Full Length Research Paper

ABSTRACT
This study evaluated the preservative ability of protein-like cell free supernatants produced by lactic acid bacteria (LAB) isolates from samples of Ictalurus punctatus (Cat fish) and slurry of fermented Zea mays (Ogi). The LAB strains were separately isolated from understudied samples using De Man, Rogosa and Sharpe (MRS) media at 37°C for 48 h. The isolated strains were characterized with Gram staining, oxidase and catalase tests, microscopy study, carbohydrate fermentation, acid production and NaCl tolerance. Thereafter, the protein concentrations of crude bacteriocin supernatants from the Gram positive, rod shaped, oxidase and catalase negative strains were studied. Also, the growth inhibition of Bacillus subtilis, Staphyloccocus aureus and Escherichia coli, heat stability, pH tolerance, effect of proteolytic enzyme and biopreservation efficiency of protein-like cell free supernatants (crude bacteriocins) were determined. Biopreservative efficiency of the crude bacteriocin samples was also determined in orange (Citrus sinenses) and Titus fish (Scomber scombrus). The isolates from intestine of I. punctatus and fermented Z. mays fermented carbohydrate, and grew optimally at 3% NaCl, and 10 and 37°C, respectively. They inhibited the multiplication of E. coli at various extents, but more effective on different strains. The bacteriocins from slurry of fermented Z. mays on the other hand, were more potent in E. coli (22.7 ± 0.8 mm) than S. aureus (7.9 ± 0.1 mm). The biopreservative efficiency of crude bacteriocin from I. punctatus was greater than that of Z. mays. The LAB obtained from the selected samples produced protein-like substances in form of bacteriocins with potent antibacterial and biopreservative proficiencies through the growth inhibition of tested pathogens and low colony counts on tested food samples, respectively. Bacterial isolates obtained from samples of I. punctatus and Z. mays can be successfully used in the preservation of food and vegetables.
Key words: Ictalurus punctatus, Zea mays, bacteriocin, protein-like substances, biopreservative ability.
INTRODUCTION
MATERIALS AND METHODS
RESULTS
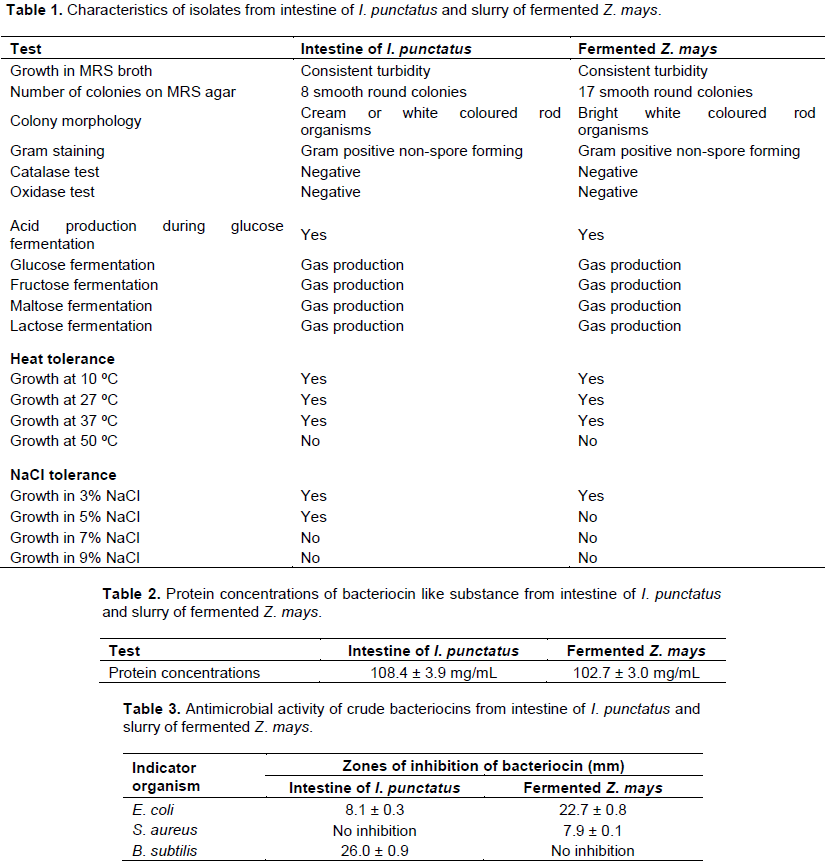
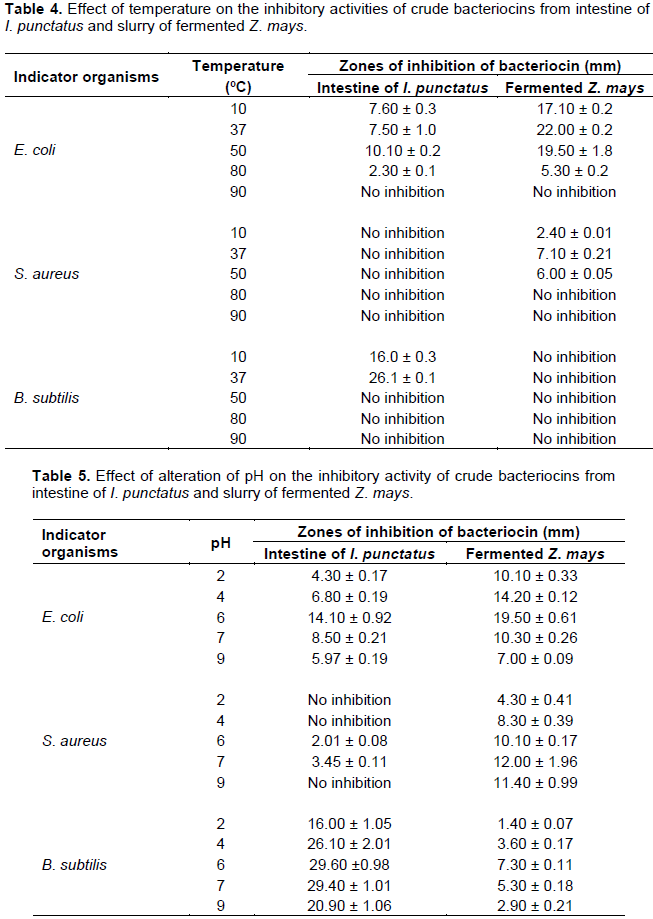
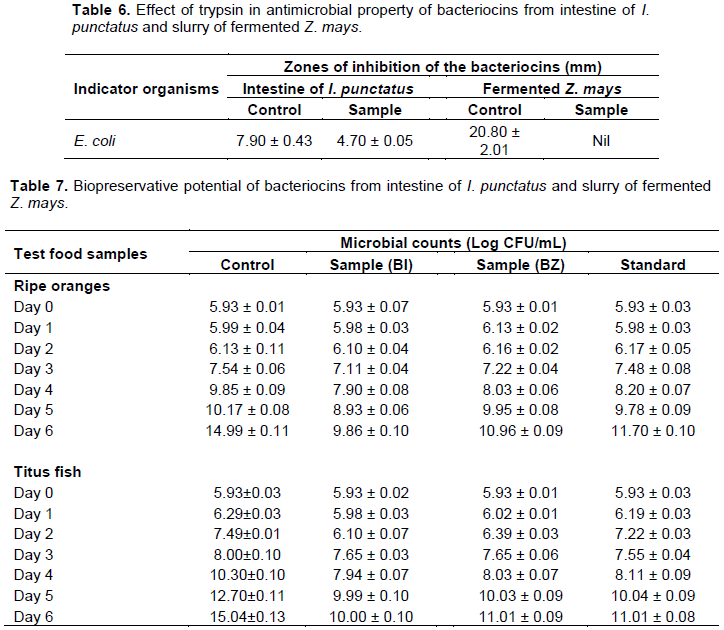
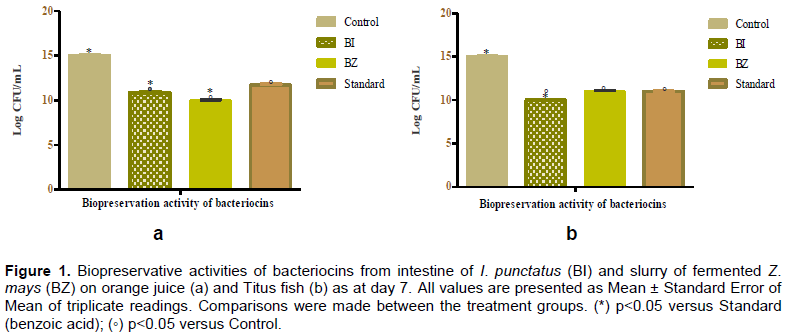
DISCUSSION
CONCLUSION
CONFLICT OF INTERESTS
ACKNOWLEDGEMENTS
REFERENCES
|
Abbasiliasi S, Tan JS, Ibrahim TAT, Ramanan RN, Vakhshiteh F, Mustafa S, Ariff AB (2012). Isolation of Pediococcus acidilactici Kp10 with ability to secrete bacteriocin-like inhibitory substance from milk products for applications in food industry. BMC Microbiology 12(1):260. |
|
|
Abriouel H, Franz C, Omar NB, Gálvez A (2011). Diversity and applications of Bacillus bacteriocins. FEMS Microbiology Reviews 35(1):201-232. |
|
|
Adesina I, Ojokoh A, Arotupin D (2016). Effect of bacteriocinogenic Pediococcus pentosaceus IO1 strain and its bacteriocin on growth performance and intestinal microbiota of albino rat. Microbiology Research Journal International. |
|
|
Adesokan I, Abiola O, Ogundiya M (2010). Influence of ginger on sensory properties and shelf-life of Ogi, a Nigerian traditional fermented food. African Journal of Biotechnology 9(12). |
|
|
Adnan AFM, Tan IK (2007). Isolation of lactic acid bacteria from Malaysian foods and assessment of the isolates for industrial potential. Bioresource Technology 98(7):1380-1385. |
|
|
Afolayan AO, Ayeni FA, Ruppitsch W (2017). Antagonistic and quantitative assessment of indigenous lactic acid bacteria in different varieties of ogi against gastrointestinal pathogens. The Pan African Medical Journal 27. |
|
|
Ageni L, Ajibade G, Yerima B, Appah J (2017). Shelf life extension study of ogi and fufu using bacteriocin isolated from Lactobacillus acidophilus of fermented dairy products. African Journal of Microbiology Research 11(32):1286-1293. |
|
|
Akalu N, Assefa F, Dessalegn A (2017). In vitro evaluation of lactic acid bacteria isolated from traditional fermented Shamita and Kocho for their desirable characteristics as probiotics. African Journal of Biotechnology 16(12):594-606. |
|
|
Al Kassaa I, Hamze M, Hober D, Chihib NE, Drider D (2014). Identification of vaginal lactobacilli with potential probiotic properties isolated from women in North Lebanon. Microbial Ecology 67(3):722-734. |
|
|
Ali AA (2010). Bene?cial role of lactic acid bacteria in food preservation and human health: a review. Research Journal of Microbiology 5(12):1213-1221. |
|
|
Alka S, Neelam Y, Shruti S (2012). Effect of fermentation on physicochemical properties and in vitro starch and protein digestibility of selected cereals. International Journal of Agricultural and Food Science 2(3):66-70. |
|
|
Alvarez-Sieiro P, Montalbán-López M, Mu D, Kuipers OP (2016). Bacteriocins of lactic acid bacteria: extending the family. Applied Microbiology and Biotechnology 100(7):2939-2951. |
|
|
Ashmaig A, Hasan A, El Gaali E (2009). Identification of lactic acid bacteria isolated from traditional Sudanese fermented camel's milk (Gariss). African Journal of Microbiology Research 3(8):451-457. |
|
|
Ayed HB, Maalej H, Hmidet N, Nasri M (2015). Isolation and biochemical characterisation of a bacteriocin-like substance produced by Bacillus amyloliquefaciens An6. Journal of Global Antimicrobial Resistance 3(4):255-261. |
|
|
Balcázar JL, Vendrell D, de Blas I, Ruiz-Zarzuela I, Muzquiz JL, Girones O (2008). Characterization of probiotic properties of lactic acid bacteria isolated from intestinal microbiota of fish. Aquaculture 278(1-4):188-191. |
|
|
Barakat OS, Ibrahim G, Tawfik N, El-Kholy W, El-Rab GD (2011). Identification and probiotic characteristics of Lactobacillus strains isolated from traditional Domiati cheese. International Journal of Microbiology Research 3(1):59. |
|
|
Barbosa AAT, Mantovani HC, Jain S (2017). Bacteriocins from lactic acid bacteria and their potential in the preservation of fruit products. Critical Reviews in Biotechnology 37(7):852-864. |
|
|
Behnsen J, Deriu E, Sassone-Corsi M, Raffatellu M (2013). Probiotics: properties, examples, and specific applications. Cold Spring Harbor Perspectives in Medicine 3(3):a010074. |
|
|
Chang JY, Chang HC (2010). Improvements in the quality and shelf life of kimchi by fermentation with the induced bacteriocin?producing strain, Leuconostoc citreum GJ7 as a starter. Journal of Food Science 75(2):M103-M110. |
|
|
Dallal MS, Zamaniahari S, Davoodabadi A, Hosseini M, Rajabi Z (2017). Identification and characterization of probiotic lactic acid bacteria isolated from traditional persian pickled vegetables. GMS Hygiene and Infection Control 12. |
|
|
Davidson PM, Taylor TM, Schmidt SE (2012). Chemical preservatives and natural antimicrobial compounds. Food Microbiology: Fundamentals and Frontiers pp. 765-801. |
|
|
De Vuyst L, Leroy F (2007). Bacteriocins from lactic acid bacteria: production, purification, and food applications. Journal of Molecular Microbiology and Biotechnology 13(4):194-199. |
|
|
De Vuyst L, Vandamme EJ (1994). Nisin, a lantibiotic produced by Lactococcus lactis subsp. lactis: properties, biosynthesis, fermentation and applications. Bacteriocins of Lactic Acid Bacteria. pp. 151-221. |
|
|
Deegan LH, Cotter PD, Hill C, Ross P (2006). Bacteriocins: biological tools for bio-preservation and shelf-life extension. International Dairy Journal 16(9):1058-1071. |
|
|
Del Nobile MA, Lucera A, Costa C, Conte A (2012). Food applications of natural antimicrobial compounds. Frontiers in Microbiology 3:287. |
|
|
Djadouni F, Kihal M (2012). Antimicrobial activity of lactic acid bacteria and the spectrum of their biopeptides against spoiling germs in foods. Brazilian Archives of Biology and Technology 55(3):435-444. |
|
|
Du Toit M, Engelbrecht L, Lerm E, Krieger-Weber S (2011). Lactobacillus: the next generation of malolactic fermentation starter cultures-an overview. Food and Bioprocess Technology 4(6):876-906. |
|
|
Ejigui J, Savoie L, Marin J, Desrosiers T (2005). Beneficial changes and drawbacks of a traditional fermentation process on chemical composition and antinutritional factors of yellow maize (Z. mays). Journal of Biological Sciences 5(5):590-596. |
|
|
Eun JB, Chung HJ, Hearnsberger J (1994). Chemical composition and microflora of channel catfish (Ictalurus punctatus) roe and swim bladder. Journal of Agricultural and Food Chemistry 42(3):714-717. |
|
|
Farkas J (2007). Physical methods of food preservation Food Microbiology: Fundamentals and Frontiers. Third Edition American Society of Microbiology pp. 685-712. |
|
|
Fontana L, Bermudez-Brito M, Plaza-Diaz J, Munoz-Quezada S, Gil A (2013). Sources, isolation, characterisation and evaluation of probiotics British. Journal of Nutrition 109(S2):S35-S50. |
|
|
Gaamouche S, Arakrak A, Bakkal M, LaglaouiA (2014). Antimicrobial activity of lactic acid bacteria and bacteriocins isolated from a traditional brine table olives against pathogenic bacteria. International Journal of Current Microbiology and Applied Science 3(11):657-666. |
|
|
Ghanbari M, Jami M, Domig KJ, Kneifel W (2013). Seafood biopreservation by lactic acid bacteria-a review. LWT-Food Science and Technology 54(2):315-324. |
|
|
Gómez-Sala B, Herranz C, Díaz-Freitas B, Hernánde PE, Sal A, Cintas LM (2016). Strategies to increase the hygienic and economic value of fresh fish: Biopreservation using lactic acid bacteria of marine origin. International Journal of Food Microbiology 223:41-49. |
|
|
Gómez-Sala B, Muñoz-Atienza E, Sánchez J, Basanta A, Herranz C, Hernández PE, Cintas LM (2015). Bacteriocin production by lactic acid bacteria isolated from fish, seafood and fish products. European Food Research and Technology 241(3):341-356. |
|
|
Gould GW (2012). New methods of food preservation: Springer Science and Business Media. |
|
|
Guetouache M, Guessas B (2015). Characterization and identification of lactic acid bacteria isolated from traditional cheese (Klila) prepared from cow's milk. African Journal of Microbiology Research 9(2):71-77. |
|
|
Hanim C (2017). Effect of pH and Temperature on Bacillus subtilis FNCC 0059 oxalate decarboxylase activity. Pakistan Journal of Biological Sciences 20(9):436-441. |
|
|
Ijarotimi SO, Keshinro OO (2011). Determination of amino acid, fatty acid, mineral, functional and choking properties of germinated and fermented popcorn (Z. mays everta) flour. European Journal of Nutrition and Food Safety pp. 102-122. |
|
|
Jose NM, Bunt CR, Hussain MA (2015). Comparison of microbiological and probiotic characteristics of lactobacilli isolates from dairy food products and animal rumen contents. Microorganisms 3(2):198-212. |
|
|
Klaenhammer TR (1988). Bacteriocins of lactic acid bacteria. Biochimie 70(3):337-349. |
|
|
Kozaki M, Uchimura T, Okada S (1992). Experimental manual of lactic acid bacteria. Tokyo, Japan: Asakurasyoten pp. 34-37. |
|
|
Lee HJ, Kim HJ (2011). Lantibiotics, class I bacteriocins from the genus Bacillus. Journal of Microbiology and Biotechnology 21(3):229-235. |
|
|
Li D, Ni K, Pang H, Wang Y, Cai Y, Jin Q (2015). Identification and antimicrobial activity detection of lactic acid bacteria isolated from corn stover silage. Asian-Australasian Journal of Animal Sciences 28(5):620. |
|
|
Liu W, Pang H, Zhang H, Cai Y (2014). Biodiversity of lactic acid bacteria. Springer pp. 103-203. |
|
|
Lück E (1985). Chemical preservation of food. Zentralblatt fur Bakteriologie, Mikrobiologie und Hygiene. 1. Abt. Originale B, Hygiene 180(2-3): 311-318. |
|
|
Mørtvedt C, Nissen-Meyer J, Sletten K, Nes I (1991). Purification and amino acid sequence of lactocin S, a bacteriocin produced by Lactobacillus sake L45. Applied and Environmental Microbiology 57(6):1829-1834. |
|
|
Ni K, Wang Y, Li D, Cai Y, Pang H (2015). Characterization, identification and application of lactic acid bacteria isolated from forage paddy rice silage PloS one 10(3):e0121967. |
|
|
Nigatu JM, Tuji FA, Tefera AT (2015). Evaluation of the antagonistic effect of six mixed cultures of lactic acid bacteria, isolated from the Ethiopian fermented milk ergo, against some foodborne pathogens inoculated into the Ethiopian cottage cheese ayib. African Journal of Microbiology Research 9(29):1789-1797. |
|
|
Noor R, Islam Z, Munshi SK, Rahman F (2013). Influence of temperature on Escherichia coli growth in different culture media. Journal of Pure and Applied Microbiology 7(2): 899-904. |
|
|
Ogodo A, Ugbogu O, Onyeagba R, Orji F (2017). Dynamics of functional properties of sorghum flours fermented with lactic acid bacteria (LAB)-consortium isolated from cereals. Food Research Journal 24(6):2666-2671. |
|
|
Ogunbanwo S, Sanni A, Onilude A (2003). Characterization of bacteriocin produced by Lactobacillus plantarum F1 and Lactobacillus brevis OG1. African Journal of Biotechnology 2(8):219-227. |
|
|
Onwuakor C, Nwaugo V, Nnadi , Emetole J (2014). Effect of varied culture conditions on crude supernatant (bacteriocin) production from four Lactobacillus species isolated from locally fermented maize (ogi) . American Journal of Microbiological Research 2(5):125-130. |
|
|
Ouali FA, Al Kassaa I, Cudennec B, Abdallah M, Bendali F, Sadoun D, Chihib NE, Drider D (2014). Identification of lactobacilli with inhibitory effect on biofilm formation by pathogenic bacteria on stainless steel surfaces. International Journal of Food Microbiology 191:116-124. |
|
|
Oyedeji O, Ogunbanwo ST, Onilude AA (2013). Predominant lactic acid bacteria involved in the traditional fermentation of fufu and ogi, two Nigerian fermented food products. Food and Nutrition Sciences 4(11):40. |
|
|
Perez RH, Zendo T, Sonomoto K (2014). Novel bacteriocins from lactic acid bacteria (LAB): various structures and applications. Microbial Cell Factories 13(1):S3. |
|
|
Pham D, Ansquer D, Chevalier A, Dauga C, Peyramale A, Wabete N, Labreuche Y (2014). Selection and characterization of potential probiotic bacteria for Litopenaeus stylirostris shrimp hatcheries in New Caledonia. Aquaculture 432:475-482. |
|
|
Piard JC, Muriana P, Desmazeaud M, Klaenhammer T (1992). Purification and partial characterization of lacticin 481, a lanthionine-containing bacteriocin produced by Lactococcus lactis subsp. lactis CNRZ 481. Applied and Environmental Microbiology 58(1):279-284. |
|
|
Piper P, Calderon CO, Hatzixanthis K, Mollapour M (2001). Weak acid adaptation: the stress response that confers yeasts with resistance to organic acid food preservatives. Microbiology 147(10):2635-42. |
|
|
Pratush A, Gupta A, Kumar A, Vyas G (2012). Application of purified bacteriocin produced by Lactococcus lactis AP2 as food biopreservative in acidic foods. Annals, Food Science Technology 13:82-87. |
|
|
Qiuju W, Yizhe C, Shengjun L, Ruijin Z (2013). Isolation and identification of lactic acid bacteria in duodenum of laying hens fed in cage. Journal of Heilongjiang Bayi Agricultural University 3:8. |
|
|
Ramu R, Shirahatti PS, Devi AT, Prasad A (2015). Bacteriocins and their applications in food preservation. Critical Reviews in Food Science and Nutrition 00-00. |
|
|
Rao KP, Chennappa G, Suraj U, Nagaraja H, Raj AC, Sreenivasa M (2015). Probiotic potential of Lactobacillus strains isolated from sorghum-based traditional fermented food. Probiotics and Antimicrobial Proteins 7(2):146-156. |
|
|
Rather IA, Galope R, Bajpai VK, Lim J, Paek WK, Park YH (2017). Diversity of marine bacteria and their bacteriocins: applications in aquaculture. Reviews in Fisheries Science and Aquaculture 25(4):257-269. |
|
|
Rattanachaikunsopon P, Phumkhachorn P (2010). Lactic acid bacteria: their antimicrobial compounds and their uses in food production Annals of Biological Research 1(4):218-228. |
|
|
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951). Protein measurement with the Folin phenol reagent. Journal of Biological Chemistry 193:265-275. |
|
|
Reis JA, Paula AT, Casarotti SN, Penna ALB (2012). Lactic acid bacteria antimicrobial compounds: characteristics and applications. Food Engineering Reviews 4(2):124-140. |
|
|
Ringø E , Gatesoupe FJ (1998). Lactic acid bacteria in fish: a review. Aquaculture 160(3-4):177-203. |
|
|
Ringø E, Hoseinifar SH, Ghosh K, Doan HV, Beck BR, Song SK (2018). Lactic acid bacteria in finfish-An update. Frontiers in Microbiology 9:1818. |
|
|
Sankar NR, Priyanka VD, Reddy PS, Rajanikanth P, Kumar VK, Indira M (2012). Purification and characterization of bacteriocin produced by Lactobacillus plantarum isolated from cow milk. International Journal of Microbiology Research 3(2):133-137. |
|
|
Sarika A, Lipton A, Aishwarya M (2010). Bacteriocin production by a new isolate of Lactobacillus rhamnosus GP1 under different culture conditions. Advance Journal of Food Science and Technololgy 2(5):291-297. |
|
|
Sarika A, Lipton AP, Aishwarya M (2019). Biopreservative efficacy of bacteriocin GP1 of Lactobacillus rhamnosus GP1 on stored fish filets. Frontiers in Nutrition 6:29. |
|
|
Shahid M, Hussain B, Riaz D, Khurshid M, Ismail M, Tariq M (2017). Identification and partial characterization of potential probiotic lactic acid bacteria in freshwater Labeo rohita and Cirrhinus mrigala. Aquaculture Research 48(4):1688-1698. |
|
|
Shahidi F (2015). Handbook of antioxidants for food preservation: Woodhead Publishing. |
|
|
Sharma S (2015). Food preservatives and their harmful effects. International Journal of Scientific and Research Publications 5(4):1-2. |
|
|
Sica MG, Brugnoni LI, Marucci PL, Cubitto MA (2012). Characterization of probiotic properties of lactic acid bacteria isolated from an estuarine environment for application in rainbow trout (Oncorhynchus mykiss, Walbaum) farming. Antonie van Leeuwenhoek 101(4):869-879. |
|
|
Silva CC, Silva SP, Ribeiro SC (2018). Application of bacteriocins and protective cultures in dairy food preservation. Frontiers in Microbiology 9:594. |
|
|
Smid E, Kleerebezem M (2014). Production of aroma compounds in lactic fermentations. Annual Review of Food Science and Technology 5:313-326. |
|
|
Stewart CM (2003). Staphylococcus aureus and staphylococcal enterotoxins. Ch 12 In: Hocking AD (ed) Foodborne microorganisms of public health significance. 6th ed, Australian Institute of Food Science and Technology (NSW Branch) Sydney pp. 359-380. |
|
|
Subramanyam MN (2020). Molecular characterisation of probiotic Lactobacillus fermentum isolated from home made curd Journal of Microbiology, Biotechnology and Food Sciences 9(4):848-855. |
|
|
Trz?skowska M, Ko?o?yn-Krajewska D, WójciakK, Dolatowski Z (2014). Microbiological quality of raw-fermented sausages with Lactobacillus casei LOCK 0900 probiotic strain. Food Control 35(1):184-191. |
|
|
Tserovska L, Stefanova S, Yordanova T (2002). Identification of lactic acid bacteria isolated from Katyk, goat's milk and Cheese. Journal of Culture Collections 3:48-52. |
|
|
Tufail M, Hussain S, Malik F, Mirza T, Parveen G, Shafaat S, Sadiq A (2011). Isolation and evaluation of antibacterial activity of bacteriocin produced by Lactobacillus bulgaricus from yogurt. African Journal of Microbiology Research 5(22):3842-3847. |
|
|
Udhayashree N, Senbagam D, Senthilkumar B, Nithya K, Gurusamy R (2012). Production of bacteriocin and their application in food products Asian Pacific Journal of Tropical Biomedicine 2(1):S406-S410. |
|
|
Van Sinderen D, Crowley S (2013). Lactobacillus plantarum species possessing broad spectrum anti-fungal activity and exhibiting high heat tolerance and osmotolerance: Google Patents. |
|
|
Vignolo G, Saavedra L, Sesma F, Raya R (2012). 22 Food bioprotection: lactic acid bacteria as natural preservatives. Progress in Food Preservation P 453. |
|
|
Wang CY, Lin PR, Ng CC, Shyu YT (2010). Probiotic properties of Lactobacillus strains isolated from the feces of breast-fed infants and Taiwanese pickled cabbage. Anaerobe 16(6):578-585. |
|
|
Woraprayote W, Malila Y, Sorapukdee S, Swetwiwathana A, Benjakul S, Visessanguan W (2016). Bacteriocins from lactic acid bacteria and their applications in meat and meat products. Meat Science 120:118-132. |
|
|
Yang E, Fan L, Jiang Y, Doucette C, Fillmore S (2012). Antimicrobial activity of bacteriocin-producing lactic acid bacteria isolated from cheeses and yogurts. AMB Express 2(1):48. |
|
|
Yang SC, Lin CH, Sung CT, Fang JY (2014). Antibacterial activities of bacteriocins: application in foods and pharmaceuticals. Frontiers in Microbiology 5:241. |
|
|
Yousef AE, Balasubramaniam V (2013). Physical methods of food preservation. Food Microbiology American Society of Microbiology pp. 737-763. |
|
|
Zorriehzahra MJ, Delshad ST, Adel M, Tiwari R, Karthik K, Dhama K, Lazado CC (2016). Probiotics as beneficial microbes in aquaculture: an update on their multiple modes of action: a review. Veterinary Quarterly 36(4):228-241. |
|
|
Zou Y, Liu F, Fang C, Wan D, Yang R, Su Q, Zhao J (2013). Lactobacillus shenzhenensis sp. nov., isolated from a fermented dairy beverage. International Journal of Systematic and Evolutionary Microbiology 63(5):1817-1823. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


