Full Length Research Paper

ABSTRACT
Rhizobacteria are possible alternative to chemical fertilizers and used to manage phyto-pathogens. In this study, rhizobacteria isolated from rice rhizosphere of coastal agriculture system of South Andaman, India were evaluated in vitro for their plant growth promoting property (PGP), antagonistic activity and genetic diversity using PCR-RFLP. From 63 isolates, 35 were positive for salt tolerance of upto 0.5 M NaCl and 27 isolates upto 1.0 M NaCl. The biochemical characterization showed that most of the isolates were positive for one or other plant growth promoting property. Antagonistic activity against the fungi Sclerotium rolfsii revealed that 13 isolates have significantly inhibited mycelial growth of S. rolfsii as compared to the control but the isolate HMI 4, HMI 5, HMI 10, HMI 14, HMI 16 and HMI 17 were most efficient in percentage inhibition of test pathogen, caused more than 35% inhibition. RFLP of 16s rDNA showed homogenous population distributed randomly at 59% similarity. 16s rDNA sequencing result showed that most predominant population were Pseudomonas spp. followed by Enterobacter and Bacillus spp. Five best isolates were evaluated in vitro for PGP activity in rice and were found promising. The present study has resulted in the isolation of PGP and antagonistic Pseudomonas (HMI 16, HMI 17) and Bacillus spp. (HMI 14) that could be useful to design strategies to use these isolates as inoculants in sustainable agriculture of Andaman, India.
Key words: Andaman, Bacillus, phosphate solubilization, PCR-RFLP, siderophore.
INTRODUCTION
One of the most widespread agricultural problem in coastal areas is soil salinity, which limits plant growth and crop yield by reducing uptake of essential nutrients especially phosphorus. Intensive use of chemical fertilizers has resulted in increased soil salinity leading to deterioration of soil health. All the possible measures in use today, use of plant growth-promoting (PGP) bacteria is an economical and environmental friendly approach (Dixon et al., 1993). Rhizobacteria inhabiting the sites exposed to frequent stress conditions are likely to be more adaptive or tolerant and may serve as better plant growth promoter under stress conditions (Lifshitz et al., 1986).
The role of microorganisms in plant growth promotion, nutrient management and disease control is eminent.
These beneficial microorganisms colonize the rhizo-sphere of plants and promote growth of the plants through various direct and indirect mechanisms (Saxena et al., 2005). Soil microorganisms transform the insoluble forms of phosphorus (P) into soluble forms and thus influence the subsequent availability of P to plant roots. They also release siderophore for the uptake of Iron. The ability of bacteria to solubilize mineral phosphates and produce siderophore has been of interest to agricultural microbiologists because it can enhance the availability of phosphorus (P) and iron (Fe) for microbial and plant growth. Plant growth-promoting rhizobacteria (PGPR) has been reported not only to improve plant growth but also suppress the growth of plant pathogens by producing enzymes and secondary metabolites such as hydrogen cyanide (HCN), organic acids, etc (Dobbelaere et al., 2003). Pseudomonas and Bacillus spp. are immensely important in this regards as these are aggressive colonizers of the rhizosphere of various crops and have broad spectrum of antagonistic activity against many pathogens (Weller et al., 2002, Siddiqui; 2005). The mechanisms involved in PGPR-mediated plant growth promotion include synthesis of phyto-hormones such as indole-3-acetic acid (IAA), cytokinins and gibberellins (Idris et al., 2004) and an increased uptake of available minerals, nitrogen and phosphorus in the soil (Dobbelaere et al., 2003).
Rice (Oryza sativa L.) is the staple food crop of India and cultivated worldwide. In Andaman Islands, paddy is cultivated along the coastline where salinity gradient dominate. An increase in salinity led to decrease in crop productivity and rhizobacterial diversity. However, the race for producing more rice by applying more chemical fertilizers has adverse effects on the soil health and microbial community structure (Borneman et al., 1996). An understanding of structural and functional diversity of rhizobacteria is therefore essential to exploit the full potentials of these microbes for the sustained crops growth in different agro-ecosystems. With the currently available tools, the microbial community structure can be examined at several levels. The simplest analysis is based on DNA profiles, generated by PCR followed by restriction digestion of PCR product, to identify differences in the community composition. 16S rRNA gene sequences have been efficiently used for the identification of bacteria at species or strain levels (Rangarajan et al., 2001; Kumar et al., 2014). The objective of the present study was to study the genetic diversity of salt tolerant, plant growth-promoting (PGP) rhizobacterial microbes from rice rhizosphere so that a firm conclusion can be drawn on the salt-tolerant PGPR and their community structure associated with rhizo-sphere soil of rice in Andaman Islands.
MATERIALS AND METHODS
Collection of soil samples and isolation of bacterial isolates
The present study was conducted at Central Agricultural Research Institute, Port Blair during 2009-10. Rice (Oryza sativa L.) plants were grown at agricultural field of Bloomsdale farm, Central Agricultural Research Institute (CARI), Port Blair, South Andaman, India. Soil was sandy loam and its characteristics were as follows: Organic carbon (O.C) 0.35, pH 4.9, nitrogen 336 kg/ha, phosphorus 15.1 kg/ha and potassium 43 kg/ha. Fifty days after transplanting two plants from each corner and middle of the rice plot, they were uprooted and loosely adhered soil was removed. Lower 2/3 portion of roots with tightly attached soil were used for isolation of bacteria. 63 isolates of bacteria were aseptically isolated by serial dilution (10-3 to 10-6) method on Nutrient Agar medium (NA, Hi-Media). To ensure culture purity, each colony was zig-zag streaked thrice on new NA plate. All the pure cultures in tubes were maintained on NA slant at 4°C.
Salt tolerance and fluorescence test
All rhizobacterial isolates were tested for their NaCl tolerance by agar plate method. Rhizobacterial isolates were spot inoculated on freshly prepared agar plates amended separately with increasing concentration of NaCl (0.1M to 2.0 M). Plates were incubated at 28±2°C for 5 days and highest concentration of NaCl sustaining the bacterial growth was defined as the maximum tolerance (Yildirim et al., 2008). The fluorescence of cultures were identified by inoculating the isolates over the King’s B medium. After incubation for 2 days at 28±2°C, fluorescent bacterium was identified under UV light (Rouatt and Katznelson, 1961).
Phenotypic and biochemical characterization
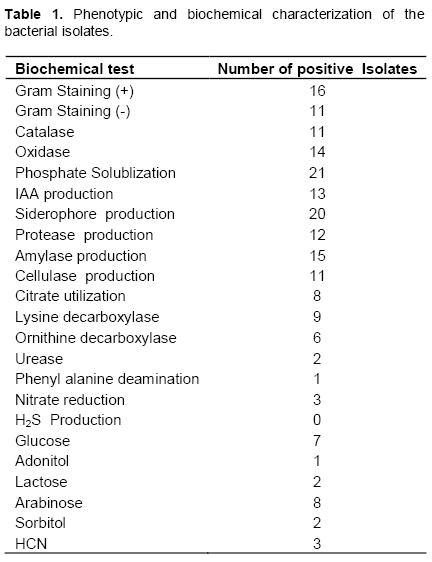
Phenotypic characterization was done on the basis of Gram’s staining, catalase, oxidase and utilization of 12 different substrates (Hi Media kit). The reactions tested were citrate utilization, lysine decarboxylase, ornithine decarboxylase, urease, phenylalanine deamination, nitrate reduction, H2S production, glucose, adonitol, lactose, arabinose and sorbitol. The results were interpreted with the help of the color change after addition of the respective reagents wherever necessary.
PO4 solublization and siderophore production
Rhizobacterial isolates were tested for phosphate solublization activity on Pikovaskaya’s Agar (PA) media (Hi-Media) and log phase bacterial culture was spot inoculated over PA. Plates were incubated at 28 ± 2°C for 2 - 3 days. Appearance of a clear halo around the colonies indicates solubilization of phosphate (Mehta and Nautiyal, 2001). Production of siderophore was determined using the chrome azurol S (CAS) agar assay. Bacterial Inoculum (10 μl) was inoculated onto the center of a CAS plate and incubated at 28 ± 2°C for 3 days. Siderophore production was assessed on the basis of change in color of the medium from blue to orange (Schwyn and Neilands, 1987).
Indole-3 acetic acid (IAA) and hydrogen cyanide (HCN) production
The bacterial isolates were grown on LB agar supplemented with L-trptophan and a sterile filter paper was placed over the culture. After five days of incubation at 28 ± 2°C, filter paper was treated with salkowski reagent (35% of perchloric acid and 0.5 M FeCl?). Development of red color indicates production of IAA by bacteria. HCN was estimated qualitatively by sulfocyanate colorimetric method. The bacterial cultures were grown on King’s B agar amended with glycin (4.4 g L) at 28 ± 2°C. One sheet of Whatmann filter paper no.1 was soaked in sodium picrate solution (2% sodium carbonate and 0.5% picric acid) for a minute and struck underneath the Petri dish lid. The plates were sealed with parafilm and incubated at 28±2°C for 48 h. Development of reddish brown color on the filter paper indicates HCN production (Vikram et al., 2007).
Production of cellulase, protease and amylase
Isolates were screened for cellulase production by plating on solid cellulose medium amended with 5 g of sodium carboxy methyl cellulose, 5 g of peptone, 20 g of agar and 10 g of yeast extract per liter. After eight days of incubation at 28 ± 2°C. Gram’s iodine solution was added and colonies surrounded by clear halos were considered positive for cellulase production (Cattelan, 1999). The protease activity was determined using skim milk agar (SMA) medium (Hi- Media). Log phase cultures were spot inoculated over SMA and incubated at 28 ± 2°C for two days. Protease activity was revealed by clear zone around the culture (Smibert and Krieg, 1994). Starch agar was used to screen the isolates producing the amylase enzyme. Cultures were spot inoculated on to the starch agar and incubated at 28 ± 2°C for two days. After incubation, iodine was added on the plates and formation of clear zone depicts positive amylase isolates.
Antagonistic test
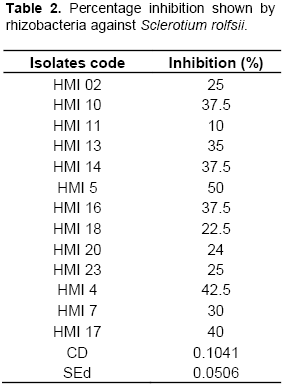
Biocontrol activity of all isolates were evaluated in vitro by dual culture test as described earlier (Skidmore and Dickinson, 1976) against single fungal plant pathogen which is Sclerotium rolfsii causing stem rot of brinjal. The pure culture of pathogen was isolated from infected part of brinjal (collar) following tissue segment method, maintained in PDA slants and preserved at 4°C. Mycelial plug (6 mm) of S. rolfsii was inoculated in the centre of petriplate previously poured with PDA (Hi Media), the bacterial isolates were streaked on the periphery of the plate. Plates were incubated at 28 ± 2°C for four days. The Petri plate with mycelial plug of pathogen only served as control. All isolates were tested in triplicates.
DNA extraction, RAPD and PCR amplification of 16s rRNA gene
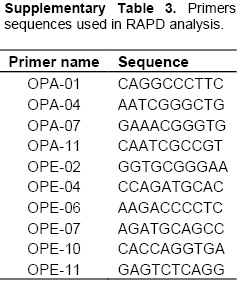
All cultures were maintained on NA at 28 ± 2°C. For DNA extraction, pure culture of 23 best isolates were grown in nutrient broth at 28 ± 2°C, 200 rpm for 24 h. Bacterial pellets were collected by centrifugation at 10,000 rpm, 4°C, 5 min. Total genomic DNA was extracted by using the method described by Sessitsch et al. (2003) with slight modification. DNA was dissolved in 100 µl 1x TE buffer. RAPD was done with 10 different primers of OPA, OPE series obtained from OPERON TECHNOLOGIES (Inc. Alameda Calif.) (Supplementary Table 3). The PCR was performed by initial denaturation at 94°C for 5 min followed by 45 cycle of denaturation at 94°C for one min., annealing at 37°C for one min and extension at 72°C for two min, and final elongation of at 72°C for 7 min. For RFLP primer PA (5’AGAGTTTGATCCTGGCTCAG 3’) and pH (5’ AAGGAGGTGATCCAGCCGCA 3’) designed by Edwards et al. (1989) were used to amplify a 1.5 Kb fragment of 16s rDNA. PCR amplification was performed in a total volume of 50 µl by mixing 50 ng of genomic DNA, 0.2 µm concentration of each primer (Gene@), 2.5 mM dNTPs (Gene@) and 1 unit of Taq DNA polymerase (Gene@) in 1x PCR buffer (Gene@). The reactions were subjected to initial denaturation of 92°C for 2.10 min followed by 45 cycle of 92°C for 1 min, 48°C for 30 s, 72°C for 2 min with a final extension of 72°C for 7 min. PCR amplified products were resolved on 1% agarose gel, prepared in 1x TAE buffer containing 0.5 µg ethidium bromide and photographed over a UV transilluminator.
RFLP and data analysis
1.5 Kb PCR product from all the 20 isolates were subjected to restriction digestion by 3 different restriction enzymes (RE) (Msp I, Hae-III, Hinf-1) in a final volume of 20 µl containing PCR product 17 µl, RE 1U (Gene@), 10x Buffer 2 µl (Gene@), incubated at 37°C for two hour. After two hour digested product was mixed with 2 µl of bromophenol blue dye and resolved over 1% agarose gel (Gene@), prepared in 1xTAE buffer containing 5 µl ethidium bromide. All the genotypes were scored for presence and absence of the RFLP bands and the data were entered into a binary matrix as discrete variables. The 0/1 matrix was used to calculate similarity as Jaccard coefficient using SIMQUAL subroutine in similarity routine. The resultant similarity matrix was employed to construct dendrogram using SAHN based UPGMA to infer genetic relationship (Rohlf, 1998).
In-vivo PGPR activity
In order to test the effective contribution to plant growth, five isolates were used for in vivo experiment. Pure bacterial cultures were grown in nutrient broth at 28±2°C until they reached a final concentration of 10 colony forming unit (CFU). Seeds of rice (Oryza sativa L) variety C-14-8 were surface sterilized in 70% ethanol for 1 min and 1.2% sodium hypochlorite for 10 min and rinsed thrice with sterile tap water. Pots filled with a sterile soil were seeded with 10 seeds/pot. Treatment consists of inoculation of 5 ml of different bacterial cultures (HMI 4, HMI 5, HMI 7, HMI 16 and HMI 17) and fertilizer. A control without microorganism and fertilizer was also maintained. Besides, each pot was irrigated with distilled water without microorganism. 30 days after sprouting, plants were harvested and data (mean) of the root and shoot length and dry matter was recorded.
Nucleotide sequence accession numbers of 16s rRNA gene
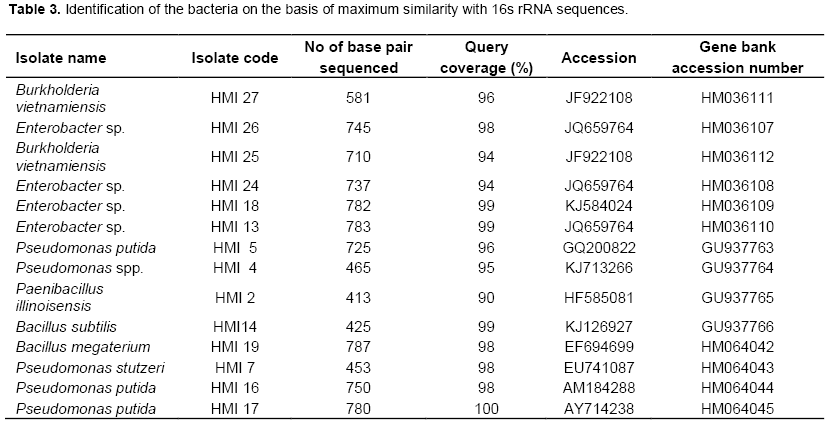
1.5 kb PCR product of some isolates was sent to commercial gene sequencing laboratory of Bangalore Genei, Bangalore, India for sequencing. Sequence analysis of these isolates were performed using BLAST (blastn)2.0 search tool (http://www.ncbi.nlm.nih.gov) available on the NCBI homepage. Unrooted phylogram was constructed using Clustal W. The nucleotide sequences of 16s rDNA were submitted in Gene bank. Their accession numbers are presented in Table 3.
RESULTS
Isolation and screening of the salt tolerant PGPR
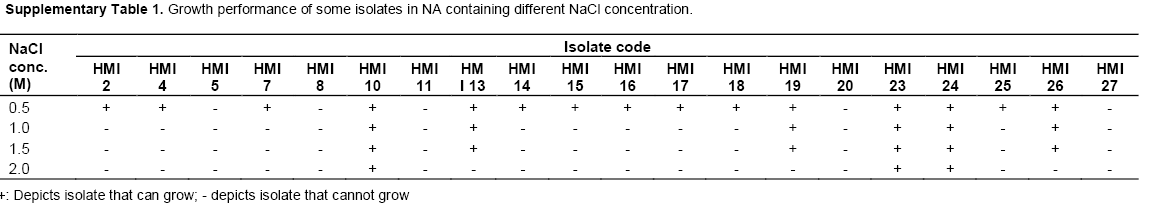
In the present study, we isolated 63 bacterial isolates from the rhizosphere soil of rice. All the bacterial isolates were evaluated for their salt tolerence trait and 35 were found positive at upto 0. 5 M NaCl, 27 were positive at upto 1.0 M NaCl, however only 3 isolates could grow in 2.0 M of NaCl. 27 isolates which survived on 1.0 M NaCl containing NA plate were further used in the study (Supplementary Table 2).
Phenotypic and biochemical characterization of rhizobacteria
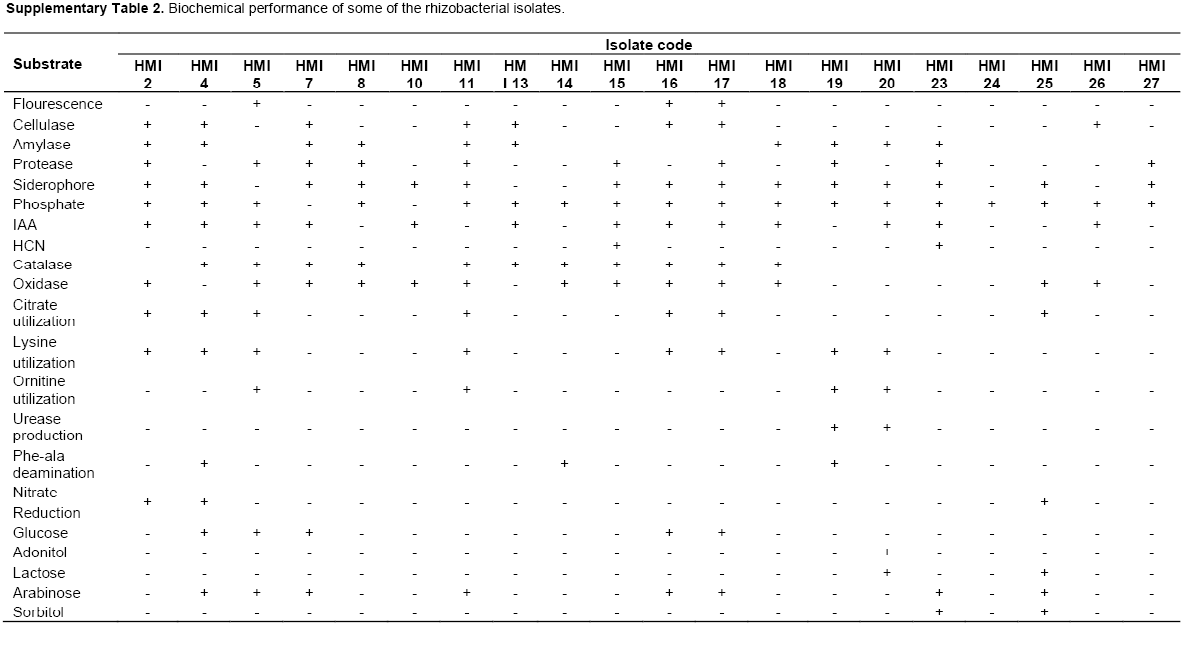
Based on the phenotypic characterization, 11 isolates (40%) were Gram negative and 16 (59%) were Gram positive. Most of the isolates (21) showed motility over motility agar. From 27 isolates 11 (40%) were positive for enzyme catalase and 14 (51%) for oxidase respectively. When biochemically characterized the isolates exhibited variation in utilization of different carbon source such as glucose, adonitol, lactose, arabinose as well as sorbitol, lysine and phenylalanine. Isolates also showed variability in production of the enzyme urease, nitrate reduction and H2S production (Table 1 and Supplementary Table 1).
Test for PGPR activity and antagonism
From 27 isolates, 20, 5, 13 and 3 isolates were positive for siderophore, fluorescence, IAA and HCN test respectively, whereas 21(77%) isolates showed phosphorus solubilization on Pikovaskaya’s agar. A significant reduction in pH of Pikovaskaya’s agar from pH 7.4 to 4.5 was observed on 10th day of incubation. The isolates were then screened for their production of the cell wall degrading and other enzymes. From 27 isolates, 12 (44%) showed proteolytic activity by inducing clear zones on the skim milk agar medium, 15 (55%) isolates were positive for amylase enzyme and 11 (40%) were positive for cellulase enzyme (Table 1 and Supplementary Table 2). The isolates were further screened for their antagonistic potential against S. rolfisii. Antagonistic activity against S. rolfsii revealed that 13 isolates have significantly inhibited mycelial growth of phyto-pathogen as compared to the control but the isolate HMI 5, HMI 4, HMI 10, HMI 14, HMI 16 and HMI 17 were most efficient in percent inhibition of test pathogen, causing more than 35% inhibition (Table 2).
Genotypic characterization
RAPD fingerprinting
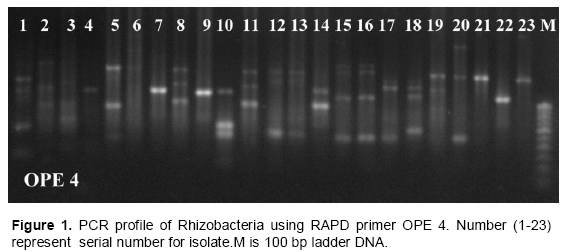
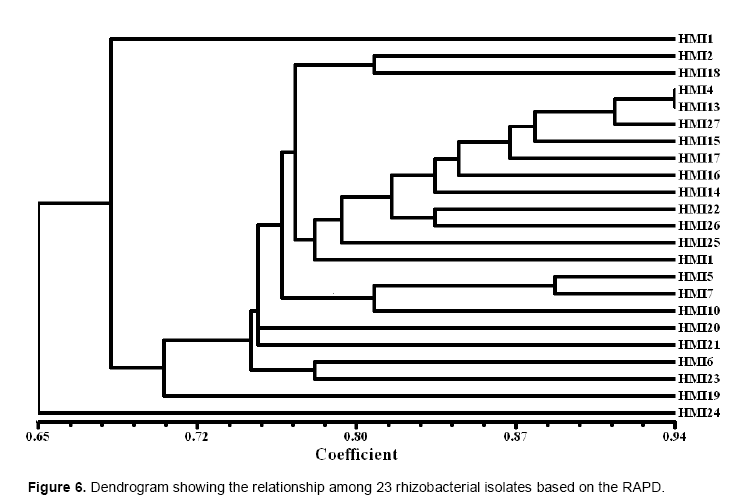
The RAPD profiles of the 23 isolates along with the markers are shown in Figure 1. All the isolates showed variation in fingerprinting pattern due to their genetic variability and distributed into different clusters. The cluster analysis based on RAPD resulted into 6 distinct clusters at a maximum similarity of 65% (Figure 6). The isolates HMI 3 and HMI 13 shows the maximum similarity of 94%.
.png)
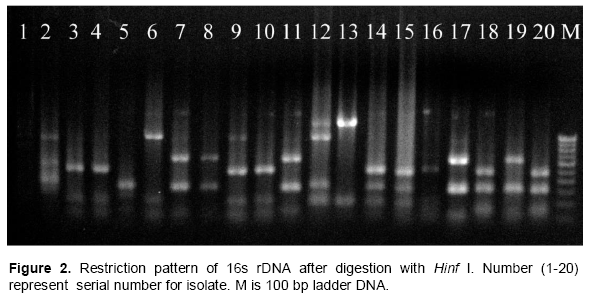
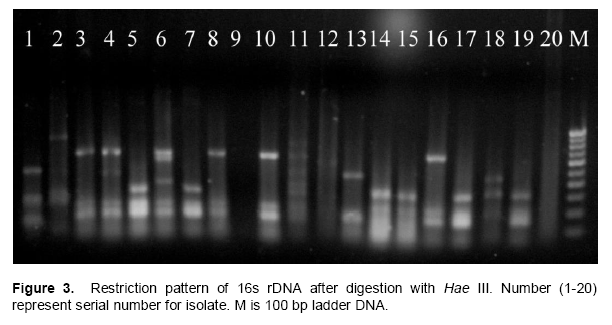
PCR-RFLP and 16S rDNA sequence
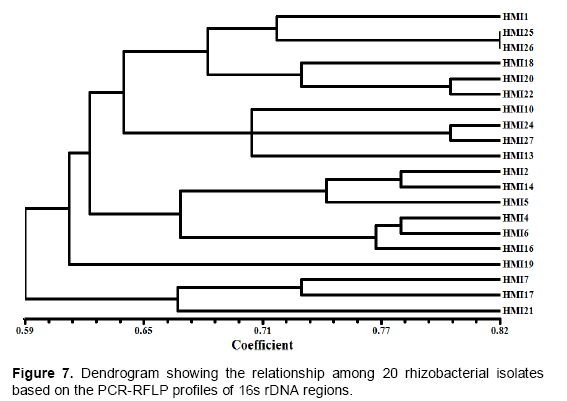
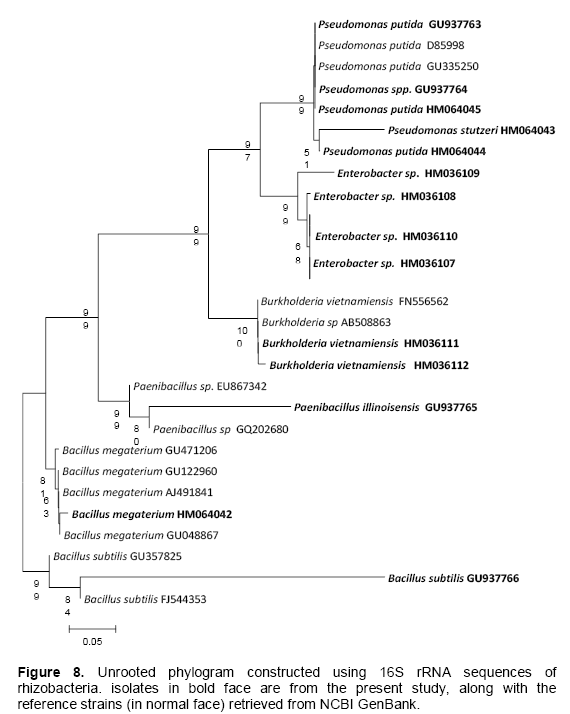
From 27 isolates 20 were used to assess the genetic diversity by PCR-RFLP. Primer PA and PH amplifies a single band of 1.5 Kb in all the isolates and no inter or intraspecific variation among the isolates were observed. A set of three different restriction enzymes (Hinf I, Hae III and Msp I) were used for RFLP analysis. All the restriction enzymes have shown restriction in all the isolates. The banding pattern generated with enzyme Hinf I showed greater polymorphism as compared to other two enzymes (Figures 2 to 3). Banding pattern generated by enzyme Hae III and Msp I was monomorphic. Based on the combined RFLP data of all the three enzymes, a dendrogram was constructed that showed at 50% similarity, all the isolates of rhizobacterial are randomly distributed (Figure 7). The dendrogram could be divided into five different clusters with similarity ranging from 0.59 to 0.82. Cluster one consist of HMI1, HMI 18, HMI 20, HMI 22, HMI 25 and HMI 26, cluster two consist of HMI 10, HMI 13, HMI 24 and HMI 27, cluster three consist of HMI 19, cluster four consist of HMI 4, HMI 6, HMI 12, HMI 14, HMI 15 and HMI 16, cluster five consist of HMI 17, HMI 17 and HMI 21 respectively. HMI 25 and HMI 26 showed 100% similarity. The 16S rDNA sequencing data showed that most of these rhizobacteria belongs to genus Pseudomonas spp., Enterobacter spp., Bacillus megateriu and Bacillus subtilis. However, some of the isolates were from genus Burkholderia vietnamiensis (Table 3). An unrooted cladogram showing similarity is present in Figure 8.
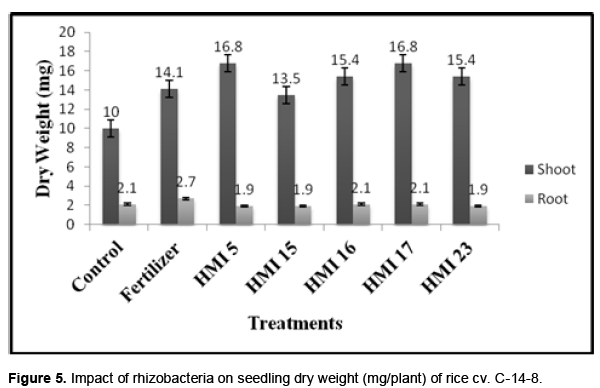
In-vitro PGPR assay
In order to confirm the PGP traits, five isolates (HMI- 4, 5, 7, 16 and 17) which showed significant PGPR activities in vitro were further evaluated for their in vivo PGP assay in rice (O. sativa L.) varieties C-14-8. Significant difference (SEd 0.05) was observed for shoot length and biomass of plant among the culture treated, fertilizer treated and control. All isolates increased the shoot length but for root no significant difference was observed among isolates (Figures 4 and 5).
DISCUSSION
Soil salinity plays an important role in the selection of microflora as environmental stress reduces their activity,
number and diversity. Bacterial adaptation to salinity mainly depends on their genetic diversity (Sudhir et al., 2009). The present investigation revealed the salt tolerant rhizobacteria diversity with innate potential of plant growth promotion and biocontrol to Sclerotium rolfsii. Population of bacterial community recovered from rice rhizosphere depicts variation in their phenotypic charac-teristics. These differences indicate significant diversity within the cultureable population. In our study, 21 and 20 isolates were positive for phosphate solubilization and siderophore production, respectively. The ability of bacteria to solubilize mineral phosphate and produce siderophore is a field of immense interest to agricultural microbiologists because it can enhance the availability of P and Fe for microbial and plant growth. It is believed that microbial mediated solubilization of insoluble phosphorus in soil is through the release of organic acids (Nautiyal et al., 2000). However, in addition to biotic acidification, other mechanisms such as chelation and exchange reactions can cause phosphate solubilization (Illumer and Schinner, 1995; Richardson et al., 2001; Gyaneshwar et al., 1998; Rodriguez et al., 2004). Siderophore production is another PGP feature that may influence plant growth by binding to the available iron form (Fe3+) in the rhizosphere. Through this process, iron is made unavailable to the phytopathogens and at the same time siderophore protects the plant health (Siddiqui, 2005).
IAA producing bacteria are known to promote plant growth (Patten and Glick, 2002). In our study, 13 isolates produced a substantial amount of IAA, however variability in the amount of IAA was observed, suggesting that these isolates could be used for plant growth promotion. Mirza et al. (2001) showed that indolic compounds production can vary among different species and strains and is also influenced by culture conditions, growth stage and availability of the substrate.
Many Bacillus and Pseudomonas spp. are capable of producing a range of different enzymes, antibiotics and toxins, many of which can be antagonistic to plant pathogens (El-Banna, 2005; Chakraborty et al., 2006). In the present study, Pseudomonas and Bacillus isolates have revealed better antagonistic activities than other isolates. Most of the Pseudomonas isolates inhibited growth of plant pathogens by 37.5-50% while isolates of Bacillus showed 37.5%. Reduction in the growth of phyto-pathogen could be due to the secretion of lytic enzymes, HCN, siderophore and other secondary metabolites. Voisard et al. (1989) reported that cyanide released by Pseudomonas flourescens suppresses the growth of phytopathogens sharing the same ecological niche, thereby acting as a biocontrol metabolite. Various species of Pseudomonas have been reported to promote plant growth and suppress disease in plants. A mixture of Bacillus amyloliquefaciens and Bacillus pumilus were found to provide a broad spectrum of protection against both soil and air-borne pathogens including cucumber mosaic virus, Colletotrichum gloeosporioides, Ralstonia solanacearum, Rhizoctonia solani and Sclerotium rolfsii (Jetiyanon et al., 2003). Adhikari et al. (2001) reported that bacterial strains isolated from the rice rhizosphere have the potential to control the seedling disease of rice. On the basis of PGPR and antagonistic activities, five Pseudomonas isolates were selected for the study of the effect of isolates on the growth performance of rice. An increase in plant growth was observed for all the tested isolated. This was due to the secretion of IAA, solubilization of phosphorus and secretion of other various beneficial secondary metabolites. All the tested isolates have shown enhanced shoot growth and biomass as compared to the control. However, no significant variation was observed for root length and biomass as higher concentration of IAA (auxin) is stimulatory for the shoot growth but inhibitory for root.
16S rRNA gene sequencing is a widely used standard technique in bacterial taxonomy and it is also routinely used in ‘polyphasic approach’ when new descriptions of bacterial species or higher taxa are made (Ludwig and Schleifer, 1999; Rossello-Mora, 2005). Genotypic analysis by PCR-RFLP as well as 16s rRNA gene sequencing analysis revealed a low degree of diversity among the isolates. In PCR-RFLP, there was a difference in only one of the restriction pattern studied. This was with Hinf I which shows a common restriction pattern in all except 9 isolates. Similarity in the digestion pattern of the other enzymes used and 11 isolates that showed any variation suggests a predominant homogenous bacterial population. Homogeneity in bacterial population can be explained on the basis of bacterial ability to tolerate salt stress. The variation in the tolerance could probably be due to the fact that rhizobacteria adopt different strategies to overcome the toxic effect of the salinity. The dominance of some components of the bacterial population in the rhizosphere could be due to rhizodeposition as size and composition of the rhizospheric microflora depends on plant species (Lynch and Whipps, 1990). Rangarajan et al. (2001) analyzed population of Pseudomonas for their biochemical characters and genetic diversity using RAPD and RFLP and found that increased salinity caused selection of P. pseudoalcaligenes and P. alcaligenes, irrespective of the host rhizosphere.
The present work identified isolates of plant growth promoting rhizobacteria with potential for biological control. These microorganisms are of particular interest because they have revealed in vitro PGP and bio-control activity against tested plant pathogen. Despite in vitro suppression of test pathogen, an in vivo PGP activity was observed. These isolates (HMI 14, HMI 16 and HMI 17) may prove to be novel PGPR isolates for effective formulation of bio-fertilizer to saline affected areas. These isolates could be utilized to improve the fertility status of soil, soil health, to protect the environment and will be cost effective. Since benefits from the PGPR could be synergistic, further experiments are needed to determine the effectiveness of these isolates under different field conditions and to understand the nature of interaction with the host plant.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENT
The authors are grateful to Indian Council of Agricultural Research (ICAR) New Delhi for the financial support.
REFERENCES
|
Adhikari TB, Joseph CM, Yang G, Phillips DA, Nelson LM (2001). Evaluation of bacteria isolated from rice for plant growth promotion and biological control of seedling disease of rice. Can. J. Micro. 47(10):916-924. |
|
|
Borneman J, Skroch PW, O'Sullivan KM, Palus JA, Rumjanek NG, Jansen JL, Nienhuis J, Triplett EW (1996). Molecular microbial diversity of an agricultural soil in Wisconsin. Appl. Environ. Microbiol. 62:1935-1943. |
|
|
Cattelan AJ, Hartel PG, Furhmann FF (1999). Screening for plant growth promoting rhizobacteria to promote early soybean growth. Soil Sci. Soc. Am. J. 63:1670-1680. |
|
|
Chakraborty U, Chakraborty B, Basnet M (2006). Plant growth promotion and induction of resistance in Camellia sinensis by Bacillus megaterium. J. Basic Microbiol. 46:186-195. |
|
|
Dixon RK, Gang VK, Rao MV (1993). Inoculation of Leucaena and Prosopis seedlings with Glomus and Rhizobium species in saline soil: rhizosphere relations and seedlings growth. Arid Soil Res. Rehabil. 7:133-144. |
|
|
Dobbelaere S, Vanderleyden J, Okon Y (2003). Plant growth-promoting effects of diazotrophs in the rhizosphere. Crit. Rev. Plant Sci. 22:107-149. |
|
|
Edwards U, Rogall T, Blocker H, Emde M, Bottger EC (1989). Isolation and direct complete nucleotide determination of entire genes. Characterization of a gene coding for 16S ribosomal RNA. Nucleic Acid. Res. 17:7843-7853. |
|
|
El-Banna NM (2005). Effect of carbon source on the antimicrobial activity of the air flora. World J. Microbiol. Biotechnol. 21:1451-1454. |
|
|
Gyaneshwar P, Kumar GN, Parekh LJ, (1998). Effect of buffering on the phosphate-solubilizing ability of microorganisms. World J. Microbiol. Biotechnol. 14: 669-673. |
|
|
Idris EES, Bochow H, Ross H, Borriss R (2004). Use of Bacillus subtilis as biocontrol agent. Phytohormone-like action of culture filtrates prepared from plant growth-promoting Bacillus amyloliquefaciens FZB24, FZB42, FZB45 and Bacillus subtilis FZB37. J. Plant Dis. Prot. 111: 583-597. |
|
|
Illumer P, Schinner F (1995). Solubilization of inorganic calcium phosphate- solubilization mechanisms. Soil Biol. Biochem. 27:257-263. |
|
|
Jetiyanon K, Jetiyanon WD, Kloepper JW (2003). Broad-spectrum protection against several pathogens by PGPR mixtures under field conditions in Thailand. Plant Dis. 87:1390-1394. |
|
|
Kumar A, Kumar A, Pratush A (2014). Molecular diversity and functional variability of environmental isolates of Bacillus species. SpringerPlus 3:312. |
|
|
Lifshitz R, Kloepper JW, Scher FM, Tipping EM, Laliberte M (1986). Nitrogen-fixing Pseudomonads isolated from roots of plants grown in the Canadian high arctic. Appl. Environ. Microbiol. 51:251-255. |
|
|
Ludwig W, Schleifer KH (1999). Phylogeny of bacteria beyond the 16S rRNA standard. ASM News 65:752-757 |
|
|
Lynch JM, Whipps JM (1990). Substrate flow in the rhizosphere. Plant Soil 129: 1-10. |
|
|
Mehta S, Nautiyal CS (2001). An efï¬cient method for qualitative screening of phosphate solubilizing bacteria. Curr. Microbiol. 43:51-56. |
|
|
Mirza MS, Ahmad W, Latif F, Haurat J, Bally R., Normand P, Malik KA (2001). Isolation, partial characterization, and the effect of plant growth-promoting bacteria (PGPB) on micro-propagated sugarcane in vitro. Plant Soil 237:47-54, |
|
|
Nautiyal CS, Bhadauria S, Kumar P, Lal H, Mondal R, Verma D (2000). Stress induced phosphate solubilization in bacteria isolated from alkaline soils. FEMS Microbiol. Lett. 182:291-296. |
|
|
Patten C, Glick CR (2002). Role of Pseudomonas putida indole acetic acid in development of host plant root system. Appl. Environ. Microbiol. 68:3795-3801. |
|
|
Rangarajan S, Loganathan P, Saleena LM. Nair S, (2001). Diversity of Pseudomonads isolated from three different plant rhizospheres. J. Appl. Microbiol. 91:742-749. |
|
|
Richardson AE, Hadobas PA, Hayes JE, O'Hara JE, Simpson RJ (2001). Utilization of phosphorus by pasture plants supplied with myoinositol hexaphosphate is enhanced by the presence of soil microorganisms. Plant Soil 229:47-56. |
|
|
Rodriguez H, Gonzalez T, Goire, I, Bashan Y (2004). Gluconic acid production and phosphate solubilization by the plant growth promoting bacterium spp. Naturwissenschaften 91:552-555. |
|
|
Rohlf FJ (1998). NTSYS: numerical taxonomy and multivariate analysis system, 2nd ed. Exeter Software, State University of New York, Stony Brook, N.Y |
|
|
Rossello-Mora R (2005). Updating prokaryotic taxonomy. J. Bacteriol. 187:6255-6257. |
|
|
Rouatt JW, Katznelson H (1961). A study of bacteria on the root surface and in the rhizosphere soil of crop plants. J. Appl. Bacteriol. 24:164-171. |
|
|
Saxena AK, Lata SR, Pandey AK (2005). Culturing of plant growth promoting rhizobacteria. In: Gopi KP, Varma A (eds) Basic research applications of mycorrhizae. I K International Pvt Ltd, New Delhi. pp. 453-474. |
|
|
Schwyn B, Neilands JB (1987). Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 160:47-56. |
|
|
Sessitsch A, Kan FY, Pfeifer U (2003). Diversity and community structure of culturable Bacillus spp. Populations in the rhizospheres of transgenic potatoes expressing the lytic peptide cecropin B. Appl. Soil Ecol. 22:149-158. |
|
|
Siddiqui ZA (2005). PGPR: prospective biocontrol agents of plant pathogens. Biocontrol and Biofertilization, Springer, Netherlands. pp. 111-142. |
|
|
Skidmore AM, Dickinson CH (1976). Colony interaction and hyphal interference between Sartoria nodorum and phylloplane fungi. Trans. Br. Mycol. Soc. 66: 57-64. |
|
|
Smibert RM, Krieg NR (1994). Phenotypic characterization. In: Gerhardt P, Murray RGE,Wood WA, Krieg NR editors. Methods for General and Molecular Bacteriology. American Society of Microbiology, Washington DC: 607-654. |
|
|
Sudhir K, Upadhyay DP, Singh RS (2009). Genetic diversity of plant growth promoting Rhizobacteria isolated from rhizospheric soil of wheat under saline conditions. Curr. Microbiol. 59:489-496. |
|
|
Vikram A, Hamzehzarghani H, Alagawadi AR, Krishnaraj PU, Chandrashekar BS (2007). Production of plant growth promoting substances by phosphate solubilizing bacteria isolated from vertisols. J. Plant Sci. 2:326-333. |
|
|
Voisard CC, Keel D, Haas G (1989). Cyanide production by Pseudomonas fluorescens helps suppress black root rot of tobacco under gnobiotic conditions. EMBO J. 8: 351-358. |
|
|
Weller DM, Raaijimakers JM, Gardners BBM, Thomashow LS (2002). Microbial population responsible for specific soil suppressiveness to plant pathogens. Ann. Rev. Phytopathol. 40:309-348. |
|
|
Yildirim E, Turan M, Donmez MF (2008). Mitigation of salt stress in radish (Raphanus sativus l.) by plant growth promoting rhizobacteria. Rom. Biotechnol. Lett. 13:3933-3943. |
|
APPENDIX
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


