Full Length Research Paper

ABSTRACT
INTRODUCTION
MATERIALS AND METHODS
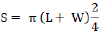
RESULTS
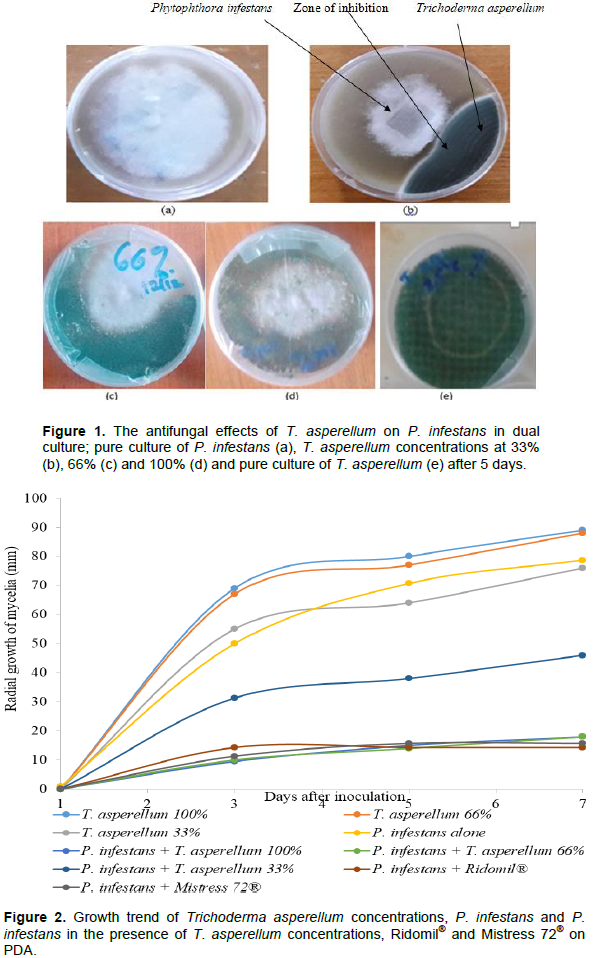
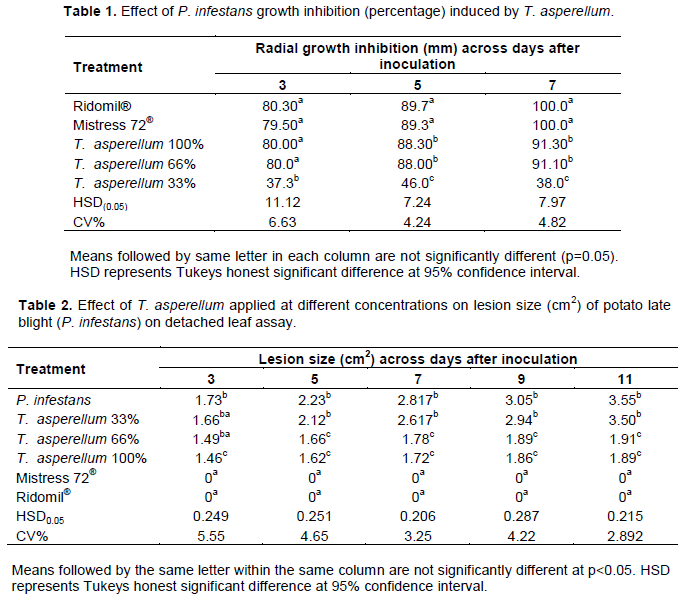

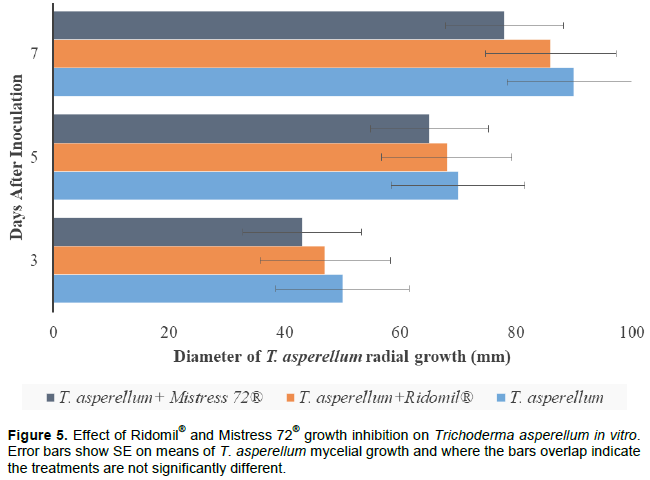
DISCUSSION
CONCLUSION
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
REFERENCES
|
Agbeniyi SO, Adedeji A R, Adeniyi DO (2014). On-Farm Evaluation of Trichoderma asperellum on the Suppression of Phytophthora megakarya Causing Pod Rot of Theobromae cacao in Nigeria. British Journal of Applied Science and Technology 4(22):3153-3159. |
|
|
Amin F, Razdan VK, Mohiddin FA, Bhat KA, Sheikh P A (2010). Effect of volatile metabolites of Trichoderma species against seven fungal plant. Journal of Phytology 2(10):34-37. |
|
|
An Y, Kang S, Kim K, Kook B, Jeun Y (2010). Enhanced defense responses of tomato plants against late blight pathogen Phytophthora infestans by pre-inoculation with rhizobacteria. Crop Protection 29(12):1406-1412. |
|
|
Aparecida M, Moura KE, Moura KE, Salomão D, Patricio RA. (2018). Compatibility of Trichoderma isolates with pesticides used in lettuce crop. Summa Phytopathologica 2(28):137-142. |
|
|
Azimuddin MD, Alam QM, Baset MA (2009). Potato for food security in Bangladesh. International Journal of Sustainable Crop Production 4(1):94-99. |
|
|
Bahramisharif A, Rose LE (2018). Efficacy of biological agents and compost on growth and resistance of tomatoes to late blight. Planta 249(3):799-813. |
|
|
Carrero-carr I, Monte E, Hermosa R, Jim RM (2016). Trichoderma asperellum is effective for biocontrol of Verticillium wilt in olive caused by the defoliating pathotype of Verticillium dahliae. Phytopathology 88:45-52. |
|
|
Childers R, Danies G, Myers K, Fei Z, Small IM, Fry WE (2014). Acquired resistance to mefenoxam in sensitive isolates of Phytophthora infestans. Phytopathology 105(3):342-349. |
|
|
Chowdappa P, Kumar S P, Lakshmi MJ, Upreti KK (2013). Growth stimulation and induction of systemic resistance in tomato against early and late blight by Bacillus subtilis OTPB1 or Trichoderma harzianum OTPB3. Biological Control 65(1):109-117. |
|
|
Cooke DE, Cano LM, Raffaele S, Bain RA, Cooke LR, Etherington GJ, Kamoun S (2012). Genome analyses of an aggressive and invasive lineage of the irish potato famine pathogen. PLoS Pathogens 8(10):223-254. |
|
|
Cooke LR, Schepers HT, Hermansen A, Bain RA, Bradshaw NJ, Ritchie F, Nielsen BJ (2011). Epidemiology and integrated control of potato late blight in Europe. Potato Research 54(2):183-222. |
|
|
Ephrem ZD, Santhanam A, Gorfu, D, Kassa B (2011). Biocontrol activity of Trichoderma viride and Pseudomonas fluorescens against Phytophthora infestans under greenhouse conditions. Journal of Agricultural Technology 7(6):1589-1602. |
|
|
FAOSTAT (2018). Statistics for global food production. Food Agricultural statistics, United Nations, Peru. https://www.potatopro.com/world/potato-statistics. Accessed 14th May 2019. |
|
|
Fontem DA, Olanya OM, Tsopmbeng GR, Owona MA (2005). Pathogenicity and metalaxyl sensitivity of Phytophthora infestans isolates obtained from garden huckleberry, potato and tomato in Cameroon. Crop Protection 24(5):449-456. |
|
|
Fatima K, Noureddine K, Henni JE, Mabrouk K (2015). Antagonistic effect of Trichoderma harzianum against Phytophthora infestans in the North-west of Algeria. International Journal of Agronomy and Agricultural Research 6(4):44-53. |
|
|
Forbes GA (1997). Manual for Laboratory Work on Phytophthora infestans CIP Training Manual. International Potato Center (CIP), Lima Peru. |
|
|
Gigot JA, Gundersen B, Inglis DA (2009). Colonization and sporulation of Phytophthora infestans on volunteer potatoes under western Washington conditions. American Journal of Potato Research 86(1):1-14. |
|
|
Goufo P, Teugwa MC, Fontem DA, Ngnokam D (2008). High efficacy of extracts of Cameroon plants against tomato late blight disease. Agronomy for Sustainable Development 26(4):567-573. |
|
|
Goufo P, Fontem DA, Ngnokam D (2017). Evaluation of plant extractsfor tomato late blight control in Cameroon.Journal of Agronomy and Sustainable Development 28:567-573. |
|
|
Haverkort AJ, Boonekamp PM, Hutten R, Jacobsen E, Lotz LA, Kessel GJ, Van Der Vossen EG (2008). Societal costs of late blight in potato and prospects of durable resistance through cisgenic modification. Potato Research 51(1):47-57. |
|
|
Istv S (2014). Potential of Trichoderma species and nematode-trapping fungi to control plant-parasitic nematodes : in vitro confrontation and gene expression assays using Caenorhabditis elegans model system. PhD Thesis Márton Szabó G ödöllÅ‘ University, Hungary. |
|
|
Itachi TM, Anamori MK, Eraoka TT, Rie TA (2007). Mode of action of Trichoderma asperellum SKT-1, a biocontrol agent against Gibberella fujikuroi. Journal of Pesticide Science 32(3):222-228. |
|
|
Jaetzold R, Schmidt H, Hornetz B, Shisanya C (2006). Farm management handbook of Kenya-Natural conditions and farm management information, 2nd Edition. Volume II/B. Ministry of Agriculture, Nairobi. |
|
|
Johnson DA, Cummings TF (2009). Latent infection of potato seed tubers by Phytophthora infestans during long-term cold storage. Plant Disease 93(9):940-946. |
|
|
Johnson DA, Cummings TF (2013). A plant stem inoculation assay for assessing transmission of Phytophthora infestans from potato seed tubers to emerged shoots. Plant Disease 97(2):183-188. |
|
|
Kabir L, Sang WK, Yun SK (2013). Biocontrol of late blight and plant growth promotion in tomato using rhizobacterial isolates. Journal of Microbiology and Biotechnology 23(7):885-892. |
|
|
Kipngeno P, Losenge T, Maina N, Kahangi E, Juma P (2015). Efficacy of Bacillus subtilis and Trichoderma asperellum against Pythium aphanidermatum in tomatoes. Biological Control 90:92-95. |
|
|
Lal M, Yadav S, Sharma S, Singh BP, Kaushik SK (2017). Integrated management of late blight of potato. Vegetos 26(2):362-367. |
|
|
Leonetti P, Chiara M, Sergio Z, Claudio M (2017). Induction of SA-signaling pathway and ethylene biosynthesis in Trichoderma harzianum- treated tomato plants after infection of the root-knot nematode Meloidogyne incognita. Plant Cell Reports 36(4):621-631. |
|
|
Mariita MO, Nyangeri J, Makatiani JK (2016). Assessing the Incidences of late blight disease on irish potato varieties in Kisii County , Kenya. Annual Research and Review in Biology 9(6):1-8. |
|
|
Matson ME, Small IM, Fry WE, Judelson HS (2015). Metalaxyl resistance in Phytophthora infestans: Assessing role of rpa190 gene and diversity within clonal lineages. Phytopathology 105(12):1594-1600. |
|
|
Miles LA, Lopera CA., Gonza S, Franco AE, Restrepo S (2012). Exploring the biocontrol potential of fungal endophytes from an Andean Colombian Paramo ecosystem. BioControl 57(5):697-710. |
|
|
Muchiri FN, Narla RD, Olanya OM, Nyankanga RO, Ariga ES (2017). Efficacy of fungicide mixtures for the management of Phytophthora infestans ( US-1 ) on potato. Phytoprotection 90:19-29. |
|
|
Muthoni J, Shimelis H, Melis R (2013). Potato production in Kenya: Farming systems and production constraints. Journal of Agricultural Science 5(5):182-197. |
|
|
Naglot A, Goswami S, Rahman I, Shrimali DD, Yadav KK, Gupta VK, Veer V (2015). Antagonistic potential of native Trichoderma viride strain against potent tea fungal pathogens in north east India. Plant Pathology Journal 31(3):278-289. |
|
|
Njoroge AW, Andersson B, Yuen JE, Forbes GA (2019). Greater aggressiveness in the 2_A1 lineage of Phytophthora infestans may partially explain its rapid displacement of the US-1 lineage in east Africa. Plant Pathology 68(3):566-575. |
|
|
Nowicki M, Majeed F (2012). Potato and tomato late blight caused by Phytophthora infestans :An overview of pathology and resistance breeding. Plant Disease 96(1):4-17. |
|
|
Nyankanga RO, Wien HC, Olanya OM, Ojiambo PS (2007). Farmers ' cultural practices and management of potato late blight in Kenya Highlands: implications for development of integrated disease management. International Journal of Pest Management 50(2):135-144. |
|
|
Okello JJ, Zhou Y, Kwikiriza N, Ogutu S, Barker I, Schulte-Geldermann E, Ahmed JT (2017). Productivity and food security effects of using of certified seed potato: The case of Kenya's potato farmers. Agriculture and Food Security 6(1):342-350. |
|
|
Patel S, Saraf M (2017). Biocontrol efficacy of Trichoderma asperellum MSST against tomato wilting by Fusarium. Archives Phytopathology and Plant Protection 5408:228-238. |
|
|
Rhouma A, Salem I, Gomez DG (2016). Efficacy of two fungicides for the management of Phytophthora infestans on potato through different applications methods adopted in controlled conditions. International Journal of Applied Science and Agriculture 2:39-45. |
|
|
Runno-Paurson E, Hannukkala AO, Kotkas K, Koppel M, Williams IH, Mänd M (2013). Impact of phytosanitary quality of seed potato and temporal epidemic progress on the phenotypic diversity of Phytophthora infestans populations. American Journal of Potato Research 90(3):245-254. |
|
|
Saravanakumar K, Fan L, Fu K, Yu C, Wang M (2016). Cellulase from Trichoderma harzianum interacts with roots and triggers induced systemic resistance to foliar disease in maize. Nature Publishing Group 6:1-18. |
|
|
Schuster A, Schmoll M (2010). Biology and biotechnology of Tricho-derma. Application Microbiology Biotechonology 87(1):787-799. |
|
|
Scott G, Labarta R, Suarez V (2010). Booms, bursts and emerging markets for potatoes and potato products in east and central Africa 1961-2010. Potato Research 56(3):205-236 |
|
|
Sharma P (2011). Complexity of Trichoderma-Fusarium interaction and manifestation of biological control. Australian Journal of Crop Science 5(8):1027-1038. |
|
|
Sharma P, Saikia MK (2013). Management of late blight of potato through chemicals. Journal of Agriculture and Veterinary Science 2(2):23-26. |
|
|
Sharma V, Salwan R, Sharma PN, Kanwar SS (2017). Elucidation of biocontrol mechanisms of Trichoderma harzianum against different plant fungal pathogens: Universal yet host specific response. International Journal of Biological Macromolecules 95(1):72-79. |
|
|
Were HK, Kabira JN, Kinyua ZM, Olubayo FM, Karinga JK, Aura J, Torrance L (2014). Occurrence and distribution of potato pests and diseases in Kenya. Potato Research 56(3):325-342. |
|
|
Widmer TL (2014). Screening Trichoderma species for biological control activity against Phytophthora ramorum in soil. Biological Control 79(3):43-48. |
|
|
Wu Q, Sun R, Ni M, Yu J, Li Y, Yu C (2017). Identification of a novel fungus, Trichoderma asperellum GDFS1009, and comprehensive evaluation of its biocontrol efficacy. PLoS ONE 12(6):1-20. |
|
|
Xu X, Jeffries P, Pautasso M, Jeger MJ (2011). Combined use of biocontrol agents to manage plant diseases in theory and practice. Phytopathology 101:1024-1031. |
|
|
Yao Y, Li Y, Chen Z, Zheng B, Zhang L, Niu B, Wang Q (2016). Biological control of potato late blight using isolates of Trichoderma. American Journal of Potato Research 93(1):33-42. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


