Full Length Research Paper

ABSTRACT
Milk is a substantial source of nutrients needed by all humans across lifespan development. Given its nutritional composition, milk is considered a vehicle for various microbes including beneficial and pathogenic bacteria. In this study, 270 milk samples comprising raw cow and buffalo milk and pasteurized milk with different shelf-life durations were tested along with pasteurized organic milk for the presence of Staphylococcus aureus and Escherichia coli. Collectively, 21 E. coli and 14 S. aureus isolates were cultivated and identified from total milk samples. All E. coli and S. aureus isolates exhibited resistance to erythromycin and penicillin, respectively. Serogroups O26, O128, and O111 were the most frequently identified amongst E. coli isolates, whereas staphylococcal enterotoxins (SEs) were inconsistently produced across S. aureus isolates. The molecular profile showed clustering of 6 isolates of E. coli by harboring stx1, stx2, eaeA genes, and 5 isolates of S. aureus by mecA gene. Findings revealed the bacteriological quality of popularly consumed milk in Egypt, including raw and pasteurized milk with preference to pasteurized organic milk and 7-day shelf life (7DSL) pasteurized milk. However, raw milk and 3MSL pasteurized milk were the major sources of E. coli and S. aureus, posing a serious public health issue.
Key words: Raw milk, pasteurization, Staphylococcus aureus and Escherichia coli, shelf-life.
INTRODUCTION
MATERIALS AND METHODS
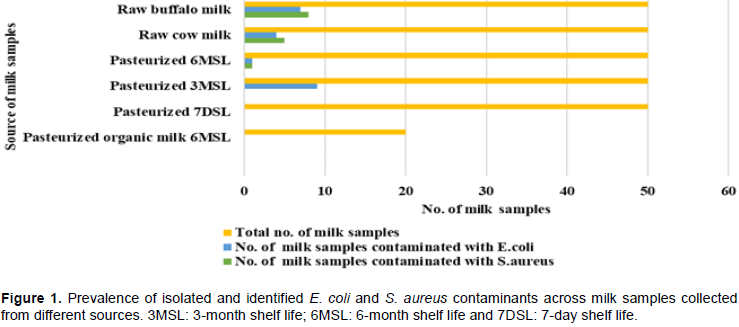
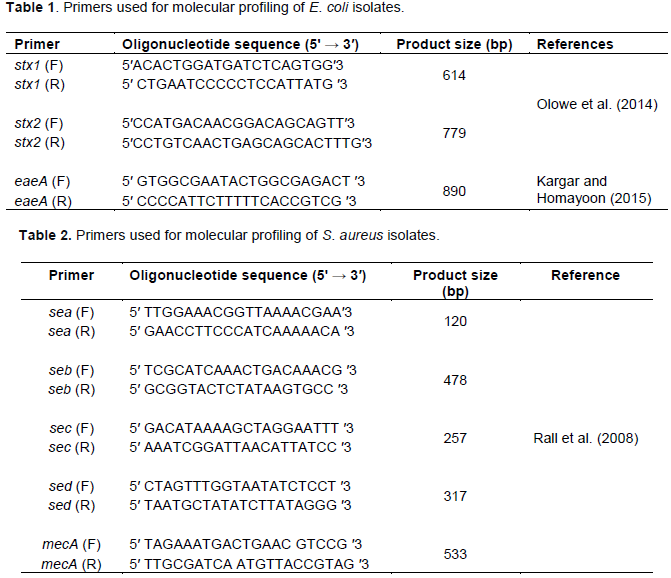
RESULTS
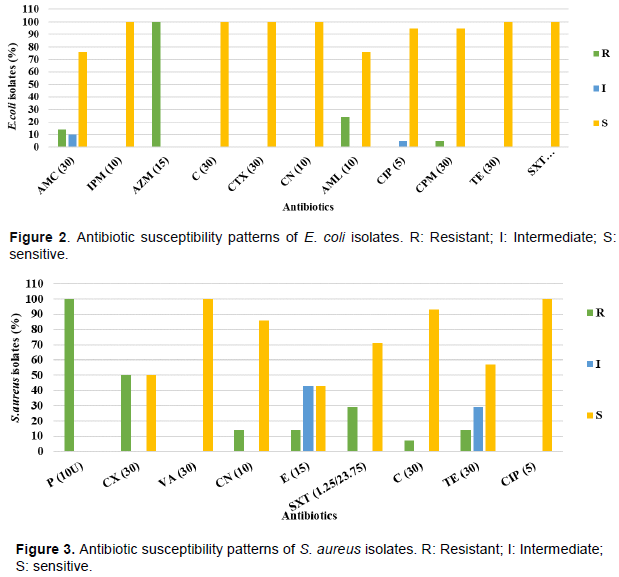
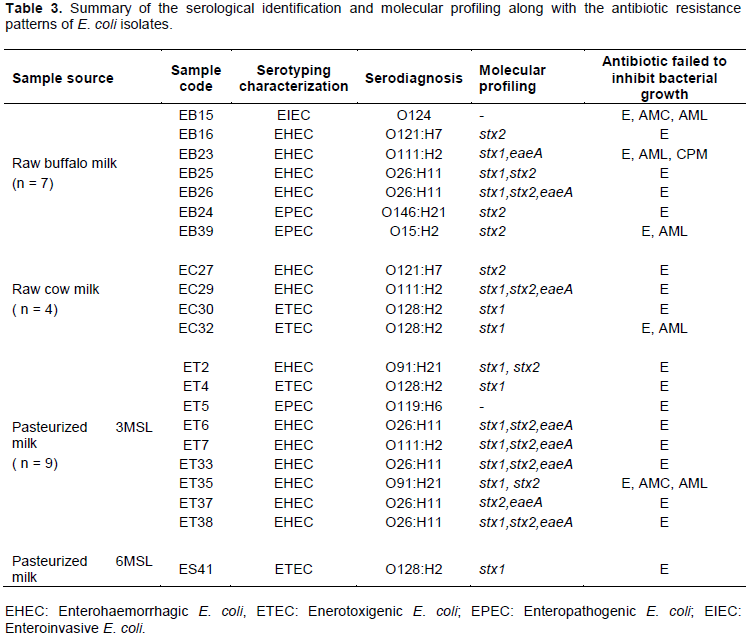
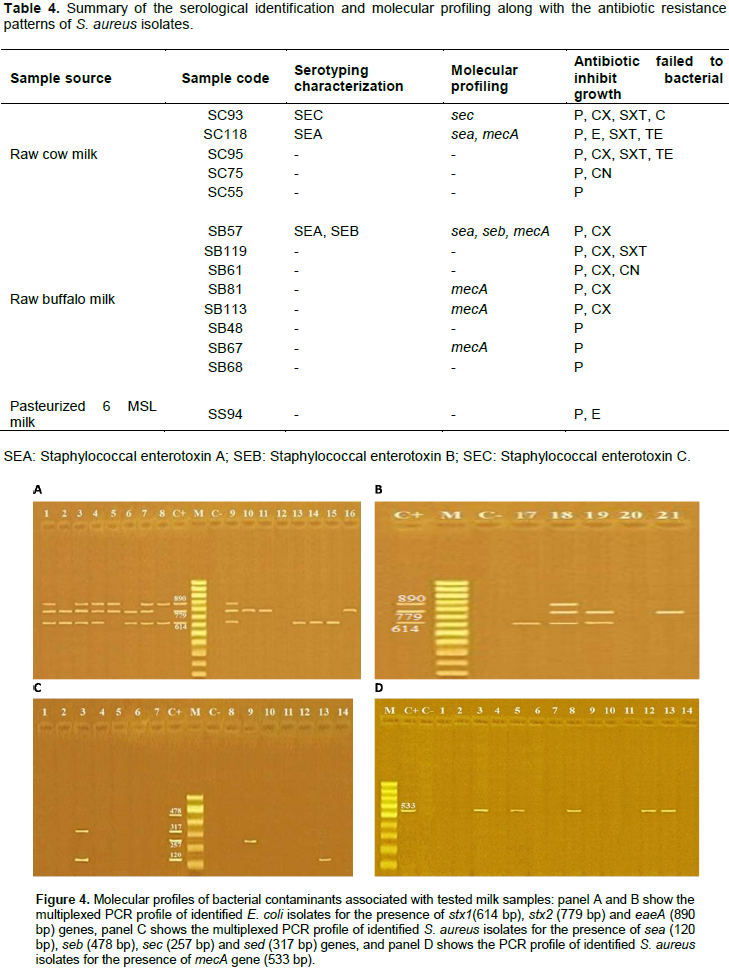
DISCUSSION
CONCLUSION
To conclude, findings revealed that raw and 3MSL pasteurized milk are most prone to be contaminated by the pathogenic E. coli and S. aureus isolates, that poses serious health issues upon direct consumption of milk from these sources. Noteworthy, pasteurized organic milk and 7DSL milk were found to be of the highest bacteriological quality when tested for the presence of E. coli and S. aureus. Eventually, our findings implicitly highlighted the importance of constituting strict regulations with regard to milk handling in local farms and dairy plants to minimize the chance of milk contamination and the transmission of bacterial pathogens along with their antimicrobial resistance from dairy animals to humans.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENT
The authors would like to thank the Department of Microbiology, Faculty of Veterinary Medicine, Damanhour University for their kind cooperation and contribution to this study. This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.
REFERENCES
|
Abebe M, Hailelule A, Abrha B, Nigus A, Birhanu M, Adane H, Genene T, Daniel H, Getachew G, Merga G, Haftay A (2014). Antibiogram of Escherichia coli strains isolated from food of bovine origin in selected Woredas of Tigray, Ethiopia. Journal of Bacteriology Research 6(3):17-22. |
|
|
Aidar-Ugrinovich L, Blanco J, Blanco M, Blanco JE, Leomil L, Dahbi G, Mora A, Onuma DL, Silveira WD, Pestana de Castro AF (2007). Serotypes, virulence genes, and intimin types of Shiga toxin-producing Escherichia coli (STEC) and enteropathogenic E. coli (EPEC) isolated from calves in São Paulo, Brazil. International Journal of Food Microbiology 115(3):297-306. |
|
|
Arcuri EF, Angelo FF, Guimarães MFM, Talon R, Borges MF, Leroy S, Loiseau G, Lange CC, Andrade NJ, Montet D (2010). Toxigenic status of Staphylococcus aureus isolated from bovine raw milk and Minas frescal cheese in Brazil. Journal of Food Protection 73(12):2225-2231. |
|
|
Argudín MÁ, Mendoza MC, Rodicio MR (2010). Food Poisoning and Staphylococcus aureus Enterotoxins. Toxins 2(7):1751-1773. |
|
|
Asao T, Kumeda Y, Kawai T, Shibata T, Oda H, Haruki K, Nakazawa H, Kozaki S (2003). An extensive outbreak of staphylococcal food poisoning due to low-fat milk in Japan: estimation of enterotoxin A in the incriminated milk and powdered skim milk. Epidemiology and Infection 130(1):33-40. |
|
|
Awadallah MA, Ahmed HA, Merwad AM, Selim MA (2016). Occurrence, genotyping, shiga toxin genes and associated risk factors of E. coli isolated from dairy farms, handlers and milk consumers. The Veterinary Journal 217:83-88. |
|
|
Bali OS, Lajnef R, Felfoul I, Attia H, Ayadi MA (2013). Detection of Escherichia coli in unpasteurized raw milk. International Journal of Agriculture and Food Science 3(2):53-55. |
|
|
Barbano DM, Ma Y, Santos MV (2006). Influence of raw milk quality on fluid milk shelf life. Journal of Dairy Science 89:E15-E19. |
|
|
Bennett SD, Walsh KA, Gould LH (2013). Foodborne Disease Outbreaks Caused by Bacillus cereus, Clostridium perfringens, and Staphylococcus aureus-United States, 1998-2008. Clinical Infectious Diseases 57(3):425-433. |
|
|
Blair JM, Webber MA, Baylay AJ, Ogbolu DO, Piddock LJ (2015). Molecular mechanisms of antibiotic resistance. Nature Reviews Microbiology 13(1):42-51. |
|
|
Clinical and Laboratory Standards Institute (CLSI) (2017). Performance standards for antimicrobial disk susceptibility tests; approved standard-12th ed. M02-A13. Clinical and Laboratory Standards Institute, Wayne, PA. |
|
|
De Oliveira GB, Favarin L, Luchese RH, McIntosh D (2015). Psychrotrophic bacteria in milk: How much do we really know? Brazilian Journal of Microbiology 46(2):313-321. |
|
|
Deeth H (2017). Optimum Thermal Processing for Extended Shelf-Life (ESL) Milk. Foods (Basel, Switzerland) 6(11). |
|
|
Desmarchelier P, Fegan N (2016). Pathogens in Milk: Escherichia coli. Reference Module in Food Science. https://doi.org/10.1016/B978-0-08-100596-5.00989-6 |
|
|
Dhanashekar R, Akkinepalli S, Nellutla A (2012). Milk-borne infections. An analysis of their potential effect on the milk industry. Germs 2(3):101-109. |
|
|
Disassa N, Sibhat B, Mengistu S, Muktar Y, Belina D (2017). Prevalence and Antimicrobial Susceptibility Pattern of E. coli O157:H7 Isolated from Traditionally Marketed Raw Cow Milk in and around Asosa Town, Western Ethiopia. Veterinary Medicine International. |
|
|
Elgadi ZAM, Abdel Gadir WS, Dirar HA (2008). Isolation and Identification of Lactic Acid Bacteria and Yeast from Raw Milk in Khartoum State (Sudan). Research Journal of Microbiology 3(3)163-168. |
|
|
Fagundes H, Barchesi L, Filho AN, Ferreira LM, Oliveira CAF (2010). Occurrence of Staphylococcus aureus in raw milk produced in dairy farms in São Paulo state, Brazil. Brazilian Journal of Microbiology 41(2):376-380. |
|
|
Food and Agriculture Organization (FAO) (2011). World Livestock. Livestock in Food Security, Rome, Italy. |
|
|
Fernandes R (2009). Liquid milk products. In: Fernandes R., Microbiology handbook dairy products. Leatherhead Food International and RSC Publishing pp. 1-19. |
|
|
Garedew L, Berhanu A, Mengesha D, Tsegay G (2012). Identification of gram-negative bacteria from critical control points of raw and pasteurized cow milk consumed at Gondar town and its suburbs, Ethiopia. BMC Public Health 12:950. |
|
|
Hennekinne JA, De Buyser ML, Dragacci S (2012). Staphylococcus aureus and its food poisoning toxins: characterization and outbreak investigation. FEMS Microbiology Reviews 36(4):815-836. |
|
|
Hoque MN, Das ZC, Rahman ANMA, Haider MG, Islam MA (2018). Molecular characterization of Staphylococcus aureus strains in bovine mastitis milk in Bangladesh. International Journal of Veterinary Science and Medicine 6(1):53-60. |
|
|
Kadariya J, Smith TC, Thapaliya D (2014). Staphylococcus aureus and Staphylococcal Food-Borne Disease: An Ongoing Challenge in Public Health. BioMed Research International pp. 1-9. |
|
|
Kandil AA, Elhadidy M, El-Gamal A, Al-Ashmawy MA (2018). Identification of S. aureus and E. coli from Dairy Products Intended for Human Consumption. Advances in Animal and Veterinary Sciences 6(11):509-513. |
|
|
Kaper JB, Nataro JP, Mobley HLT (2004). Pathogenic Escherichia coli, Nature reviews, Microbiology 2(2):123-40. |
|
|
Kargar M, Homayoon M (2015). Prevalence of shiga toxins (stx1, stx2), eaeA and hly genes of Escherichia coli O157:H7 strains among children with acute gastroenteritis in southern of Iran. Asian Pacific Journal of Tropical Medicine 8(1):24-28. |
|
|
Kreig N, Holt J (1984). Bergey's Manual of systemic bacteriology (1) Williams and Wilkins, Baltimore M.D.21202, USA. |
|
|
Lachica RVF, Genigeorgis C, Hoeprich PD (1971). Metachromatic agar-diffusion methods for detecting staphylococcal nuclease activity. Applied Microbiology 21(4):585-587. |
|
|
Lewis SJ, Gilmour A (1987). Microflora associated with the internal surfaces of rubber and stainless steel milk transfer pipeline. Journal of Applied Bacteriology 62(4):327-333. |
|
|
Hassan GM, Meshref AMS, Gomaa SM (2015). Microbiological Quality and Safety of Fluid Milk Marketed in Cairo and Giza Governorates. Current Research in Dairy Sciences 7(1):18-25. |
|
|
MacFaddin JF (2000). Biochemical Tests for Identification of Medical Bacteria. 3rd Edition, Lippincott Williams & Wilkins, Philadelphia. |
|
|
Mallet A, Guéguen M, Kauffmann F, Chesneau C, Sesboué A, Desmasures N (2012). Quantitative and qualitative microbial analysis of raw milk reveals substantial diversity influenced by herd management practices. International Dairy Journal 27(1-2):13-21. |
|
|
Mansour AS, Wagih GE, Morgan SD, Elhariri M, El-shabrawy MA, Abuelnaga ASM, Elgabry EA (2017). Detection of Staphylococcus aureus enterotoxigenic strains in bovine raw milk by reversed passive latex agglutination and multiplex polymerase chain reaction. Veterinary World 10(8):843-847. |
|
|
Matallah AM, Bouayad L, Boudjellaba S, Mebkhout F, Hamdi TM, Ramdani-Bouguessa N (2019). Staphylococcus aureus isolated from selected dairies of Algeria: Prevalence and susceptibility to antibiotics. Laboratory of Medical Biology 12(2):205-210. |
|
|
McEwen SA, Collignon PJ (2018). Antimicrobial Resistance: a One Health Perspective. Microbiology Spectrum 6(2). |
|
|
Miller JH (1992). A Short Course in Bacterial Genetics: A Laboratory Manual and Handbook for Escherichia coli and related bacteria. Plainview, NY: Cold Spring Harbor Laboratory Press; 1992. |
|
|
Momtaz H, Farzan R, Rahimi E, Safarpoor Dehkordi F, Souod N (2012). Molecular characterization of Shiga toxin-producing Escherichia coli isolated from ruminant and donkey raw milk samples and traditional dairy products in Iran. The Scientific World Journal 2012:231342. |
|
|
Mora A, Blanco JE, Blanco M, Alonso MP, Dhabi G, Echeita A, González EA, Bernárdez MI, Blanco J (2005). Antimicrobial resistance of Shiga toxin (verotoxin)-producing Escherichia coli O157:H7 and non-O157 strains isolated from humans, cattle, sheep and food in Spain. Research in Microbiology 156(7):793-806. |
|
|
Morandi S, Brasca M, Lodi R, Cremonesi P, Castiglioni B (2007). Detection of classical enterotoxins and identification of enterotoxin genes in Staphylococcus aureus from milk and dairy products. Veterinary Microbiology 124(1-2):66-72. |
|
|
Murphy SC, Martin NH, Barbano DM, Wiedmann M (2016). Influence of raw milk quality on processed dairy products: How do raw milk quality test results relate to product quality and yield? Journal of Dairy Science 99(12):10128-10149. |
|
|
Nataro JP, Kaper JB (1998). Diarrheagenic Escherichia coli. Clinical Microbiology Reviews 11(1):142-201. |
|
|
Neher S, Hazarika AK, Sharma RK, Barkalita LM, Bora M, Deka P (2015). Detection of Shiga-Toxigenic Escherichia Coli in Milk Samples of Cattle by PCR. Journal of Agriculture and Veterinary Science 8(5):75-78. |
|
|
Nobili G, Franconieri I, Basanisi MG, La Bella G, Tozzoli R, Caprioli A, La Salandra G (2016). Short communication : Isolation of Shiga toxin-producing Escherichia coli in raw milk and mozzarella cheese in southern Italy. Journal of Dairy Science 99(10):7877-7880. |
|
|
Olowe OA, Aboderin BW, Idris OO, Mabayoje VO, Opaleye OO, Adekunle OC, Olowe RA, Akinduti PA, Ojurongbe O (2014). Genotypes and phenotypes of Shiga toxin-producing Escherichia coli (STEC) in Abeokuta, Southwestern Nigeria. Infection and Drug Resistance 7:253-259. |
|
|
Ombarak RA, Zayda MG, Hinenoya A, Yamasaki S (2019). Serotypes, pathogenic potential and antimicrobial resistance of Escherichia coli isolated from subclinical bovine mastitis milk samples in Egypt. Japanese Journal of Infectious Diseases 72(5):337-339. |
|
|
Park CE, Szabo R (1986). Evaluation of the reversed passive latex agglutination (RPLA) test kits for detection of staphylococcal enterotoxins A, B, C, and D in foods. Canadian Journal of Microbiology 32(9):723-727. |
|
|
Postollec F, Falentin H, Pavan S, Combrisson J, Sohier D (2011). Recent advances in quantitative PCR (qPCR) applications in food microbiology. Food Microbiology 28(5):848-861. |
|
|
Quigley L, McCarthy R, O'Sullivan O, Beresford TP, Fitzgerald GF, Ross RP, Stanton C, Cotter PD (2013a). The microbial content of raw and pasteurized cow milk as determined by molecular approaches. Journal of Dairy Science 96(8):4928-4937. |
|
|
Quigley L, O'Sullivan O, Stanton C, Beresford TP, Ross RP, Fitzgerald GF, Cotter PD (2013b). The complex microbiota of raw milk. FEMS Microbiology Reviews 37(5):664-698. |
|
|
Rais M, Acharya S, Sharma N (2013). Food processing industry in India: S&T capability, skills and employment opportunities. Journal of Rural Development 4(9):1-13. |
|
|
Rall VLM, Vieira FP, Rall R, Vieitis RL, Fernandes A, Candeias JMG, Cardoso KF, Araújo JP (2008). PCR detection of staphylococcal enterotoxin genes in Staphylococcus aureus strains isolated from raw and pasteurized milk. Veterinary Microbiology 132(3-4):408-413. |
|
|
Ranjbar R, Dehkordi FS, Shahreza MHS, Rahimi E (2018). Prevalence, identification of virulence factors, O-serogroups and antibiotic resistance properties of shiga-toxin producing Escherichia coli strains isolated from raw milk and traditional dairy products. Antimicrobial resistance and infection control 7(53). |
|
|
Reta MA, Bereda TW, Alemu AN (2016). Bacterial contaminations of raw cow's milk consumed at Jigjiga City of Somali Regional State, Eastern Ethiopia. International Journal of Food Contamination 3(1):4. |
|
|
Sarkar S (2015). Microbiological Considerations: Pasteurized Milk. International Journal of Dairy Science 10:206-218. |
|
|
Schroeder CM, Meng J, Zhao S, Debroy C, Torcolini J, Zhao C, McDermott PF, Wagner DD, Walker RD, White DG (2002). Antimicrobial resistance of Escherichia coli O26, O103, O111, O128, and O145 from animals and humans. Emerging infectious diseases 8(12):1409-1414. |
|
|
Shah MS, Qureshi S, Kashoo Z, Farooq S, Wani SA, Hussain MI, Banday MS, Khan AA, Gull B, Habib A, Khan SM, Dar BA (2019). Methicillin resistance genes and in vitro biofilm formation among Staphylococcus aureus isolates from bovine mastitis in India. Comparative Immunology, Microbiology and Infectious Diseases 64:117-124. |
|
|
Tabaran A, Mihaiu M, Flaviu T, Colobatiu L, Reget O, Borzan MM, Dan SD (2017). First study on characterization of virulence and antibiotic resistance genes in verotoxigenic and enterotoxigenic E . coli isolated from raw milk and unpasteurized traditional cheeses in Romania. Folia Microbiologica 62(2):145-150. |
|
|
Tadesse HA, Gidey NB, Workelule K, Hailu H, Gidey S, Bsrat A, Taddele H (2018). Antimicrobial Resistance Profile of E . coli Isolated from Raw Cow Milk and Fresh Fruit Juice in Mekelle, Tigray, Ethiopia. Vetrinary Medicine International. |
|
|
Tirado C, Schmidt K (2001). WHO Surveillance Programme for Control of Foodborne Infections and Intoxications: Preliminary Results and Trends Across Greater Europe. Journal of Infection 43(1):80-84. |
|
|
Umaru AG, Kabir J, Umoh VJ, Bello M, Kwaga JKP (2013). Methicillin-resistant staphylococcus aureus (MRSA) in fresh and fermented milk in Zaria and Kaduna, Nigeria. International Journal of Drug Research and Technology 3(3):67-75. |
|
|
Vahedi M, Nasrolahei M, Sharif M, Mirabi AM (2013). Bacteriological study of raw and unexpired pasteurized cow's milk collected at the dairy farms and super markets in Sari city in 2011. Journal of preventive medicine and hygiene 54(2):120-123. |
|
|
Van Belkum A (2003). Molecular diagnostics in medical microbiology: yesterday, today and tomorrow. Current Opinion in Pharmacology 3(5):497-501. |
|
|
Van Boeckel TP, Brower C, Gilbert M, Grenfell BT, Levin SA, Robinson TP, Teillant A, Laxminarayan R (2015). Global trends in antimicrobial use in food animals. Proceedings of the National Academy of Sciences of the United States of America 112(18):5649-5654. |
|
|
Vanitha HD, Sethulekshmi C, Latha C (2018). An epidemiological investigation on occurrence of enterohemorrhagic Escherichia coli in raw milk. Veterinary World 11(8):1164-1170. |
|
|
Vendramin T, Kich DM, Molina RD, Souza CFVde, Salvatori RU, Pozzobon A, Bustamante-filho IC (2014). Molecular screening of bovine raw milk for the presence of Shiga toxin-producing Escherichia coli (STEC) on dairy farms. Food Science and Technology (Campinas) 34(3):604-608. |
|
|
Virpari PK, Nayak JB, Brahmbhatt MN, Thaker HC (2013). Study on isolation, molecular detection of virulence gene and antibiotic sensitivity pattern of Escherichia coli isolated from milk and milk products. Veterinary World 6(8):541-545. |
|
|
Vithanage NR, Dissanayake M, Bolge G, Palombo EA, Yeager TR, Datta N (2016). Biodiversity of culturable psychrotrophic microbiota in raw milk attributable to refrigeration conditions, seasonality and their spoilage potential. International Dairy Journal 57:80-90. |
|
|
Vranješ AP, Popović M, Jevtić M (2015). Raw milk consumption and health. Srpski Arhiv Za Celokupno Lekarstvo 143(1-2):87-92. |
|
|
Wu S, Duan N, Gu H, Hao L, Ye H, Gong W, Wang Z (2016). A Review of the Methods for Detection of Staphylococcus aureus Enterotoxins. Toxins 8(7). |
|
|
Zeinhom MMA, Abdel-Latef GK (2014). Public health risk of some milk borne pathogens. Beni-Suef University Journal of Basic and Applied Sciences 3(3):209-215. |
|
|
Zeinhom MMA, Abdel-Latef GK, Jordan K (2015). The Use of Multiplex PCR to Determine the Prevalence of Enterotoxigenic Staphylococcus aureus isolated from Raw Milk, Feta Cheese, and Hand Swabs. Journal of Food Science 80(12):M2932-M2936. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


