounds in organic matter by other researchers and this concurs with this study finding.
During the study of the isolates for their antimicrobial compound production potential, an antagonistic activity was observed differing from one isolate to the other. This could be as a result of differing of the modes of activity and action of each biochemical constituent of the isolates that were represented. Majority of the secondary metabolites that are produced by bacteria are potential antibodies and this makes them to be targeted for isolation in industries for large scale screening. The microbes have become a commercial interest because they produce secondary metabolites that are anti-fungal, anti-bacterial or anti-protozoal (Berdy, 2005). Six isolates in this study were affiliated to microorganisms belonging to the genus Bacillus while one isolate belonged to genus Alcaligenes. Among these were; B. sonorensis, B. pumilus, B. licheniformis, B. methylotrophicus, B. siamensis, B. amyloliquefaciens and A. feacalis (Table 3). Bacillus species are among the most commonly found aerobic alkalithermophiles in soda lakes as well as selected environments (Mwirichia et al., 2010). Members of the genus Bacillus that have been previously isolated from Lake Magadi clustered with Bacillus members that are alkalitolerant, obligate alkaliphilic while others are alkalithermotolerant.
HS 3 closely clustered to B. siamensis (NR 117274.1) with scored 86% similarity. HS 10 closely clustered to B. lichenformis (NR 118996.1) with a similarity of 98%. B. licheniformis which is rod shaped produces a variety of extracellular enzymes associated with the cycling of nutrients in nature. B. licheniformis excretes a protease enzyme able to tolerate pH levels ranging between 9 and 10 and it is a desired ingredient in laundry detergent for preventing clothes against fading and shrinkage. B. licheniformis excretes an antibiotic called Bacitracin composed of cyclic polypeptides mixture and its ironical purpose is to stop the growth of B. licheniformis. Bacitracin lyses the cells of B. licheniformis in the presence of cadmium or zinc ions (Takami and Krulwich, 2000). B. licheniformis also has an antifungal activity and contributes to nutrient cycling as shown in current research.
HS 13 was closely related to B. methylotrophicus (NR 116240.1) with 90% homology. B. methylotrophicus is a gram positive bacterium; its cells are aerobic, motile, rod shaped and forms endospores (Beibei et al., 2016). B. methylotrophicus is used as plant growth-promoting rhizobacteria since it stimulates plant growth and development via emission of volatiles. HS 18 clustered to B. amyloliquefaciens (NR 137421.1) with 86% similarity. B. amyloliquefaciens is known for its ability to degrade extracellular proteins and also a source of BamH1 restriction enzyme. It produces enzyme subtilisin which is used in industries to make laundry detergents (Benardini et al., 2003).
Isolate HS 28 scored 96% similarity with Alcaligenes faecalis strain NR 113606.1. Several isolates from the hot springs of Lake Magadi have an affiliation to Gammaproteobacteria class members and the genera in this class constitute of: Brevibacterium Rhodobacter, Stenotrophomonas, Xenorhabdus Alcaligenes, Klebsiella, Anoxybacillus, Proteus and Rhodobacter, (Kambura et al., 2012; Nyakeri et al., 2018). Alcaligenes faecalis produces an extracellular Serine Protease that is used in agriculture as a nematicide. Isolate HS 29 had 87% similarity to B. pumilus (NR 116191.1). B. pumilus strain has been isolated from a wide range of plants, environmental surfaces and soils from the interior of Sonoran Desert basalt (Kaushal et al., 2017). B. pumilus produces lipopeptide antibiotic iturin A which is an attractive biopesticide with the potential to replace chemical-based pesticides for controlling pathogens. HS 41 showed similarity to B. sonorensis (NR 113993.1) and is named after the Sonoran Desert, where the organism was collected from. It is Gram positive, motile, catalase-positive and facultative anaerobic. B. sonorensis is a plant growth promoting Rhizobacterium that produces indole acetic acid, siderophore, chitinase and hydrogen cyanide that helps in improving growth, nutrition, and yield of crops (Palmisano et al., 2001; Thilagar et al., 2018).
The study presented here has identified useful bacteria from the hot springs of Lake Magadi as potential sources for amylases, cellulases, proteases, and xylanases. However, much more needs to be explored; constant sampling can raise chances to redeem more novel species which could possess commercial importance. Diversity studies on the springs of Lake Magadi have hinted presence of uncultured extremophiles. Therefore, there is a remaining challenge in obtaining more of the microbes in culture for the purposes of studying their physiology and classifying them to into the species and strain level. The isolates characterized in this study can be considered for commercial use by elevating production of metabolites for biotechnological applications.
The authors have not declared any conflict of interests.
The authors thank JICA-icmob for providing a Research Grant for this work.
REFERENCES
|
Akhwale JK, Göker M, Rohde M, Spoer C, Schumann P, Klenk HP, Boga HI (2015). Antonie van Leeuwenhoek 107(5):1249-1259.
Crossref
|
|
|
|
Antony CP, Kumaresan D, Hunger S, Drake HL, Murrell JC, Shouche YS (2012). Microbiology of Lonar Lake and other soda lakes. The International Society for Microbial Ecology Journal 7(3):468-476.
Crossref
|
|
|
|
|
Bartholomew JW (1962). Variables influencing results, and the precise definition of steps in Gram staining as a means of standardizing the results obtained. Stain Technology 37(3):139-155
Crossref
|
|
|
|
|
Baumgarte S (2003). Microbial diversity of soda lake habitats. Thesis, Brauchweig, Germany. Technische Universitat Braunschweig.
|
|
|
|
|
Beibei G, Binghua L, Thinn TN, Wenjun Z, Liming S, Kecheng Z (2016). Bacillus methylotrophicus strain NKG-1, isolated from Changbai Mountain, China, has potential applications as a biofertilizer or biocontrol agent. PLOS One 11(11):e0166079.
Crossref
|
|
|
|
|
Bell T, Newman JA, Silverman BW, Turner SL, Lilley AK (2005). The contribution of species richness and composition to bacterial services. Nature 436 (7054):1157-1160
Crossref
|
|
|
|
|
Benardini JN, Sawyer J, Venkateswaran K, Nicholson WL (2003). Spore UV and acceleration resistance of endolithic Bacillus pumilus and Bacillus subtilis isolates obtained from Sonoran desert basalt: implications for lithopanspermia. Astrobiology 3(4):709-717.
Crossref
|
|
|
|
|
Berdy J (2005). Bioactive microbial metabolites. Journal of Antibiotics 58(1):1-26.
Crossref
|
|
|
|
|
Deshmukh KB, Pathak AP, Karuppayil MS (2011). Bacterial Diversity of Lonar Soda Lake of India. Indian Journal of Microbiology 51(1):107-111.
Crossref
|
|
|
|
|
Duckworth AW, Grant WD, Jones BE, van Steenbergen R (1996). Phylogenetic diversity of soda Lake Alkaliphiles. FEMS Microbiology and Ecology 19(3):181-191.
Crossref
|
|
|
|
|
Ellaiah P, Kalyan D, Rao VS, Rao BV (1996). Isolation and characterization of bioactive actinomycetes from marine sediments. Hindustan Antibiotics Bulletin 38(1-4):48-52.
|
|
|
|
|
Embley TM, Stackebrandt E (1994). The molecular phylogeny and systematics of the actinomycetes. Annual Review of Microbiology 48:257289.
Crossref
|
|
|
|
|
Fatope MO, Aduom OA, Takeda Y (2000). C18 acetylenic fatty acids of Ximenia americana with potential pesticidal activity. Journal of Agricultural and Food Chemistry 48(5):1872-1874.
Crossref
|
|
|
|
|
Felsenstein J (1985). Confidence limits on phylogenies: An approach using the bootstrap. Evolution 39(4):783-791.
Crossref
|
|
|
|
|
Grant WD, Heaphy S (2010). Metagenomics and recovery of enzyme genes from alkaline saline environments. Environmental Technology 31(10):1135-1143.
Crossref
|
|
|
|
|
Grant WE (2006). Alkaline Environments and Biodiversity in Extremophiles. Encyclopedia of Life Support Systems (EOLSS), developed under the Auspices of the UNESCO, EOLSS Publishers, Oxford, UK.
|
|
|
|
|
Holt JG, Krieg NR, Sneath, PHA, Stanley JT, William ST (1994). Bergey's Manual of Determinative Bacteriology. Williams and Wilikins, Baltimore, pp. 786-788.
|
|
|
|
|
Horikoshi K (1999). Alkaliphiles: some applications of their products for biotechnology. Microbiology and Molecular Biology Review 63(4):735-750.
Crossref
|
|
|
|
|
Janssen PH, Yates PS, Grinton BE, Taylor PM, Sait M (2002). Improved Culturability of soil bacteria and isolation in pure culture of novel members of the divisions Acidobacteria, Actinobacteria, Proteobacteria and Verrucomicrobia. Applied and Environmental Microbiology 68(5):2391-2396.
Crossref
|
|
|
|
|
Jukes TH, Cantor CR (1969). Evolution of protein molecules. In: Munro HN, editor, Mammalian Protein Metabolism: 21-132, Academic Press, New York.
Crossref
|
|
|
|
|
Kambura AK, Mwirichia RK, Kasili RW, Karanja EN, Makonde HM, Boga HI (2016). Bacteria and Archaea diversity within the hot springs of Lake Magadi and Little Magadi in Kenya. BioMed Central Microbiology 16(136).
Crossref
|
|
|
|
|
Kambura AK, Mwirichia RK, Ngaira J, Boga HI (2012). Isolation and characterization of bacterial isolates from Lake Magadi. Journal of Tropical Microbiology and Biotechnology 8(1):17-25.
|
|
|
|
|
Kaushal M, Kumar A, Kaushal R (2017). Bacillus pumilus strain YSPMK11 as plant growth promoter and bicontrol agent against Sclerotiniasclerotiorum. 3 Biotech 7(2):90.
Crossref
|
|
|
|
|
Marmur JA (1961). Procedure for the isolation of deoxyribonucleic acid from micro-organisms. Journal of Molecular Microbiology 3(2):208-218.
Crossref
|
|
|
|
|
Mesbah NM, Wiegel J (2012). Life under multiple extreme conditions: Diversity and physiology of the halophilic alkalithermophiles. Applied and Environmental Microbiology 78(12):4074-4082.
Crossref
|
|
|
|
|
Moreira FMS, Siqueira JO (2002). Microbiologia e bioquímica do solo. Lavras: Universidada Federal de Lavras.
|
|
|
|
|
Mwirichia R, Muigai AW, Boga HI, Stackebrandt E (2010). Isolation and characterization of bacteria from the haloalkaline Lake Elmenteita, Kenya. Extremophiles 14(4):339-348.
Crossref
|
|
|
|
|
Palmisano MM, Nakamura LK, Duncan KE, Istock CA, Cohan FM (2001). Bacillus sonorensis sp. nov., a close relative of Bacillus licheniformis, isolated from soil in the Sonoran Desert, Arizona. International Journal of Systematic and Evolutionary 51:1671-1679.
Crossref
|
|
|
|
|
Nyakeri EM, Mwirichia R, Boga H (2018). Isolation and characterization of enzyme producing bacteria from Lake Magadi, an extreme soda lake in Kenya. Journal of Microbiology and Experimentation 6(2):57-68.
Crossref
|
|
|
|
|
Robertson CE, Spear JR, Harris JK, Pace NR (2009). Diversity and stratification of archaea in a hypersaline microbial mat. Applied Environmental Microbiology 75(10):1801-1810.
Crossref
|
|
|
|
|
Saitou N, Nei M (1987). Neighbour Joining Method: a new method for reconstructing phylogenetic trees. Molecular Biology and Evolution 4(4):406-425.
|
|
|
|
|
Sambrook J, Fritsch EF, Maniatis T (1989). Molecular Cloning: A Laboratory Manual. 2nd Edition. Cold Spring Harbor Laboratory Press, New York.
|
|
|
|
|
Sorokin DY, Banciu H, Robertson LA, Kuenen JG, Muntyan MS, Muyzer G (2013). Halophilic and Haloalkaliphilic Sulfur-Oxidizing Bacteria. In: Rosenberg E, DeLong EF, Lory S, Stackebrandt E, Thompson F (editors) The Prokaryotes. Springer, Berlin, Heidelberg.
Crossref
|
|
|
|
|
Sorokin DY, Berben T, Melton ED, Overmars L, Vavourakis CD, Muyzer G (2014). Microbial diversity and biogeochemical cycling in soda lakes. Extremophiles 18(5):791-809.
Crossref
|
|
|
|
|
Sorokin DY, Kublanov IV, Gavrilov SN, Rojo D, Roman P, Golyshin PN, Slepak VZ, Smedile F, Ferrer M, Messina E, La Cono V, Yakimov MM (2016). Elemental sulfur and acetate can support life of a novel strictly anaerobic haloarcheon. The International Society for Microbial Ecology Journal 10:240-252.
Crossref
|
|
|
|
|
Takami H, Krulwich TA (2000). Re-identification of facultative alkaliphilic Bacillus firmus OF4 as Bacillus pseudofirmus OF4. Extremophiles 4(1):19-22.
Crossref
|
|
|
|
|
Tamura H, Yamada A, Kato H (2007). Identification and characterization of a dextranase gene of Streptococcus criceti. Microbiology and Immunology 51(8):721-732.
Crossref
|
|
|
|
|
Thilagar G, Bagyaraj DJ, Podile AR, Vaikuntapu PR (2018). Bacillus sonorensis, a novel plant growth promoting Rhizobacterium in improving growth, nutrition and yield of (Capsicum annuun L.). Proceedings of the National Academy of Sciences, India Section B: Biological Sciences 88(2):813-818.
Crossref
|
|
|
|
|
Vavourakis CD, Ghai R, Valera FR, Sorokin DY, Tringe SG, Hugenholtz P, Muyzer G (2016). Metagenomic insights into uncultured diversity and physiology of microbes in four hypersaline soda lake brines. Frontiers in Microbiology 7:211.
Crossref
|
|
|
|
|
Yang ZJ, Fu L, Zhang GW, Yang Y, Chen SY, Wang J, Lai SJ (2013). Identification and association of SNPs in TBC1D1 gene with growth traits in two rabbit breeds. Asian-Australasian Journal of Animal Sciences 26(11):1529-1535.
Crossref
|
|