Full Length Research Paper

ABSTRACT
Microbial resistance to antimicrobials is spreading all over the world making it difficult to treat diseases effectively. This study aims to understand the mechanism of resistance towards the antimicrobials Ceftazidime, Moxifloxacin and Nalidixic acid, by using resistant Enterobacter spp. and sensitive Enterobacter spp. While there was complete inhibition of growth of sensitive Enterobacter spp. at 8 µg/ml Ceftazidime, 0.125 µg/ml of Moxifloxacin and 16 µg/ml of Nalidixic acid, resistant Enterobacter spp. even tolerated 256, 32 and 1536 µg/ml Ceftazidime, Moxifloxacin and Nalidixic acid respectively. Browthdilution method was used to determine the growth of organisms at different concentrations of antibacterial agents. Mechanism of resistance was found to be present in plasmid. Absence of Beta lactamase enzyme seems to be an important finding in this sudy. B. lactamase enzyme was checked with help of the acidometric and idometric method. Plasmid isolation and analysis was done by agarose gel electrophoresis. Successful curing of plasmid was carried out with 10% sodium dodecyl sulfate (SDS). When colonies after SDS treatment were tested, resistant strains were found which were later converted to sensitive ones. In this study, the resistant Enterobacter spp. executed resistance to three different classes of antimicrobials due to the resistance plasmid. The results obtained in this study support most of the previous study findings who contributed in this field. Therefore, it might be useful to recognize the resistance mechanism, and to determine the correct practicing of drug usage.
Key words: Antimicrobials, enterobacter, sensitive enterobacter, resistance mechanism, inhibition, plasmid.
INTRODUCTION
Pathogenic enteric bacteria that exhibit antimicrobial resistance are a widespread phenomenon and arguably constitute a global epidemic (Chau et al., 2007). Whilst the depth of knowledge regarding antimicrobial-resistant organisms isolated from patients with infection or circulation in the hospital environment is broad, less is known about antimicrobial resistant organisms that are disseminated in the community. Furthermore, little is known about the antimicrobial resistance patterns of community-acquired organisms that circulate in developing countries where antimicrobials are available without prior consultation with a physician. Quinolones and fluoroquinolones are groups of antimicrobial compounds that are commonly used for the treatment of many bacterial infections (Strahilevitz et al., 2007). However, multiple studies have highlighted that, in recent years, resistance to fluoroquinolones has increased globally, particularly in members of the Enterobacteriaceae (Wang et al., 2008).
Enterobacter species are motile aerobic gram negative bacilli belonging to the family Enterobacteriaceae. The major species are Enterobacter cloacae, Enterobacter aerogenes and Enterobacter agglomerans. They first achieved wide notoriety as pathogens in 1976 following a nationwide outbreak of septicemia in 378 patients at 25 hospitals resulting from contaminated intravenous solutions (Maki et al., 1976).
The genus Enterobacter was first proposed by Hormaeche and Edwards (1960). Enterobacter species are found in the natural environment in habitats such as water, sewage, vegetables and soil. Before the widespread use of antibiotics, Enterobacter species were rarely found as pathogens, but these organisms are now increasingly encountered, causing nosocomial infections such as urinary tract infections and bacteremia (Eickhoff et al., 1966). In addition, they occasionally cause community-acquired infections. In 1975 in the United States, Enterobacter species accounted for 4.6% of all pathogens causing infections, and accounted for 5.7% of all cases of primary bacteremia (Center for Disease Control, 1977).
Enterobacter species accounted for 5.9% of all nosocomial infections in U.S. hospitals and 6.3% of all nosocomial bacteremia (Centers for Disease Control, 1984). Enterobacter cloacae occur as a commensal organism in water, sewage, soil, meat, hospital environments, the skin, and in the intestinal tracts of humans and animals. Among 234 patients, the rate of stool carriage of E. cloacae on admission to the hospital was 2.6%. This rate increased to 4% after antibiotic therapy (Rose and Schreier, 1968). Like other enteric gram-negative rods, Enterobacter species cause a wide variety of nosocomial infections, including those affecting the lungs, urinary tract, intrabdominal cavity and intravascular devices. Enterobacter sakazakii causes neonatal sepsis with meningitis (Bar-Oz et al., 2001; Nazarowec-White and Farber, 1997). Sanders et al. (1996) described successful therapy with cefepime of 17 infections due to Enterobacter strains resistant to third generation cephalosporins. These patients had infections at a variety of sites. All patients responded clinically, and bacteriologic eradication was documented in 88%.
Enterobacter infections can include bacteremia, lower respiratory tract infections, skin and soft-tissue infections, urinary tract infections (UTIs), endocarditis, intra-abdominal infections, septic arthritis, osteomyelitis, central nervous system (CNS) infections and ophthalmic infections. Enterobacter infections can necessitate prolonged hospitalization, multiple and varied imaging studies and laboratory tests, various surgical and nonsurgical procedures, and powerful and expensive antimicrobial agents. With few exceptions, the major classes of antibiotics used to manage infections with these bacteria include the beta-lactams, carbapenems, fluoroquinolones, aminoglycosides and sulfamethoxazole and trimethoprim (SMZ-TMP). Because most Enterobacter species are either very resistant to many agents or can develop resistance during antimicrobial therapy, the choice of appropriate antimicrobial agents is complicated. Consultation with experts in infectious diseases and microbiology is usually indicated. In 2006, Paterson published a good review of resistance among various Enterobacteriaceae (Paterson, 2006).
Infections caused by antibiotic-resistant bacteria, especially the “ESKAPE” pathogens (Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aerugenosa and Enterobacter species), cause significant morbidity and mortality (Rice, 2008; Boucher et al., 2009). The incidence of nosocomial infections due to Enterobacter is rising, and broad resistance to third generation cephalosporins, penicillins and quinolones is an increasing problem. A number of agents remain effective for treatment. Among the beta-lactams, the fourth generation cephalosporins and carbapenems are the most attractive options. Aminoglycosides retain good activity but usually require combination with another agent. Quinolones are highly active against most strains, but emerging resistance is a major concern. Trimethoprim-sulfamethoxazole is under-utilized as therapy of Enterobacter infections (http://www.antimicrobe.org/b97.asp#r36).
Bacteria may be intrinsically resistant to more than one class of antimicrobial agents, or may acquire resistance by de novo mutation or via the acquisition of resistance genes from other organisms. Acquired resistance genes may enable a bacterium to produce enzymes that destroy the antibacterial drug and most resistant strains express efflux systems that prevent the drug from reaching its intracellular target required to kill or inhibit the infecting pathogen (Tenover and MacDonald, 2006). Quinolone resistance in Enterobacter is usually due to chromosomal genes that may upregulate efflux pumps (Nikaido, 2001) or confer resistance due to altered DNA gyrase (Dekitsch et al, 1999). Ceftazidime resistance was an inclusion criterion because of the strong association between qnr genes and plasmids carrying cephalosporinase genes. The ciprofloxacin MIC was the minimum expected for Enterobacteriaceae containing a qnr gene. (Gay et al., 2006). High prevalence of quinolone resistance determinants was identified, particularly the qnrS gene, in both - community and hospital-associated strains (Vien, 2009). The data suggest that intestinal commensal organisms are a significant reservoir for the dissemination of plasmid-mediated quinolone resistance in Ho Chi Minh City.
This study attempted to study the mechanism of resistance in Enterobacter species in relation with the earlier mentioned work. The phenomenon of resistance was assessed by the finding such as absence of B lactamase and resistance plasmid isolation from the resistant species of Enterobacter after the determination of MIC values for sensitive species and the tolerance of resistant species towards the antimicrobials at various concentrations above the MIC level determined for the sensitive species of Enterobacter. During plasmid isolation both the species of Enterobacter species had been processed but the sensitive Enterobacter spp. failed to show any band on the agarose gel when run along with the sample obtained from resistant species and the DNA ladder.
The dissimilar growing concentrations of the sensitive Enterobacter and resistant Enterobacter were found within different concentrations of Ceftazidime, Moxifloxacin and Nalidixic acid.
MATERIALS AND METHODS
Bacterial species and their cultivation
The strains of resistant and sensitive Enterobacter spp. were obtained from BAC TEST Laboratory Nashik, Maharashtra and stocked in this laboratory (Microbiology Laboratory, Department of Life Sciences, University of Mumbai). This particular pathogenic microorganism was selected for the study after testing various pathogens for their susceptibility towards the antimicrobials. These organisms were grown on a nutrient agar at 37°C and maintained at 5°C. The inoculum of MacFarland Turbidity standard was prepared in nutrient broth after subculturing, and was incubate for 48 h. The cells of Resistant and sensitive species of Enterobacter were cultured on nutrient agar (NA) (HiMedia, India) slants containing Beef extract 0.5 g; Peptone 2.5 g; Sodium chloride 2.5 g; Agar 15 g in a liter of distilled water. pH was maintained at 7.4, slants were incubated at 37°C for 24 h (Lankeshwar and Bagde, 2013) .
Detection of antimicrobial resistance and sensitivity in bacteria
Disc diffusion method
Antimicrobial susceptibility of resistant and sensitive Enterobacter Species was determined with the help of disc diffusion method of Kirby et al. (1994) with modification by the NCCLS (1999, 2001). The antimicrobial discs of Ceftazidime (30 mcg), Moxifloxacin (30 mcg), Nalidixic acid (5 mcg), and Muller Hinton (MH) Agar were procured from HiMedia (India). Zone of inhibition was measured after agar plates were incubated overnight and the zone of inhibition were measured with the help of standard scale (Standard strains have been used for reference).
Broth dilution method
The broth dilution method was used to determine the growth of organisms at different concentrations of antibacterial agents according to the guideline given by NCCLS (2001) protocol. The MH broth and the antimicrobials were purchased in powder form from HiMedia (India). The experiments were carried out in tubes and 100 ml Ehrlenmeyer flasks with side arm. These flasks were inoculated with 1.0 ml inoculum (Mac farland standard/103 CFU/ml) prepared in nutrient broth after subculturing 48 h incubation. Optical density of culture was measured at 540 nm. The final concentrations of antimicrobials tested were Ceftazidime 1.0, 2.0, 4.0, 8.0 µg/ml, Moxifloxacin 0.016, 0.032, 0.064, 0.125 µg/ml, Nalidixic acid 2.0, 4.0, 8.0, 16.0 µg/ml, for the sensitive strain and ceftazidime 32, 64, 128, 256 µg/ml, Moxifloxacin 4, 8, 16, 32 µg/ml, and Nalidixic acid 192, 384, 768, 1536 µg/ml for the resistant strain, respectively.
β lactamase activity
Detection of β lactamase was carried out by tube method and paper strip method of idometric test and acidometric test (Livermore and Brown, 2001), and B lactamase activity testing was done by qualitative plate test (George et al., 1983), nitrocefin test was not done due to unavailability of nitrocefin.
Plasmid isolation
Plasmid Isolation was conducted for separation and analysis of nucleic acid by alkaline lysis method (Sambrook et al., 1989). Overnight, bacterial culture in LB broth containing 50 ug/ml of ampicillin was chilled for an hour and centrifuged at high speed. Then the suspending/neutralizing buffer, lysis buffer, renaturation buffer, acidic potassium phosphate, phenol: chloroform solution were used to isolate the plasmid by this procedure. It was precipitated with chilled ethanol. The pellet was allowed to air dry for 15 to 20 min and stored with 50 ul of T10E1 Buffer at 4°C.
Spectrophotometric method
Qualitative determination of DNA was carried out with help of the spectrophotometric method given by Maniatis et al. (1982).
Agarose gel electrophoresis
Agarose gel electrophoresis of plasmid DNA was done according to the method of Sambrook et al. (1989). Plasmid DNA from Enterobacter resistant species was electrophoresed with DNA ladder in 1.0% agarose gel slab at 50 V in Tris Acetate buffer (TAE) running buffer (0.5M Tris base, 0.5M EDTA, 1 M Glacial acetic acid). Ethidium bromide incorporated in the agarose gel stained the plasmid DNA and the bands of plasmid seen under UV illumination were photographed. The size of plasmid was estimated by comparing with standard Supermix DNA ladder obtained from BangloreGenei, (Banglore, India). It consisted of DNA fragments ranging from 500 to 33500-24500 bp generated from number of propriotory plasmids and lambda DNA.
Plasmid curing
Elimination of resistance due to plasmid was carried out by using the sodium dodecyl sulphate (SDS) treatment described by Tomoeda et al. (1968). An overnight culture of resistant (R) cells in penassay broth ( 5 g/L Peptone, 1.5 g/L Yeast extract, 1.5 g/L Beef extract, 3.5 g/L Sodiuim chloride, 1 g/ Dextrose, 3.86 g/L potassium phosphate dibasic, 1.32g/L potassium phosphate monobasic. pH adjusted to 6.9) containing MIC concentration of antimicrobial (µg/ml) for sensitive species (103 cells/ml ) diluted in broth, and added to the tubes containing 10% (w/v) SDS and shaken at 37°C. After appropriate dilutions in saline cells were spread on nutrient agar and the sensitivity discs were placed on it.
RESULTS
Effect of antimicrobials on growth of sensitive and resistant species of Enterobacter
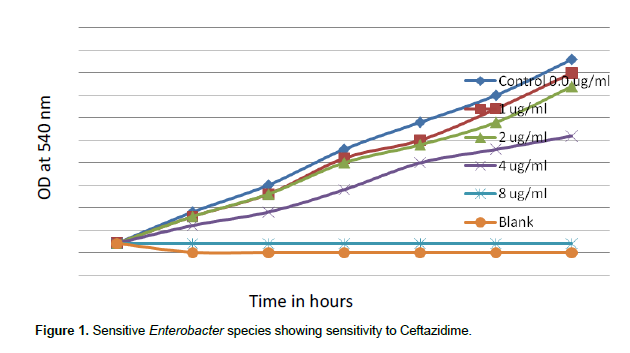
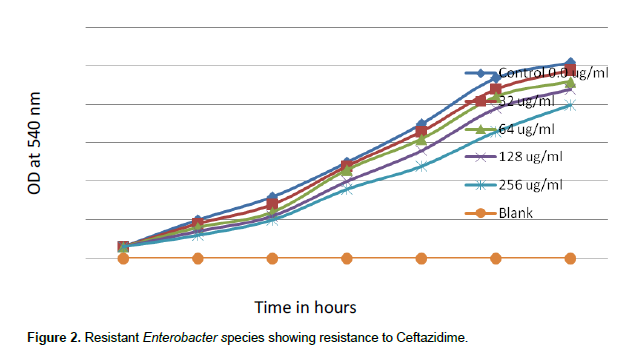
When concentrations of ceftazidime of 1.0, 2.0, 4.0 and 8.0 µg/ml were tested, sensitive species of Enterobacter showed MIC 8 µg/ml for ceftazidime. There was complete inhibition growth of sensitive Enterobacter spp. at 8 µg/ml ceftazidime (Figure 1). But when concentrations of ceftazidime 32, 64, 128 and 256 µg/ml were tested, resistant Enterobacter spp. tolerated 256 µg/ml Ceftazidime (Figure 2).
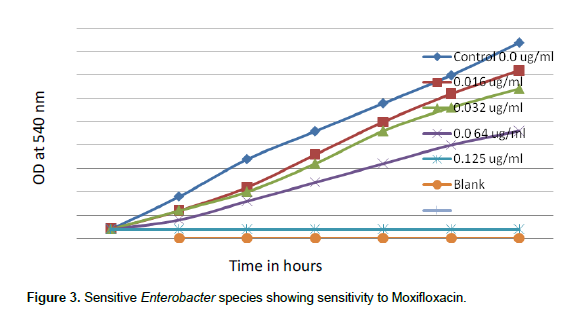
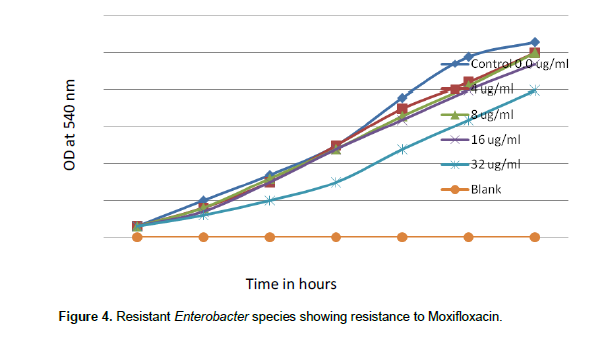
When concentrations of moxifloxacin 0.016, 0.032, 0.064 and 0.125 µg/ml were tested for sensitive species of Enterobacter, Moxifloxacin MIC was noted as 0.125 µg/ml. There was complete inhibition of growth of sensitive Enterobacter spp. at 0.1258 µg/ml of moxifloxacin (Figure 3), but when concentrations of moxifloxacin 4, 8, 16 and 32 µg/ml were tested, resistant Enterobacter spp. even tolerated 32 µg/ml moxifloxacin (Figure 4).
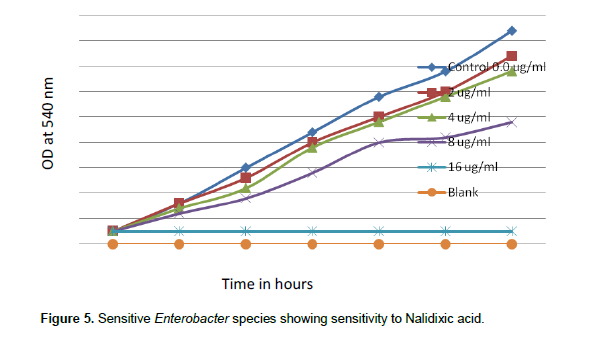
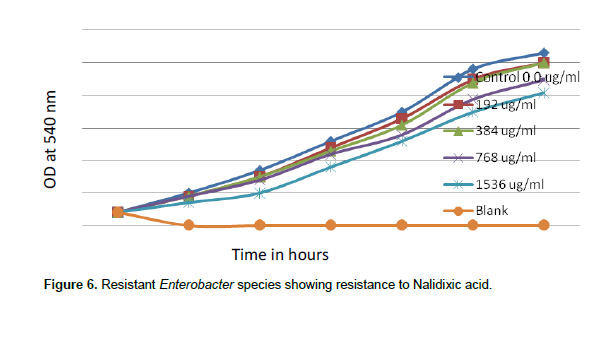
When concentrations of nalidixic acid 2.0, 4.0, 8.0 and 16.0 µg/ml were tested in the sensitive species of Enterobacter, nalidixic acid MIC was found to be 16 µg/ml. There was complete inhibition growth of sensitive Enterobacter spp. at 16 µg/ml of nalidixic acid (Figure 5). But when concentrations of nalidixic acid 192, 384, 768 and1536 µg/ml were tested, resistant Enterobacter spp. even tolerated1536 µg/ml Nalidixic acid (Figure 6).
The susceptibility of resistant and sensitive species of Enterobacter was noted as size of inhibition zone in mm. The sensitive Enterobacter showed zone of inhibition for ceftazidime -25 mm, for moxifloxacin –30 mm, and for Nalidixic acid –25 mm. The resistant species showed 0, 11, 12 and 6 mm, respectively size of inhibition zone in Nalidixic acid, Ceftazidime and moxifloxacin respectively. This clearly indicated the difference between the susceptibility of both species of Enterobacter (Table 1).
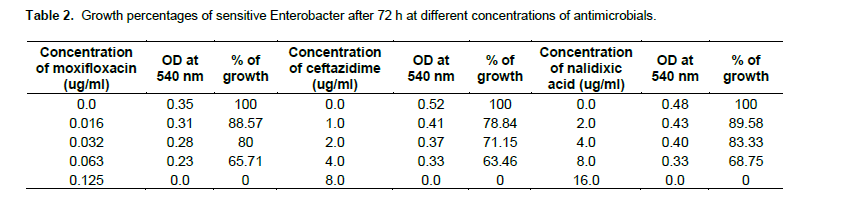
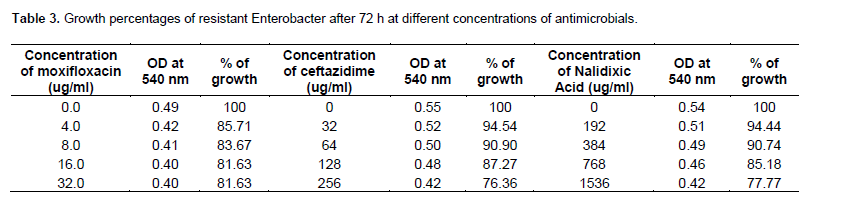
As shown in Table 2, the growing concentrations of sensitive Enterobacter (103 CFU/ml)in 4, 0.063, 8, µg/ml of ceftazidime, Moxifloxacin and Nalidixic acid, were found to be 63.46, 65.71 and 68.75% after 72 h. The growing concentrations of resistant Enterobacter at 103 CFU/mlin 256, 32 and 1536 µg/ml, of ceftazidime, Moxifloxacin and Nalidixic acid respectively, were found about 76.36, 81.63 and 77.77 after 72 h (Table 3). The higher most antimicrobial concentration tolerated by the resistant species is given here.
β Lactamase detection
As to beta lactamase detection, no decolorization occurred in 5 min in Idometric method within the tube or on the strip. Violet to Yellow color change was not observed within 5 min in Acidimetric method. No yellow color was observed around the colony within 60 min in qualitative plate test. Therefore, β Lactamase was not detected.
Plasmid isolation
In this experiment, sensitive and resistant Enterobacter were used for plasmid isolation. Plasmid was isolated successfully from the resistant species of Enterobacter and electrophoresed on agarose gel. Confirmation of plasmid was done with help of the DNA ladder.
Quantitative estimation of DNA
The purity of the isolated plasmid DNA was calculated as 1.65. The absorbance of plasmid of resistant Enterobacter species was 0.982. at 260 nm and 0.595 at 280 nm. The concentration of plasmid DNA from resistance Enterobacter calculated was 3928 gm. Therefore, the ratio of protein to extracted DNA was 1.65. Agarose gel electrophoresis of plasmid DNA isolated from Enterobacter species resistant to Ceftazidime, Moxifloxacin, Nalidixic acid Showed 4 different bands (Figures 6 and 7) and the Supermix DNA ladder showed bands of different molecular weight, while the sensitive species of Enterobacter did not show any band in lane 3, 4 and 6 (Figure 7).
.png)

The plasmid DNA has been run along with the DNA ladder for the purpose of conformation only. There are more than 1 plasmid DNA found near the bands of ladder having molecular weight of 15000 bp (at lower side), and molecular weight more than 33500 bp (both at upper side).
Plasmid curing
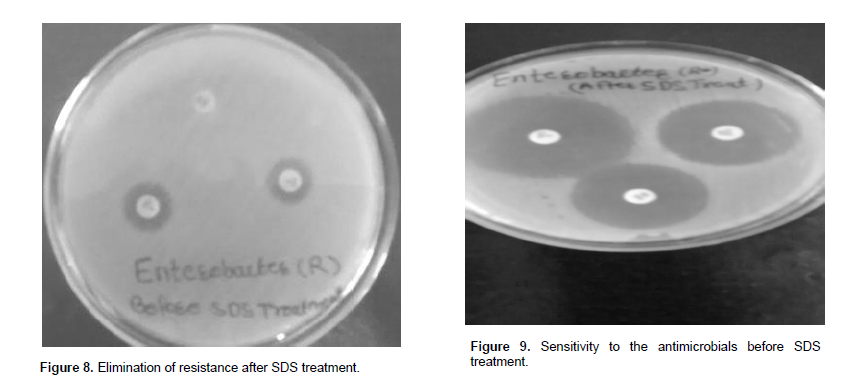
In this experiment, the cells of Enterobacter carrying resistance plasmid were treated with 2 to 10% concentration of sodium dodecyl sulphate. The treatment of sodium dodecyl sulphate was found to be effective to turn the resistant cells into susceptible ones. The resistant species showed up to 51 to 27% growth at 2 to 10% SDS concentration as compared to control set. After SDS treatments, resistant strains were analyzed at interval for the presence or absence of drug resistance against different concentration of antimicrobials. The resistance was lost and the resistant species completely changed into the sensitive species (Figure 8 and 9).
DISCUSSION
The present study has exposed the mechanism of resistance towards the antimicrobials. There was complete inhibition of growth of sensitive Enterobacter spp. at 8 µg/ml Ceftazidime, 0.125 µg/ml of Moxifloxacin, and 16 µg/ml of Nalidixic acid, while the resistant Enterobacter spp. even tolerated 256, 32 and 1536 µg/ml of Ceftazidime, Moxifloxacin and Nalidixic acid respectively. Different ranges of concentrations of the earlier mentioned antimicrobials have been incorporated in the MIC study after determination of the MIC value of each antimicrobial through several trials of experiment in various batches. Evaluation of a wide range of antibiotics tested against E. cloacae, Enterobacter hormaechei and Enterobacter asburiae strains, have provided a database for their natural susceptibility (Stock et al., 2001).
In this present study, the growing concentrations were found 76.36, 81.63 and 77.77% at 103 CFU/ml after 72 h for 256, 32, 16 and 1536 µg/ml of ceftazidime, Moxifloxacin, and Nalidixic acid, respectively in resistant Enterobacter. The growing concentrations of sensitive Enterobacter ( 103 CFU/ml) were found up to 63.46, 65.71 and 68.75% after 72 h in 4, 0.063, 4, and 8 µg/ml of ceftazidime, Moxifloxacin, and Nalidixic acid. These findings can be compared with the findings of other studies. The antimicrobial-resistance rates of qnr-positive strains to Ciprofloxacin, Levofloxacin, Norfloxacin, Nalidixic acid, and Moxifloxacin were 51.1, 46.8, 46.8, 74.5 and 53.2%, respectively (Haeng et al., 2011). Most isolates of the E. cloacae complex are susceptible to fluoroquinolones, trimethoprim/sulfamethoxazole, chloramphenicol, aminoglycosides, tetracyclines, piperacillin-tazobactam and carbapenems, while they are intrinsically resistant to ampicillin, amoxicillin, amoxicillin–clavulanate, first-generation cephalosporins and cefoxitin owing to the production of constitutive AmpC β-lactamase. In particular, fosfomycin seems to have a different activity against all species, because E. cloacae and E. asburiae are both naturally susceptible and resistant, while E. hormaechei is only naturally sensitive (Stock et al., 2001).
Plasmid was not isolated from sensitive strain, but there was successful isolation of plasmids from resistant species of Enterobacter which supported to state that the mechanism of resistance was plasmid mediated. Similar observations were made in earlier studies where successful isolation of one or more plasmids of various molecular sizes was reported from resistant species, and plasmid was not isolated from sensitive strains (Ghosh et al., 1997; Lankeshwar and Bagde, 2004, 2008, 2013).
The result of agarose gel electrophoresis is clearly showing the presence of 3 different bands into the second picture in Figure 9. It could be specified as nicked circle plasmid DNA, linear plasmid DNA and supercoiled plasmid DNA (Birnboim and Doly, 1979; Ish-Horowicz and Burke, 1981).
The more recent discovery and rapid dissemination of plasmid-mediated quinolone resistance (PMQR) genes has further highlighted the problem of quinolone (Nalidixic acid) and fluoroquinolone (Moxifloxacin) resistance and increased the study understanding of resistance mechanisms associated with these antibacterial compounds (Robicsek et al., 2006). Although quinolone resistance results mostly from chromosomal mutations in Enterobacteriaceae, it may also be mediated by plasmid-encoded Qnr determinants. Qnr proteins protect DNA from quinolone binding and compromise the efficacy of quinolones such as Nalidixic acid (Nordmann and Laurent, 2005). The experiment done in this study is in agreement with this statement, as the resistant species of Enterobacter found to be resistant to different classes of antimicrobials namely, third generation cephalosporin (Ceftazidime), quinolone (Moxifloxacin) and fluroquinolone (Nalidixic acid).
In present study Enterobacter carrying resistance plasmid was treated with 2 to 10% concentration of sodium dodecyl sulphate. The treatment of sodium dodecyl sulphate was found to be effective to turn the resistant cells into susceptible ones. The resistance was lost and the resistant species completely changed into the sensitive species. This was very much in agreement to earlier studies of plasmid elimination using SDS (Tomoeda et al., 1968; Pan-Hau et al., 1981; Lankeshwar and Bagde, 2008, 2013). Reportedly also, acriflavine was used for elimination of resistance to penicillin in S. aureus (Hashimoto et al., 1964). However, antimicrobial susceptibility testing for clinically significant strains is highly recommended, as resistance to antibacterial agents may be strain dependent (Max et al., 2011).
CONCLUSION
This study revealed the mechanism of resistance on molecular basis towards the antimicrobials Ceftazidime, Moxifloxacin and Nalidixic acid as the resistance plasmid was isolated in resistant species of Enterobacter and not from the Sensitive. The mechanism of resistance to Ceftazidime in two clinical isolates of Enterobacter cloacae that emerged during therapy with broad-spectrum beta-lactam antibiotics was studied earlier (Quinn et al., 1987). Plasmid mediated fluroquinolones resistance was also reported by Maria et al. (2012) due to Qnr-mediated topoisom protection with enzyme type Qnr(A, B, S, C, D) in E. cloacae and E. hormaechei. Ceftazidime resistance was an inclusion criterion because of the strong association between qnr genes and plasmids carrying cephalosporinase genes (Gay et al., 2006).
It is reported that the maximum resistance was seen against Ceftazidime (74.8%) followed by Cefotaxime (70.6%). In ESBL, producing bacteria was (59.6%) mostly in K. pneumoniae (68.8. %) followed by E. coli (65.0 %).
ESBL producing bacteria showed maximum resistance to Ceftazidime (95.4%), followed by Cefotaxime (94.6%), while minimum resistance was seen with Imipenem (0%), followed by Piperacillin/Tazobactam (3.8%) and Cefepime (7.7%) (Ahmed et al., 2013).
In accordance with these findings, the Ceftazidime resistance in this study can be specified with Enterobacter spp. Over the past years, the development and application of molecular diagnostic techniques has initiated a revolution in the diagnosis and monitoring of infectious diseases (Yi-Wei et al., 1997). This study might contribute to understand the status and phenomenon of resistance in pathogens, efficacy of drugs, relation with MDR and finally the correct usage of antimicrobials.
CONFLICT OF INTEREST
The authors have not declared any conflict of interest.
REFERENCES
|
Ahmed OB, Alfadel OO, Asghar AH, Elhassan MM (2013). Increasing prevalence of ESBL-producing Enterobacteriaceae in Sudan community patients with UTIs. Egypt. Acad. J. Biol. Sci. 5(1):17-24. |
|
|
Bar-Oz B, Preminger A, Peleg O, Block C, Arad I (2001). Enterobacter sakazakii infection in the newborn. Acta Paediatr. 90(3):356-358. |
|
|
Birnboim HC, Doly J (1979). A rapid alkline procedure for screening recombinant plasmid DNA. Nucleic Acids Res. 7:1513-1523. |
|
|
Boucher HW, Talbot GH, Bradley JS (2009). Bad bugs, no drugs: no ESKAPE! An update from the Infectious Diseases Society of America. Clin Infect. Dis. 48:1-12. |
|
|
Centers for Disease Control (1977). National nosocomial infection study report: annual summary 1975. Atlanta. |
|
|
Centers for Disease Control (1984). Nosocomial infection surveillance. CDC Surveillance Summaries 33:17SS– 29SS. E. |
|
|
Chau TT, Campbell JI, Galindo CM, Van Minh Hoang N, Diep TS, Van VinhChau N, Tuan PQ, Page AL, other authors (2007). Antimicrobial drug resistance of Salmonella entricserover Typhi in Asia and molecular mechanism of reduced susceptibility to the fluoroquinolones. Antimicrob. Agents Chemother. 51:4315-4323. |
|
|
Dekitsch C, Schein R, Markolpulos E, Kuen B, Graninger W, Georgopoulos A (1999). Analysis of mutations to gyrA in quinolone-resistant clinical isolates of E cloacae. J. Med. Microbiol. 48:73-77. |
|
|
Eickhoff TC, Steinhauer BW, Finland M (1966). The Klebsiella-Enterobacter-Serratia division: Biochemical and serologic characteristics and susceptibility to antibiotics. Ann. Int. Med. 65:1163-1179. |
|
|
Gay KA, Robicsek J, Strahilevitz CH, Park GA, Jacoby TJ, Barrett F, Medalla TM, Chiller, Hooper DC (2006). Plasmid mediated quinolone resistance in non-Typhi serotypes of Salmonella enterica. Clin. Infect. Dis. 43(3):297-304. |
|
|
Ghosh S, Mahaptra NR, Banerjee PC (1997). Metal resistance in Acidocella strain and plasmid mediated transfer of this character to Acidophilliummultivorum and Escherichia coli. Appl. Environ. Microbiol. 63:4523-4527. |
|
|
Hashimoto H, Kono K, MitsuhashiS (1964). Elimination of penicillin resistance of Staphylococcus aureus by treatment with acriflavine. J. Bacteriol. 88:261-262. |
|
|
Hormaeche E, Edwards PR (1960). A proposed genus Enterobacter. Int. Bull. Bacteriol. Nomencl. Taxon. 10:71-74. |
|
|
Ish-Horowicz D, Burke JF (1981). Rapid and efficient cosmid cloning. Nucleic acids Res. 9:2989-2998. |
|
|
Kirby WM (1994). Extraction of a highly potent penicillin inactivator from penicillin resistant staphylococci. Science 99:452-455. |
|
|
Lankeshwar N, Bagde US (2004). Mechanism of Antibiotic Resistance in Pathogenic Shigelladysenteriae. Asian J. Microbiol. Biotechnol. Environ. Sci. 6(2):285-290. |
|
|
Lankeshwar N, Bagde US (2013). Mechanism of Ciprofloxacin resistance in Shigelladysenteriae. Afr. J. Biotechnol. 12(41):6021-6025 |
|
|
Lankeshwar N, Bagde US (2008). Mechanism of Drug resistance in Staphylococcus aureus. Asian J. Microbiol. Biotechnol. Environ. Sci. 10(4):829-834 |
|
|
Livermore DM, Brown DFJ (2001). Detection of Beta-lactamase mediated resistance. J. Antimicrob. Chemother. 48:59-64. |
|
|
Maki DG, Rhame FS, Mackel DC, Bennett JV (1976). Nationwide epidemic of septicemia caused by contaminated intravenous products. Am. J. Med. 60:471-485. |
|
|
Maniatis T, Frisch EF, Sambrook J (1982). Large isolation of Plasmid DNA. In: Molecular Cloning: A laboratory manual ed. Maniatis T, Frisch EF and Sambrook J. Cold Spring Harbor Laboratory Press N.Y. 0879691360(pbk). pp. 86-96. |
|
|
National Committee for Clinical Laboratory Standards (NCCLS). (1999). Document M 100-S22 Performance standards for antimicrobial susceptibility testing. 22nd informational supplement. NCCLS, Wayen. pp. 44-49. |
|
|
National Committee for Clinical Laboratory Standards (NCCLS) (2001). Document M 100-S11. Performance standards for antimicrobial susceptibility testing. 9th informational supplement. NCCLS, Wayen. p. 122. |
|
|
Nazarowec-White M, Farber J (1997). E sakazakii: a review. Int. J. Food Microbiol. 34:103-113. |
|
|
Nikaido H (2001). Preventing drug access to targets: cell surface permeability barriers and in bacteria. Semin. Cell Dev. Biol. 12:215-223. |
|
|
Nordmann P, Poirel L (2005). Emergence of plasmid-mediated resistance to quinolones in Enterobacteriaceae. J. Antimicrob. Chemother. 56:463-469. |
|
|
Pan-Hou HS, Nishimoto M, Imura N (1981). Possible role of membrane protein in mercury resistance of Enterobacteraerogenes. Arch. Microbiol. 130:93-95. |
|
|
Paterson DL (2006). Resistance in gram-negative bacteria: enterobacteriaceae. Am. J. Med. 119(6 Suppl 1):S20-8; discussion S62-70. |
|
|
Quinn JP, DiVincenzo CA, Foster J (1987). Emergence of resistance to ceftazidime during therapy for Enterobacter cloacae infections. J. Infect. Dis. 155(5):942-947. |
|
|
Rice LB, (2008). Federal funding for the study of antimicrobial resistance in nosocomial pathogens: no ESKAPE. J. Infect. Dis. 197:1079-1081. |
|
|
Robicsek A, Jacoby GA, Hooper DC (2006). The worldwide emergence of plasmid-mediated quinolone resistance. Lancet Infect. Dis. 6:629-640. |
|
|
Rose HD, Schreier J (1968). The effect of hospitalization and antibiotic therapy on the Gram-negative fecal flora. Am. J. Med. Sci. 255:228-236. |
|
|
Sambrook J, Fritsch EF, Mantiatis T (1989). Molecular cloning; a laboratory manual. Cold Spring Harbor Laboratory Press, NY, U.S. A. |
|
|
Sanders WE, Tenney JH, Kessler RE (1996). Efficacy of cefepime in the treatment of infections due to multiply resistant Enterobacter species. Clin. Infect. Dis. 23:454-461. |
|
|
Stock I, Grüger T, Wiedemann B (2001). Natural antibiotic susceptibility of strains of the Enterobacter cloacae complex. Int. J. Antimicrob. Agents 18(6):537-545. |
|
|
Strahilevitz J, Engelstein D, Adler A, Temper V, Moses AE, Block C, Robicsek A (2007). Changes in qnr prevalence and fluoroquinolone resistance in clinical isolates of Klebsiella pneumonia and Enterobacter spp. collected from 1990 to 2005. Antimicrob. Agents Chemother. 51:3001-3003. |
|
|
Tenover FC, MacDonald LC (2005). Vacomycine-resistant Staphylococci and enterococci: epidemiology and control. Curr. Opin. Infect. Dis. 18:300-305. |
|
|
Tomoeda M, Inuzulka M, Kubo N, Nakamura S (1968). Effective elimination of drug resistance and sex factors in Escherichia coli by sodium dodecyl sulfate. J. Bacteriol. 95:1078-1089. |
|
|
Vien LTM, Baker S, Le TPT, Thuy CT, Nga T, Hoang NVM, Campbell JL, Yen LM, Hieu NT, VinhChau NV, Farrar J, Schultsz C (2009). High prevalence of plasmid-mediated quinolone resistance determinants in commensal members of the Enterobacteriaceae in Ho Chi Minh City, Vietnam. J. Med. Microbiol. 58:1585-1592. |
|
|
Wang A, Yang Y, Lu Q, Wang Y, Chen Y, Deng L, Ding H, Deng Q, Wang L, Shen X (2008). Occurrence of qnr-positive clinical isolates in Klebsiellapneumoniae producing ESBL or AmpCtype b-lactamase from five pediatric hospitals in China. FEMS Microbiol. Lett. 283:112-116. |
|
|
Yi-Wei Tang, Gary W. Procop, Persing DH (1997). Molecular diagnostics of infectious diseases. Clin. Chem. 43(11):2021-2038. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


