Full Length Research Paper

ABSTRACT
This study was carried out to investigate changes in proximate and chemical components of cassava root sieviates (CRS) treated with extracted enzymes obtained from (a) Aspergillus niger (An); (b) Rhizopus stolonifer (Rs); (c) Trichoderma viridae (Tv) and (d) Mucor mucedo (Mm) applied on autoclaved CRS at 250 ml/kg. Another treatment had a commercial enzyme Roxazyme G2G (RG2G) as the degrading agent. At the end of the seventh day after enzyme application, crude protein, ash and metabolizable energy content increased but crude fibre, pectin, neutral detergent fibre (NDF), acid detergent fibre (ADF), acid detergent lignin (ADL), hemicellulose and cellulose content decreased. The highest value of metabo-lizable energy (ME), ash and crude protein were: 2807.81 kcal/kg, 17.32 and 18.32 g/100g dry matter, respectively obtained when enzymes obtained from A. niger, T. viridae and A. niger were added respectively. Results of levels of sugars (mg/mL) in undegraded CRS showed that glucose level increased by 72.4, 67.2, 53.8, 50.1 and 32.5% when enzyme preparations from A. niger, T. viridae, R. stolonifer, M. mucedo and Roxazyme G2G were applied on CRS respectively. Results obtained indicate that the use of enzymes from the above named fungi defiberised the CRS and hence promoted better bioavailability of the hidden nutrients in it.
Key words: Cassava root sieviate, enzymes, biodegradation, nutrient enhancement.
INTRODUCTION
Feed insecurity describes a situation whereby there is shortage of feed ingredients in quantity or in quality and this is aggravated by the ever widening demand for conventional feed sources in developing countries. No doubt, there is continuous need for animal protein and low availability of it is the chief factor known for the occurrence of child/infant morbidity, kwashiorkor, poor level of productivity and reduction of life span. Agromisa (2006) revealed that the state of consumption of meat and other animal protein in Nigeria is estimated at about 8 g per caput per day and this is about 27g less than the 35 g per caput minimum requirements recommended by the Food Agriculture Organization (FAO, 2003). Besides, in Nigeria, Olerede (2005) noted that poultry constitute over 90% of the current national livestock population and are of appreciable economic and social value to the investors and consumers. Large scale egg production is one of the ways to ensure better availability of animal protein. Regmi (2007) observed the unprecedented population growth that has occurred in the last half of the century has created an additional demand for meat and general food in developing countries. The prices of conventional feed ingredients keep rising steadily because of the competition between man and livestock and their increasing populations. The cost of feeds is as high as 70 to 75% of the total cost of production (Oluremi et al., 2007). In an attempt to overcome this challenge, several studies have been carried out on the possibility of using agro industrial by products and crop residues. Therefore, of late, agro industrial by-products are receiving attention. Meanwhile, agro industrial by-products (AIBs) that have their source from plants have cell walls which contain a variety of polysaccharides, the distribution of which varies within primary and secondary cell wall and between mono and dicotyledonous plants. The polymers are interlinked by covalent linkages or via non-carbohydrate compounds. The non-starch polysaccharides (NSPs) comprise 700-900 g/kg of the plant cell wall with the remaining being lignin, protein, fatty acids and waxes. Plant cell wall NSP is a diverse group of molecules with varying degree of water solubility, size and structure which may influence the rheological properties of the gastro intestinal content (Taibipour and Kermanshahi, 2004; Yineth and Mario, 2014). AIBs constitute those parts of crops that are left after removal of value giving components. These residues still contain considerable amount of energy and protein which may be present as intracellular compounds (Huang et al., 2008). AIBs represent potential valuable and renewable resources which find application in various areas that include use as animal feed. AIBs have been successfully incorporated into poultry diets at various levels in developing countries with resultant effect on reduction of cost of feeding. The use alleviates the existing critical situation of inadequate feed supply. Numerous successful studies on the supplementation of agro-industrial by-products with enzymes have been reviewed by several authors (Kelly-Yong et al., 2005; Khajavi et al., 2005). Lignin has been recognized as the chief barrier to monogastric digestion of cell wall structural polysaccharides (Kumar et al., 2009) and removal of lignin by chemical treatment enhances fibre digestibility (Oms-Oliu et al., 2009). Lignin offers the recalcitrant and adamant posture to fibre by the strength of its chemical nature though lignin may not be solely responsible for the variation in digestibility that is in AIBs (Pandey and Nagveni, 2007; Bachtar, 2005). In Nigeria, large quantities of CRS are produced by the cassava tuber processing industries (Aderemi, 2000). The aim of this study was to investigate possibility of defiberising the polymers that are interlinked by covalent linkages in the CRS and study the changes in proximate and sugars levels components of cassava root sieviates treated with extracted enzymes obtained from Aspergillus niger (An), Rhizopus stolonifer (Rs), Trichoderma viridae (Tv) and Mucor mucedo (Mm) and to possibly recommend the most effective biodegrader of CRS among the four fungi and the commercial enzyme.
MATERIALS AND METHODS
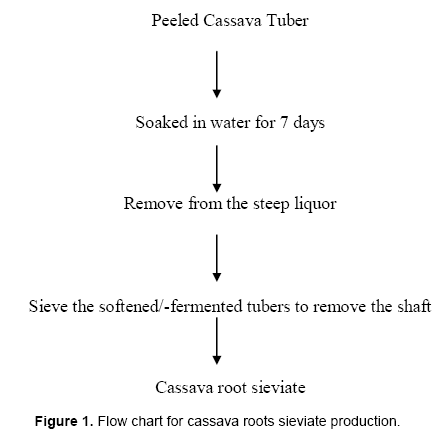
The CRS used for this study was obtained from a cassava pro-cessing centre in Ibadan, Nigeria. Figure 1 shows the flow chart for cassava roots sieviate production. It was milled with a 0.84 mm sieve and autoclaving was done at 121°C for 15 min.
Sources of fungi used
R. stolonifer and A. niger were obtained by exposing a piece of moist bread and a dough made from cassava flour to the air for 24 h and then covered for 3 days. After the third day, growth of the fungi appeared on the bread and cassava dough. Pure T. viride was obtained from the Department of Microbiology, University of Ibadan.
Isolation of fungi
A sterile wire loop was used to collect the spores and the mycelia of the actively growing fungi. The spores and mycelia were then inoculated aseptically on the centre of sterile potatoes dextrose agar (PDA) plates. The spores and the mycelia were aseptically spread on the PDA plates using the sterile wire loop in a sterilized chamber. The inoculated plates were then incubated at 34°C in a Gallenkamp incubator. The PDA plates were examined for growth after 48 h. It was observed that there was mix culture of fungi. Spores from each fungus was aseptically isolated and subcultured on fresh sterile PDA and reincubated for 48 h. Pure culture was then obtained and the mycelium from this was put on slant of sterile PDA. The characterization of the obtained A. niger and R. stolonifer was known by the use of manual of Barnett and Hunter (1992).
Enzyme production procedure
Enzymes were produced by extraction from cassava root sieviate. The fungi used were A. niger, R. stolonifer and T. viridae. A com-mercial exogenous enzymes (Roxazyme G2G) that contained cellulase (endo-1, 4-(β-glucanase), β- glucanase (endo-1, 3 (4)-β-glucanase) and xylanase (endo-1, 4- β xylanase) was also used. Every 50 g of the substrate was moistened with 20 mL of the requisite basal medium (KNO3, 5.0 mg; KH2PO4, 2.0 g; MgSO4.7H2O, 0.5 g; Tryptone, 0.5 g; FeSO4.4H2O, 3.5mg; C6H5NO2, 0.5 mg; C12H17ClN4OS, 0.05 mg and C10H16N2O3S. Biotin, 0.05 mg per litre of distilled H2O) and then 1.0 ml of an aqueous spore’s suspension of each isolate was added. The conical flasks were covered with sterilized cotton wool and kept in the incubator at 330C for seven days. After the growth of the fungi, the contents of each flask were mixed with 100 mL of the requisite buffer (Phosphate; pH of 7.2) and then filtered through double layered muslin clothe. The filtrate in the flask was in a chilled environment (4°C) to prevent dena-turation of the enzymes. It was centrifuged at 3000 rpm for 15 min by using the centrifuge manufactured by Measuring and Scientific Equipment, MSE (UK) Ltd named MSE Cellsep 6/720R centrifuge. The supernatant was collected and taken as raw enzymes (Onilude and Oso, 1999). Raw enzymes were concentrated 5-fold by means of a Vacuum Rotator Evaporator at 3000 rpm. The concentrated enzymes were dialyzed using phosphate buffer of pH 7.2 (0.1M). The in vitro dry matter enzymic degradation (IVDMED) was carried out in three replicates per treatment. Ground and sterilized CRS was obtained from the mentioned fungi. The enzymes were directly applied on CRS at 250 ml/kg and allowed to stay for seven days. Mouths of conical flasks that contained the CRS and enzymes were plugged with sterilized cotton wool to prevent contamination. This was carried out in a sterilized environment. At the end of the seventh day, samples were oven dried at 70°C for 16 h to stop further actions of enzymes. This was tagged biodegraded CRS. Soluble sugars were determined spectrophotomerically using the methods of Association of Official Analytical Chemists (1995).
Chemical and statistical analyses
After drying, all the samples were milled with a 0.84 mm sieve and the following analyses were carried out: crude protein, crude fibre and ether extract, using Association of Official Analytical Chemists, AOAC (1995) method while acid detergent fibre, neutral detergent fibre, cellulose and acid detergent lignin were determined using the method of Van Soest and Robertson (1991). The absorbencies were read from the spectrophotometer at the specific wavelength 105°C for 8 h. Hemicellulose was estimated as the difference for each sugar. Dry matter was determined by drying the samples at between neutral detergent fibre and acid detergent fibre. Crude protein was determined as Kjeldahl nitrogen x 6.25. metabolizable energy (ME) of degraded and undegraded GNP samples was determined with the use of Pauzenga (1985) method: ME = 37 x CP% + 81.8 x fat% + 35.5 x NFE%. Data collected were subjected to analysis of variance (SAS, 1999). Significant differences between means were determined using the Duncan multiple range test (SAS, 1999).
RESULTS AND DISCUSSION
Results of the enzymic biodegradation of cassava root sieviate (CRS) as shown in proximate and detergent fibre composition are presented in Table 1. Enzymes increased the crude protein content of the CRS by 37.14, 40.99, 32.61, 32.52 and 16.26% with A. niger, R. stolonifer, M. mucedo, T. viride and Roxazyme G2G respectively. Values were significantly different (P<0.05) among fungi. Increase in the crude proteins value of the degraded AIBs was partly due to the ability of the enzymes to increase the bioavailability of the protein hitherto encapsulated by the cell walls. Fungal enzymes have the potential of improving not only the NSPs but also the crude proteins as well as other dietary components such as ash and fatty acids (Liu and Baidoo, 2005). Crude fibre content in the CRS was also significantly (P<0.05) reduced. A. niger caused the highest reduction in crude fibre level by reducing it from 10.98 g/100 g to 4.82 g/100 g which represented a 56.10% reduction and this was followed by changes orchestrated by T. viridae (49.73%). The cellulose content in the degraded CRS decreased by 36.3, 30.9, 24.0, 18.3 and 12.6% with T. viride, A. niger, M. mucedo, R. stolonifer and Roxazyme G2G, respectively. This confirms the reports by other authors (Iyayi and Aderolu, 2004; Iyayi and Losel, 2001; Yoshinori et al., 2014). Iyayi and Aderolu (2004) reported reduction in crude fibre content of brewer dried grain, maize offal and wheat offal when A. niger, A. flavus and Penicillium sp. were used for their biodegradation.
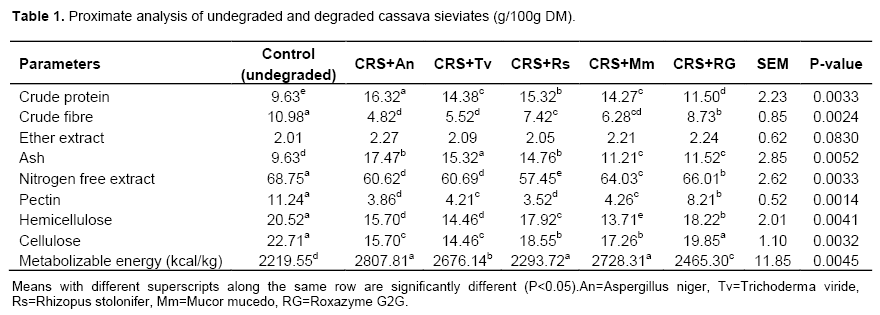
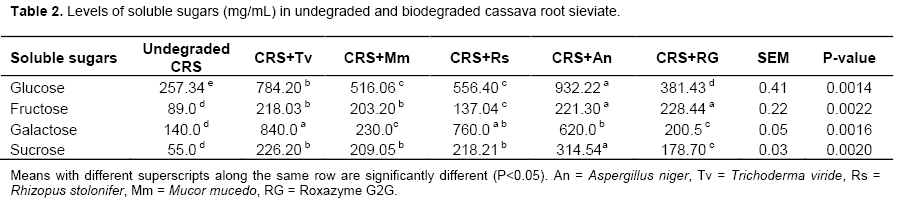
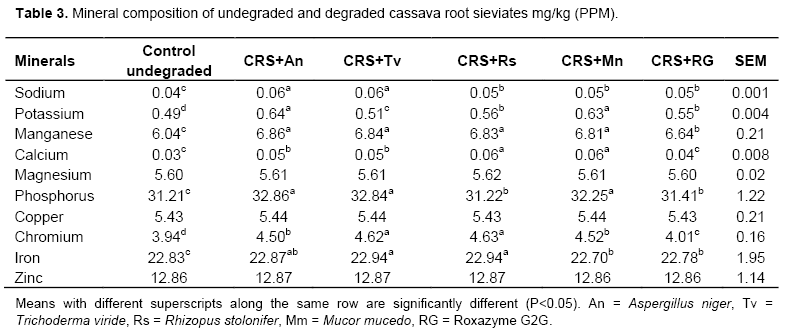
Crude fibre in the above mentioned AIBs were significantly (P<0.05) reduced by all the fungi until after the fourteenth day. A. niger consistently caused the highest reduction in crude fibre in all the AIBs followed by A. flavus and Penicillium sp. The use of enzyme has pre-dominantly been related to the hydrolysis of fibre or non-starch polysaccharides fractions in the AIBs. Glucans chains of cellulose are held together in an organized manner by inter and intra molecular hydrogen which renders the carbohydrates and other nutrients insoluble and resistant to enzymic hydrolysis. The structure and properties of β-glucans are described as polymer of glucose with a β-1, 4 linked backbones and β-1, 3 side linkages (Atik et al., 2006; Ezieshi and Olomu, 2004). Apart from the fact that β-glucans and the insoluble NSPs arabinoxylans are also found in crude fibre and they are in endosperm cell walls. Crude fibres were hydrolyzed by the synergetic action of xylanase and glucanases (Dare et al., 2010; Hughes et al., 2000). Table 3 shows mineral composition of undegraded and degraded cassava root sieviates. It reveals improvement in the potassium, phosphorus, calcium, iron, chromium and manganese content after enzymic degradation of CRS. The highest value (32.86 mg/kg) for phosphorus was found in the A. niger degraded sample. Possibly, enzyme phytase was part of the cocktail of enzymes produced by the used fungi and this must have assisted in the liberation of the phytate bound phosphorus. Phytate, like oxalates and tannins, is an organic compound (myo-inositol hexaphosphate) which occurs in all plants and these mineral binding factors possess anti-nutritional properties because they are potential chelators of minerals and, thus, their presence in any AIBs will strongly have negative effect on the digestibility of such materials (Sofia et al., 2014; Hannah et al., 2014). From the results, biodegradation of CRS by the fungi did enhance the bioavailability of the mineral elements (Na, K, Mg Ca, P and Fe). The results of sugar fractions (glucose, fructose, galactose and sucrose) in the undegraded and degraded CRS are shown in Table 2. Glucose was the highest produced sugar (glucose > fructose > sucrose > galactose). The highest glucose yield was in the CRS degraded with A. niger with a value of 932.22 g/mL as compared to 381.43 g/mL in the RG2G degraded CRS. The enzymes increased the metabolic energy content of the CRS by 21.0, 18.7, 17.1, 10.0 and 3.2% with A. niger, M. mucedo, T. viride, Roxazyme G2G and R. stolonifer, respectively. The values were significantly different (P<0.05) among the fungi. The increase in the sugar content and the metabolizable energy value of the degraded CRS was partly due to the ability of the enzymes to disrupt the cell wall structure (Ates et al., 2008; Gunal and Yasar, 2004). AIBs are known to have low ME values and they often have a high content of growth inhibiting, viscous, water-soluble, non starch polysaccharides (WNSPs). According to Oluremi et al. (2007), the monogastrics are unable to release the intrinsic energy portion of the AIBs as the energy remains latent in the fibrillian complex which renders resistance to the endogenous enzymes. In addition, Martins et al. (2000) observed a continuous increase in sugar production in the substrates until after the fourteenth day of fungi on the substrates when there was gradual reduction in sugar availability in the substrates. There were significant (P<0.05) increase in sugar production when A. niger, A. flavus and Penicillium species were inoculated on brewer dried grain, maize offal and wheat offal. This study shows that with fungal biomass increase, the nutrients in the substrate medium were quickly used up. Beyond fourteenth days, the fungi started using up the products of breakdown of the NSPs, hence, the observed reduction in the sugar level. There was increase in the ME because there was increase in the soluble sugars availability. Soluble sugars are the assimilable forms of carbohydrates needed for energy production. However, fungal enzymes added to the AIB were able to break the cell walls. Hence, the degraded CRS had higher values of sugars than the undegraded one and this shows the effects of the enzymes on the CRS.
CONCLUSION
This study leads makes us believe that the treatment of cassava root sieviate with extracted enzymes from A. niger, M. mucedo, R. stolonifer, T. viride and Roxazyme G2G may improve the crude protein, metabolizable energy, sugars and mineral and reduce the fibre fractions. Furthermore, the increase in crude protein and metabolizable energy of the degraded CRS due to enzymic degradation made the CRS beneficial thereby increasing its nutritive value and solving the problem of environmental pollution. The application of this technique helps in the con-version of AIBs into animal feeds thereby improving their preservation and utilization by livestock animals. Ultimately, this will have direct positive effect on availability of animal protein. Of the four fungi and the commercial Roxazyme G2G, enzymes from A. niger appeared to be the most suitable for the biodegradation of cassava root sieviate as it succeeded in effecting the highest improvement in terms of crude protein, crude fibre, ash, sugar and metabolizing energy. Therefore, the use of A. niger in enhancement of CRS as animal feed ingredient is preferred to T. viride, M. mucedo, R. stolonifer and Roxazyme G2G.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Aderemi F (2000). Enzymic supplementation of cassava root sieviate and its Utilization by layers. A Ph. D. thesis in Department of Animal Science, University of Ibadan. pp. 1-238 |
|
|
Agromisa A (2006). Agrodok small-scale chicken production. Co-published by CTA, wegeningen, the Netherland. p. 91. |
|
|
AOAC (1995). Official Methods of Analysis, 16th Edition. Association of Official Analytical Chemists, Washington D.C |
|
|
Ates S, Ni Y, Atik C, Imamoglu S (2008). Pretreatment by Ceriporiopsis subvesmispora and Phlebia subserialis of wheat straw and its impact on subsequent soda-AQ and Kraft-AQ pulping. Biotechnol. Lett. 13: 3914-3921. |
|
|
Atik C, Imamoglu S, Bermek H (2006). Impact of xylanase pretreatment on peroxide bleaching stage of biokraft pulp. Int. Biodeterior. Biodegrad. 58:22-26. |
|
|
Bachtar B (2005). Improvement of nutritive of crop by products using bioprocess technique and their uses for animals. J. Feed Technol. 21:1-5. |
|
|
Barnett HL, Hunter D (1992). Illustrated genes of imperfect fungi. Minnesota: Burgess Publishing Company, USA. |
|
|
Dare V, Papinutti L, Forchiassin F, Levin L (2010). Biobleaching of loblolly pine kraft pulp with Trametes trogii culture fluids followed by a peroxide stage: Application of Doehlert experimental design to evaluate process parameters. Enzyme Microb. Technol. 46:281-286. |
|
|
Ezieshi EV, JM Olomu (2004). Comparative Performance of Broiler Chickens Fed Varying Levels of Palm Kernel Cake and Maize Offal. Pak. J. Nutr. 3:254-257. |
|
|
Food and Agricultural Organization (FAO) (2003). Egg marketing. A guide for the production and sale of eggs. FAO Agricultural Services bulletin 150 Rome, Italy. p. 10. |
|
|
Gunal M, Yasar S (2004). Performance and some digest parameters of broiler chickens given low or high viscosity Wheat-Based Diets with or without Enzyme Supplementation. Turk. J. Vet. Anim. Sci. 28:323-327. |
|
|
Hannah L, Woo TC, Hazena BA, Simmonsa H, Kristen M, De Angelisa CI (2014). Enzyme activities of aerobic lignocellulolytic bacteria isolated from wet tropical forest soils. Syst. Appl. Microb. 37:60-67. |
|
|
Huang, D, Zeng G, Peng Z, Zhang P, Hu S, Jiang X, Feng C and Chen Y (2008). Biotransformation of rice straw by Phanerachaete chrysosporium and the related ligninolytic enzymes. Int. J. Biotechnol. 10:86-92. |
|
|
Hughes RJ, Choct M, Korcher A, Van Barneveld RJ (2000). Effect of food enzymes on AME and composition of digests from broilers chickens fed on diets containing non-starch polysaccharides isolated from pumpkin kernel. J. Nutr. 62:457-463. |
|
|
Iyayi EA, Aderolu ZA (2004). Enhancement of the feeding value of some agro industrial by products for laying hens after their solid state fermentation with Trichoderma viride. Afr. J. Biotechnol. 3(3):182-185. |
|
|
Iyayi EA, Losel DM (2001). Changes in carbohydrates fractions of cassava peel following fungal solid state fermentation. J. Food Technol. 6(3):10-13. |
|
|
Kelly-Yong TL, Lee KT, Mohamed AR, Bhatia S (2007). Potential of hydrogen from oil palm biomass as a source of renewable energy worldwide. J. Energy Policy 35:5692-5701. |
|
|
Khajavi SH, Kimura Y, Oomori T, Matsuno R, Adachi S (2005). Degradation kinetics of monosaccharides in subcritical water. J. Food Eng. 68:309-313. |
|
|
Kumar R, Mago G, Balan V, Wyman CE (2009). Physical and chemical characterizations of corn stover and poplar solids resulting from leading pretreatment technologies. Bioresour. Technol. 100:3948-3962. |
|
|
Liu YG and Baidoo SK (2005). Exogenous enzymes for Pigs diets: An overview. Proceedings of Asian conference on Feed Biotechnology 2005, pp. 67-70. |
|
|
Martins ES, Silva R, Gomes E (2000). Solid state production of thermostable pectinases from thermophilic Thermoascus aurantiacus. J. Process Biochem. 37:949-954 |
|
|
Olerede BR (2005). Non-conventional feed stuffs in poultry Nutrition. Trans-Akab Ltd., Sokoto, Nigeria |
|
|
Oluremi OIA, Ngi J, Andrew AI (2007). Phytonutrients in citrus fruit peel meal and nutritional implication for livestock production. Livestock Research for Rural Development. Volume 9, Article #89 http://www.cipav.org.co/lrrd19/7/olurl9089.htm |
|
|
Oms-Oliu G, Odriozola-Serrano I, Soliva-Fortuny R, Martín-Belloso O (2009). Use of Weibull distribution for describing kinetics of antioxidant potential changes in fresh-cut watermelon. J. Food Eng. 95:99-105. |
|
|
Onilude AA, Oso B A (1999). Effect of fungi enzyme mixture supplementation of various dietary fibres contains diets fed broiler chicks 1 performance and carcass characteristic. World J. Microbiol. Biotechnol. 15:17-19. |
|
|
Pandey KK, Nagveni H (2007). Rapid characterisation of brown and white rot degraded chir pine and rubberwood by FTIR spectroscopy. Eur. J. Wood Prod. 65:477-481. |
|
|
Pauzenga U (1985). Feeding parent-stock. J. Zoo Tech. Int. 19:22-23 |
|
|
Regmi A (2007). Urbanization and Food Consumption. A United States Development Authority City study available at http://search.ers.usda.gov/search?utf8=%E2%9C%93&sc=0&query=Urbanization+and+Food+Consumption&m=&affiliate=ers&commit=Search |
|
|
Sofia Q, Rashida RZ, Afsheen A, Shah AU (2014). Enhanced production of cellulose degrading CMCase by newly isolated strain of Aspergillus versicolor. Carbohydr. Polym. 104: 199-203. |
|
|
Statistical Analysis System (1999). SAS Users Guide. SAS Institute Inc. Cary, NC USA. |
|
|
Taibipour K, Kermanshahi H (2004). Effect of levels of tallow and NSP degrading enzyme supplements on nutrient efficiency of broiler chickens. In: Proceedings of the Annual Conference of the British Society of Animal Science, University of York, York, UK, 5-7 April, 2004, p. 273 |
|
|
Van Soest PJ, Robertson JB (1991). Methods for dietary fibre neutral detergent fibre and non-starch polysaccharides in relation to animal nutrition symposium: Carbohydrates, methodology, metabolism and nutritional implications in dairy cattle. J. Dairy Sci. 74:3583-3597. |
|
|
Yineth PC, Mario VL (2014). Biodegradation kinetics of oil palm empty fruit bunches by white rot fungi. Int. J. Biodeterior. Biodegrad. 91:24-28. |
|
|
Yoshinori S, Mutsumi A, Rika K (2014). Microbial deterioration of tsunami-affected paper-based objects: A case study. Int. Biodeterior. Biodegrad. 88:142-149. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


