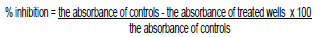ABSTRACT
Acinetobacter baumannii is an opportunistic pathogen which is a major cause of nosocomial infection. Bacteriophages are bacterial viruses that are used as alternative agents in the treatment of multidrug-resistant bacterial infections. In this research, our purpose was to investigate the efficacy of a cocktail of five bacteriophages. Ten (10) bacteriophage isolates were determined for the host range analysis and five bacteriophages, ØABP-02, ØABP-19, ØABP-29, ØABP-39 and ØABP-44 that showed broad host range (36.9-64.6 %) were selected for the preparation of the bacteriophage cocktail. Transmission electron microscopy revealed ØABP-02 and ØABP-44 belonged to family Myoviridae and ØABP-19, ØABP-29, ØABP-39 belonged to family Podoviridae. The bacteriophage cocktail was tested for its efficacy on growth inhibition against 44 A. baumannii clinical isolates using the colorimetric microtiter plate method. The results of the growth inhibition assay of the bacteriophage cocktail showed that the growth inhibition against A. baumannii ranged from 45.1- 96%. High efficacy of the bacteriophage cocktail was found against the A. baumannii strains that can be infected by the five bacteriophages (>77%). Our study demonstrates high efficacy of the bacteriophage cocktail on inhibiting the growth of A. baumannii. The bacteriophage cocktail is a valuable alternative agent for controlling the multi- drug resistant A. baumannii in hospitals.
Key words: Bacteriophage, Acinetobacter baumannii, growth inhibition, host range.
A. baumannii are opportunistic pathogens that cause a variety of infections such as respiratory tract infections, urinary tract infections, skin infections and bacteremia. A. baumannii have developed antibiotic resistance which is a serious problem in the treatment of these infections. The infections of multi-drug resistant A. baumannii (MDR-
AB) and extensively drug-resistant A. baumannii (XDR-AB) are of particular concern in patients in intensive care and burns units which cause high mortality rates (Alp et al., 2012; Park et al., 2013). In recent decades, the prevalence of MDR-AB and XDR-AB has increased worldwide. MDR-AB and XDR-AB infected patients must stay longer in hospital than patients without such drug resistant infections. This leads to a higher cost of hospitalization. Bacteriophage therapy has been claimed to be a potential candidate for the treatment of MDR-AB and XDR-AB. Bacteriophages are viruses that specifically infect bacteria and kill the host cell when the progeny of the phage particles leave the cells. So far, more than 10 A. baumannii bacteriophages have been isolated and investigated for their biological properties (Chang et al., 2012; Jin et al., 2012; Kitti et al., 2014; Lee et al., 2011; Lin et al., 2010; Merabishvili et al., 2014; Popova et al., 2012; Shen et al., 2012; Thawal et al., 2012; Yang et al., 2010; Yele et al., 2012). These bacteriophages have been examined for being potential candidates in phage therapy. The host range of bacteriophages is significant for use as antibacterial agents. However, narrow host specificity of A. baumannii bacteriophages, ranging from 13-59%, have been reported from different geographical isolates (Jin et al., 2012; Merabishvili et al., 2014; Thawal et al., 2012). Consequently, to assure that bacteriophages possess a broad host range activity that includes the target bacteria, multiple phage types possessing a diversity of host ranges are often combined into mixtures called ‘phage cocktails’ (Chan et al., 2013). Phage cocktails have been applied to improve the chances of success in bacterial killing in the treatment of Pseudomonas aeruginosa and E. coli O157:H7 (Chan et al., 2013). Due to the narrow host range of A. baumannii bacteriophages, the objective of our study was to screen new bacteriophages with broad host range activity and determine the efficacy of a A. baumannii bacteriophage cocktail in inhibiting the growth of A. baumannii.
Bacterial strains
Eleven MDR-AB isolates obtained from Buddhachinaraj hospital, Phitsanulok, Thailand were used as host bacteria for bacteriophage isolation (Niumsup et al., 2009). Sixty-five clinical isolates of A. baumannii obtained from Sawan Pracharak Hospital, Nakorn Sawan, Thailand were used for growth inhibition assay. Bacteria were cultivated in Luria - Bertani broth (LB) or Luria - Bertani Agar (LBA). Confirmation of the Acinetobacter species was based on biochemical tests and detection of the 16S rRNA gene.
Isolation of bacteriophage
All bacteriophages were isolated from wastewater treatment plants from two hospitals in Phitsanulok Province. Samples were collected and centrifuged to remove debris. The supernatant was filtered. Then, 5 ml of the filtered supernatants were mixed with 5 ml of double strength broth containing overnight culture A.baumannii. After 48 h, the culture was centrifuged, and the supernatant was used for the detection of lytic bacteriophages by a double-layer method. Phage enrichment and puriï¬cation were performed as described by Kitti et al. (2014).
Host range analysis
Host range analysis of the ten bacteriophages was determined by spot tests. Hundred microliter of overnight bacterial cultures were added to 2.5 ml of soft agar, mixed gently and poured into an agar plate. Subsequently, 5 µl aliquots of phage suspension (1.0 x 108 PFU) were spotted on the lawn of bacteria. Plates were dried and incubated at 37°C for 7 h. The clearance zone indicating lysis at the spot of phage inoculation implied that the host was sensitive.
Morphology of bacteriophages
Five bacteriophages which showed high lytic activity were used to determine the morphology. Phage suspension (1012 PFU/ml) was dropped onto the surface of a formvar-coated grid and negatively stained with 0.5% uranyl acetate for 3-5 min. After drying, the preparations were observed in a transmission electron microscope (Philips, Oregon, USA).
Bacteriohage enrichment and preparation of bacteriophage cocktail
A. baumannii were grown in 100 ml LB broth until OD600 reached 0.4. Phages were added at a multiplicity of infection (MOI) of 0.5 and incubated at 37°C until complete lysis. Then, 2-3 ml chloroform was added and bacterial debris was pelleted by centrifugation at 4000 g for 10 min. Each phage was enriched three times and the phage concentration was determined using the plaque assay method (Kitti et al., 2014). The phage stocks were stored at 4 ± 1°C. Bacteriohage titer was determined before a growth inhibition assay was performed. The titer of each bacteriophage was determined and diluted into 106 PFU/ml in SM buffer. Each bacteriophage was mixed (1:1:1:1:1) to prepare the bacteriophage cocktail.
Growth inhibition assay
A bacterial growth inhibition assay was performed as described by Knezevic and Petrovic (2008) using the cocktail of five A. baumannii bacteriophages (Knezevic and Petrovic, 2014). Briefly, A. baumannii were cultured in LB broth incubated at 37°C with shaking at 150 rpm / min for 24 h.
The overnight cultures were diluted (1:100) into LB broth and incubated at 37°C with shaking at 150 rpm/ min until OD650 reached 0.4 then 50 ul (approximately 2 × 108 CFU/ml) were inoculated in 50 ul double strength LB broth into 96-well microtiter plates wells flat bottom (Nunc,USA). One hundred microliters of the five bacteriophage cocktail was added to each well. Then 50 ul of 0.1% filter sterilized TTC (Hi-media) was added. The plate was incubated at 37°C in the dark for 24 h. The absorbance at OD540 was measured by a Micro-plate Reader. The experiment was replicated twice with triplicate samples. The percentage inhibition of the bacteriophage cocktail against all A. baumannii was calculated using the formula:
Isolation of lytic phage
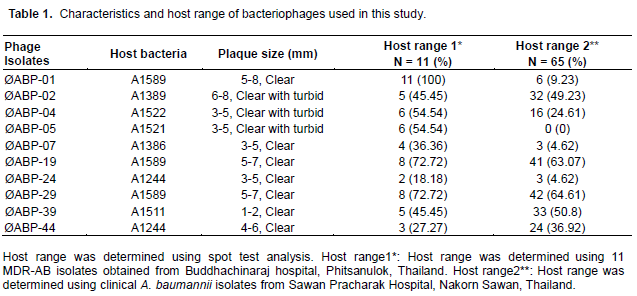
Fifty-four isolates of A. baumannii bacteriophages were collected from two treatment plants. Ten isolates with high lytic activity on a broad range of 11 A. baumannii were selected for further characterization. These are designated as ØABP-01, ØABP-02, ØABP-04, ØABP-05, ØABP-07, ØABP-19, ØABP-24, ØABP-29, ØABP-39 and ØABP-44 (Table 1).
Host range analysis
Ten (10) A. baumannii bacteriophages isolated from hospitals in Phitsanulok Province were used to investigate the ability to infect 65 A. baumannii isolated from a hospital in Nakorn Sawan Province. Five bacteriophages that showed a broad host range were ØABP-02(49.23%), ØABP- 19 (63.07%), ØABP-29(64.61%), ØABP-39(50.8 %) and ØABP-44(36.9 %) (Table 1). Five that showed low specific host ranged from 0-25.75% were ØABP-01, ØABP-04, ØABP-05, ØABP- 07 and ØABP-24 (Table 1). ØABP-02, ØABP-19, ØABP-29, ØABP-39 and ØABP-44 were selected in this study for the preparation of the bacteriophage cocktail.
Bacteriophage susceptibility type of A. baumannii
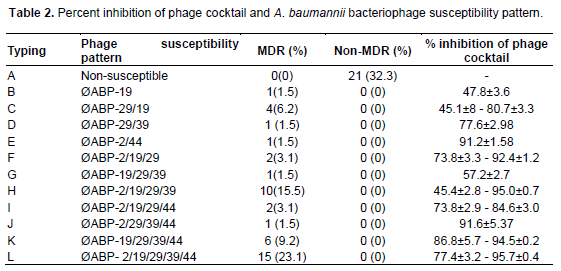
We classified 65 A. baumannii by spot tests using five bacteriophages, into 12 groups; A-L (Table 2).
Among the bacterial strains tested, 21 were bacteriophage non-susceptible strains (group A) and were also non multi-drug resistant strains. Fourteen A. baumannii were susceptible to infection by 5 bacteriophages. All of them were multi-drug resistant bacteria and belonged to group L (Table 2).
Phage morphology
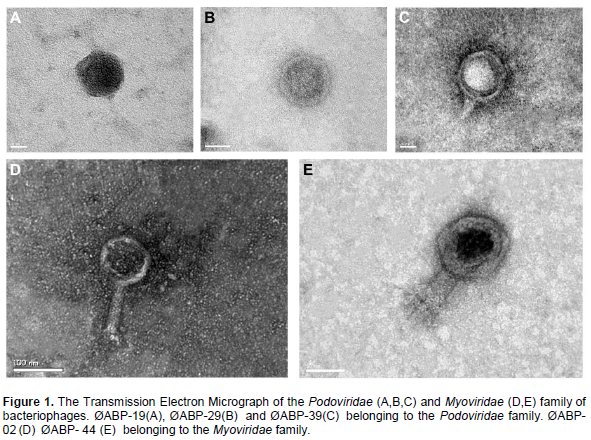
To classify ØABP-02, ØABP-19, ØABP-29, ØABP-39 and ØABP-44 into morphology specific groups, phage particles were examined with an electron microscope. Our data shows ØABP-19, ØABP-29and ØABP-39 have an icosahedral head (70,110, 70 nm) with a short tail, belonging to the Podoviridae family (Figure 1a, b and c). ØABP-02 and ØABP-44 showed a contractile tail (60, 11 nm) and an icosahedral head (80, 44 nm) (Figure 1d and e), a morphology characteristic of the family Myoviridae. All five bacteriophages were tailed bacteriophages and identified as members of order Caudovirales.
Effect of bacteriophage cocktail on A. baumannii growth
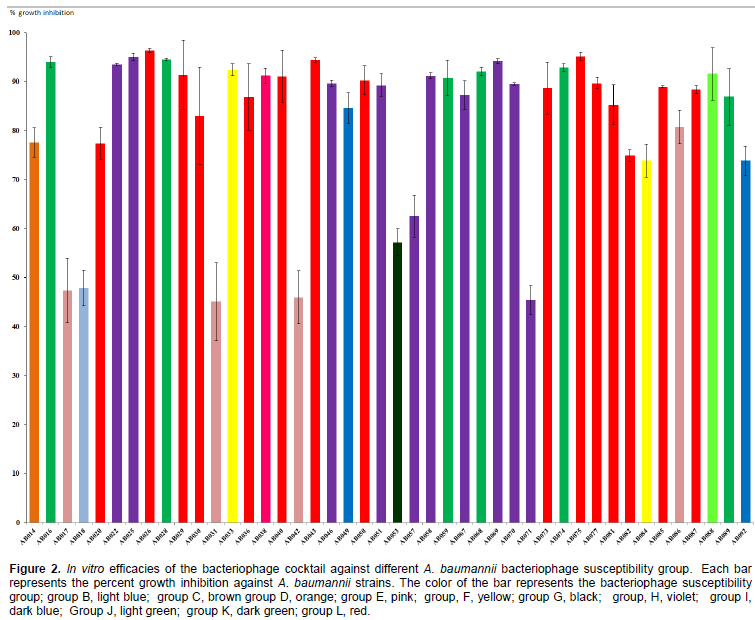
We used the colorimetric microtiter plate method to determine the efficacy of the five bacteriophages in inhibiting A. baumannii growth. The ratio of bacteriophage to infected target host was preliminarily determined using one bacteriophage. We found that the amount of bacteriophage and A. baumannii that showed good inhibition effects was 106 PFU/ml and 108 CFU/ml (data not shown). The effects of the bacteriophage cocktail on bacterial growth was tested using 44 A. baumannii. The results of the growth inhibition assay using the bacteriophage cocktail are presented in Table 2 and Figure 2. The bacteriophage cocktail exhibiting the growth inhibition of A. baumannii ranged from 45.1-96% (Figure 2). A. baumannii that can be infected with the five bacteriophages (group L) showed the percentage growth inhibition higher than 77% (Figure 2). No growth inhibition was found in the non MDR-AB strains which are bacteriophage non susceptible strains (group A).
The bacteria–phage interactions, host range and dosage are important information to the successful development of phages for therapy (Levin and Bull, 2004; Hyman and Abedon, 2010).
In this study, we determined the host range of ten bacteriophages to identify five potential bacteriophages as candidates for developing phage cocktails. Our results classified 65 A. baumannii into 12 bacteriophage susceptibility groups (Table 2). The phage-susceptibility test provides a specific method for differentiating the MDR-AB from non MDR-AB. A. baumannii strains in group L (14 isolates) were infected with the five bacteriophages, whereas A. baumannii strains in group A (21 isolates) were non-bacteriophage susceptible strains. All of the strains in group A are non-MDR-AB. Bacteria have evolved several adaptive immunity mechanisms to resist bacteriophage infection. These include adsorption resistance which results in reduced interaction between phage and bacterium (Hyman and Abedon, 2010). The variety of receptor sites on the bacterial surface are involved in phage adsorption and penetration into the host cell (Rakhuba et al., 2013). In addition, the differences in the cell wall polysaccharide structure receptor on the bacterial surface are major factors in bacteriophage sensitivity (Ainsworth et al., 2014). The diversification of restriction–modification (RM) systems include phage-genome uptake blocks, superinfection immunity, restriction modification, and the generation of anti-phage sequences in CRISPR loci are also immunity mechanisms to resist bacteriophage infection after phage adsorption (Hyman and Abedon, 2010; Bikard et al., 2013). A previous study showed that ØABP-01 exhibited high lytic activity and a broad host range (100%) when using A. baumannii host isolated from the same hospital as the bacteriophage isolated (Kitti et al., 2014). Our data indicates that ØABP-01 showed narrow host range activity (9.23%) against A. baumannii host isolated from a different hospital as the bacteriophage isolate. This can be explained by the difference in the immunity mechanism of bacteriophage infection resistance among geographic differences A. baumannii clones.
Bacteriophages that have a broad host range are valuable candidates for further study of the efficacy of A. baumannii bacteriophage cocktail. Among the five bacteriophages in our study, two families of bacteriophages belonging to Podoviridae and Myoviridae were observed under an electron microscope. This reflects similar previous findings that most of the A. baumannii bacteriophages identified so far belong to Podoviridae and Myoviridae (Jin et al., 2012; Lee et al., 2011; Lin et al., 2010; Popova et al., 2012; Thawal et al., 2012). They are classified in Caudovirales which includes 96% of bacteriophages isolates identified to date (Ackermann, 2009).
Most studies in A. baumannii bacteriophages assessing the efï¬cacy of bacteriophages used one bacteriophage at a time. Our studies used the colorimetric microtiter plate method to determine the efficacy of bacteriophage cocktail in bacterial growth inhibition. This test is economically cheaper and less time consuming than the conventional method. The different MOI of bacteria and bacteriophages are involved in the efficacy of inhibition in this test. We found the MOI 0.01 of bacteriophages and A. baumannii showed good inhibition effects. This MOI was used in the growth inhibition system. The bacteriophage cocktail showed high percent growth inhibition (> 70%) against the A. baumannii groups I, J, K and L. These strains are susceptible to at least 4 or 5 bacteriophages. However, a low percent of growth inhibition was found against some of the groups of A. baumannii (B, C, G and H). The occurrence of bacteriophage resistant mutant isolates may explain this phenomenon. Mutation can occur during overnight incubation of bacteria with bacteriophages in a microplate system (Fridholm et al., 2005). Isolation of bacteriophages that infected the resistant mutant isolates are needed and further investigated for fulfilling the limitation of bacteriophages in the treatment of MDR-AB.
In conclusion, this study showed the high efficacy of the bacteriophage cocktail against most phage susceptible A. baumannii. All of them were multi-drug resistant bacteria. The bacteriophage cocktail was found to be potential agent against multi-drug resistance bacteria and have some future important in phage therapy.
The authors did not declare any conflict of interest.
We thank Dr. Pannika Ritvirool and staff from Sawan Pracharak hospital for providing A. baumannii clinical specimens. We are grateful to Mr. Roy Morien from Naresuan University language center for editing this manuscript. This work was supported by the grant from Office of Research Council of Thailand 2014 (R2557B004).
REFERENCES
|
Ackermann HW (2009). Phage classification and characterization. Methods Mol. Biol. 501:127-140.
Crossref
|
|
|
|
Ainsworth S, Sadovskaya I, Vinogradov E, Courtin P, Guerardel Y, Mahony J, Grard T, Cambillau C, Chapot-Chartier MP, van Sinderen D (2014). Differences in lactococcal cell wall polysaccharide structure are major determining factors in bacteriophage sensitivity. mBio 5(3): e00880-14.
Crossref
|
|
|
|
|
Alp E, Coruh A, Gunay GK, Yontar Y, Doganay M (2012). Risk factors for nosocomial infection and mortality in burn patients: 10 years of experience at a university hospital. J. Burn Care Res. 33(3):379-385.
Crossref
|
|
|
|
|
Bikard D, Marraffini LA (2012). Innate and adaptive immunity in bacteria: mechanisms of programmed genetic variation to fight bacteriophages. Curr. Opin. Immunol. 24(1):15-20.
Crossref
|
|
|
|
|
Chan BK, Abedon ST, Loc-Carrillo C (2013). Phage cocktails and the future of phage therapy. Future Microbiol. 8(6): 769-83.
Crossref
|
|
|
|
|
Chang KC, Lin NT, Hu A, Lin YS, Chen LK, Lai MJ (2012). Genomic analysis of bacteriophage ØAB1, a ØKMV-like virus infecting multidrug-resistant Acinetobacter baumannii. Genomics 97: 249–255.
Crossref
|
|
|
|
|
Fridholm H, Everitt E (2005). Rapid and reproducible infectivity end-point titration of virulent phage in a microplate system. J. Virol. Methods 128(1-2): 67-71.
Crossref
|
|
|
|
|
Hyman P, Abedon ST (2010). Bacteriophage host range and bacterial resistance. Adv. Appl. Microbiol. 70:217-48.
Crossref
|
|
|
|
|
Jin J, Li ZJ, Wang SW, Wang SM, Huang DH, Li YH, Ma YY, Wang J, Liu F, Chen XD, Li GX, Wang XT, Wang ZQ, Zhao GQ (2012). Isolation and characterization of ZZ1, a novel lytic phage that infects Acinetobacter baumannii clinical isolates. BMC Microbiol. 12:156.
Crossref
|
|
|
|
|
Kitti T, Thummeepak R, Thanwisai A, Boonyodying K, Kunthalert D, Ritvirool P, Sitthisak S (2014). Characterization and detection of endolysin gene from three Acinetobacter baumannii Bacteriophages Isolated from Sewage Water. Indian J. Microbiol. 54(4):383-8.
Crossref
|
|
|
|
|
Knezevic P, Petrovic O (2008). A colorimetric microtiter plate method for assessment of phage effect on Pseudomonas aeruginosa biofilm. J. Microbiol. Methods 74(2-3):114-118.
Crossref
|
|
|
|
|
Lee CN, Tseng TT, Lin JW, Fu YC, Weng SF, Tseng YH (2011). Lytic myophage Abp53 encodes several proteins similar to those encoded by host Acinetobacter baumannii and phage phiKO2. Appl. Environ. Microbiol. 77:6755-6762.
Crossref
|
|
|
|
|
Levin BR, Bull JJ (2004). Population and evolutionary dynamics of phage therapy. Nat. Rev. Microbiol. 2:166-173.
Crossref
|
|
|
|
|
Lin NT, Chiou PY, Chang KC, Chen LK, Lai MJ (2010). Isolation and characterization of phi AB2: a novel bacteriophage of Acinetobacter baumannii. Res. Microbiol. 161:308-314.
Crossref
|
|
|
|
|
Merabishvili M, Vandenheuvel D, Kropinski AM, Mast J, De Vos D, Verbeken G, Noben JP, Lavigne R, Vaneechoutte M, Pirnay JP (2014). Characterization of newly isolated lytic bacteriophages active against Acinetobacter baumannii. PLoS One 9(8):e104853.
Crossref
|
|
|
|
|
Niumsup PR, Boonkerd N, Tansawai U, Tiloklurs M (2009). Carbapenem-resistant Acinetobacter baumannii producing OXA23 in Thailand. Jpn. J. Infect. Dis. 62:152-154.
Pubmed
|
|
|
|
|
Park SY, Choo JW, Kwon SH, Yu SN, Lee EJ, Kim TH, Choo EJ, Jeon MH (2013). Risk factors for mortality in patients with Acinetobacter baumannii bacteremia. Infect. Chemother. 45(3): 325-30.
Crossref
|
|
|
|
|
Popova AV, Zhilenkov EL, Myakinina VP, Krasilnikova VM, Volozhantsev NV (2012). Isolation and characterization of wide host range lytic bacteriophage AP22 infecting Acinetobacter baumannii. FEMS Microbiol. Lett. 332:40-46.
Crossref
|
|
|
|
|
Rakhuba DV, Kolomiets EI, Dey ES, Novik GI (2010). Bacteriophage receptors, mechanisms of phage adsorption and penetration into host cell. Pol. J. Microbiol. 59(3):145-155.
Pubmed
|
|
|
|
|
Shen GH, Wang JL, Wen FS, Chang KM, Kuo CF, Lin CH, Luo HR, Hung CH (2012). Isolation and characterization of φkm 18p, a novel lytic phage with therapeutic potential against extensively drug resistant Acinetobacter baumannii. PLoS One 7: 1–1.
Crossref
|
|
|
|
|
Thawal ND, Yele AB, Sahu PK, Chopade BA (2012). Effect of a novel podophage AB7-IBB2 on Acinetobacter baumannii biofilm. Curr. Microbiol. 65:66-72.
Crossref
|
|
|
|
|
Yang H, Liang L, Lin S, Jia S (2010). Isolation and characterization of a virulent bacteriophage AB1 of Acinetobacter baumannii. BMC Microbiol. 10:131.
Crossref
|
|
|
|
|
Yele AB, Thawal ND, Sahu PK, Chopade BA (2012). Novel lytic bacteriophage AB7-IBB1 of Acinetobacter baumannii: isolation, characterization and its effect on biofilm. Arch. Virol. 157(8):1441-1450.
Crossref
|
|