Review

ABSTRACT
Neuropathic pain is the consequence of abnormal processing in the peripheral or central nervous system (CNS) elicited by neuronal injury. Due to its heterogeneous nature and important unwanted adverse effects of the commonly prescribed psychoactive drugs like benzodiazepines (BDZ), the treatment of neuropathic pain has remained a challenge for the scientific community. Flavonoids initially isolated from plants and used as tranquilizers in Folkloric medicine, have been reported to possess selective affinity for BDZ binding site. These positive ionotropic modulators of γ-amino butyric acid-A (GABAA) receptors enhance the chloride ion flux and provide a strong inhibitory effect. Therefore, for the treatment of central nervous system-related disorders such as neuropathic pain, these selective GABAA receptor modulators stand amongst the strongest candidates. This review provides an update on research development that has confirmed the activity of different flavonoids on GABAA receptors.
Key words: Neuropathic pain, flavonoid, γ-amino butyric acid-A (GABAA) receptors, animal models neuropathy.
Abbreviation: AD, Alzheimer’s disease; ASCIs, acid-sensing ions channel; BDZs, Benzodiazepines; CNS, central nervous system; COX-2, Cycloxygenase-2; CRPS II, Complex Regional Pain Syndrome II; DRG, dorsal root ganglion; GABA, γ-amino butyric acid; IASP, International Association for the Study of Pain; ISSVD, International Society for the Study of Vaginal Disease; MAPK, mitogen-activated protein kinase; NeuPSIG, Neuropathic Pain Special Interest Group; NMDA, N-methyl-D-aspartate; PD, Parkinson’s disease; TNF-α, tumor necrosis factor- α; TRPV, transient receptor potential.
INTRODUCTION
Formerly, neuropathic pain was defined by International Association for the Study of Pain (IASP) as “pain initiated or caused by a primary lesion or dysfunction of the nervous system”; but recently it has been revised as “pain arising as a direct consequence of a lesion or disease affecting the somatosensory system” by the IASP neuropathic pain special interest group (NeuPSIG) (Treede et al., 2008). So, this pain has been primarily attributed to be a corollary of a disease upsetting the ‘somatosensory system’ rather than the ‘nervous system’ in the revised definition, thus, further clarifying its origin. Under normal conditions, the somatosensory system is involved in the diffusion of noxious information to the central nervous system. Hence lesion of the somatosensory system can not only stop the innervations of nerve cells but can also result in pain with or without sensory hypersensitivity episode in the painful area (Jensen, 2006).
A lesion in the somatosensory system can manifest as positive sensory symptoms or negative sensory symptoms. The negative sensory symptoms arise due to fractional or entire loss of input to the nervous system; whereas, the positive sensory symptoms arise due to regeneration and disinhibition of the nerve cells, as a result of reduced sensory input. The positive symptoms can be either spontaneous or stimulus-evoked.
Paraesthesias (tingling or ant crawling sensations over the skin), spontaneous ongoing or shooting pain (stimulus independent) and electric shock-like sensations are included in spontaneous positive symptoms; whereas, hyperalgesia and allodynic (further classified according to the static and dynamic nature of the stimulus) pain are stimulus-evoked positive symptoms of neuropathy (Rasmussen et al., 2004). Negative symptoms of neuropathic pain includes hypoesthesia (reduced sensations to non-painful/innocuous stimuli), pallyhypoesthesia (reduced sensations to vibration), hypoalgesia (abridged impression to noxious stimuli) and thermohypoesthesia (abridged impression to cold/warm stimuli) (Jensen, 2006).
Numerous controlled studies have demonstrated potential effectiveness of opioids, tricyclic anti-depressants, gabapentin, pregabalin, phenytoin, lamotrigine, dextromethorphan, tramadol and mexiletine for painful sensory neuropathy (Harden, 1999; Attal, 2001). However, these therapies result in a 30–50% reduction in pain and are often restricted due to significant side effects, with sedation being the most dominant.
PATHOPHYSIOLOGICAL MECHANISMS OF NEUROPATHIC PAIN
Experimental work in rodents has provided a broadened picture of the pathophysiological mechanisms that generate neuropathic pain. It involves both peripheral and central mechanisms. The peripheral sensitization is carried out via unmyelinated C- and thinly myelinated Aδ- primary afferent neurons that normally elicit the pain sensation by responding to noxious stimuli. However, the peripheral nerve lesions sensitize these neurons advancing into spontaneous activity. Moreover, these lesions cause dramatic changes on the cellular and molecular levels triggering the nerve cells (Figure 1) (Baron, 2006).

An increased mRNA expression for voltage gated sodium channels in the primary afferent neurons is considered to be responsible for an ectopic spontaneous activity after a nerve injury. This phenomenon may result in the clustering of these channels that lowers the action potential threshold leading to hypersensitivity. Hence, sodium channel blockers like lidocaine produce pain relief in neuropathic pain via this mechanism (Lai et al., 2003).
Up regulation of a variety of receptor proteins also result from peripheral nerve injury. They are present at the membrane of the primary afferents and are only slightly uttered in physiological circumstances. Vanilloid receptors (TRPV 1) being one of them are involved in the sensing of noxious heat greater than 43ºC (Patapoutian et al., 2003), whereas, TRPM8 receptors have been identified as cold and menthol-sensitive raised by temperature in the range of 8-28ºC. TRPM8 receptor is revealed in small diameter DRG (dorsal root ganglia) neurons (McKemy et al., 2002). A nerve injury may result in either up regulation or gating of this channel leading to peripheral sensitization of C-nociceptors that causes cold hyperalgesia (Wasner et al., 2004).
Acid-sensing ion channels (ASCIs) are believed to be participating in static mechanical hyperalgesia (Price et al., 2001). On the other hand, α1- and α2- adrenoceptors located on the cutaneous afferent fibers also play a pronounced function in the hypersensitivity from nerve injury (Baron et al., 1999). Adrenergic sensitivity has been widely expressed in post herpetic neuralgias, complex regional pain syndromes II (CRPS II) and post-traumatic neuralgias; whereas, in polyneuropathies no sensitivity in the primary afferent neurons has been reported (Uphoff and Binder, 2006). Thus, temperature-induced and sympathetically-induced pain could be treated by blocking their respective receptors on nociceptive neurons.
An ectopic activity is also induced by inflammation in both wounded and adjoining normal primary afferent nociceptors triggered by a nerve lesion that release proinflammatory cytokines, especially TNF-α (Sommer, 2003). Deep proximal and paroxysmal pain are pronounced features in the patients diagnosed with peripheral neuropathies e.g., HIV neuropathy. Nerve biopsy specimens of such patient have shown an increased concentration of COX-2 and proinflammatory cytokines (Table 1) (Lindenlaub and Sommer, 2003).

CNS constitutes specific anatomical links with the spinal cord, brain stem, thalamus and the cortex. These connections link sensations generated in the high threshold primary afferents with the cortical regions of the central nervous system, that further process it into ultimate painful sensations (Woolf, 2011). The ongoing hyperactivity generated by injured nerves serves as a trigger for central sensitization and simulates activity-dependent synaptic flexibility occurring within the cortex. Diverse synaptic modulators, changes in ion channel kinetics, excitatory amino acid, increased bulk of ionotropic receptors and pre- and post-synaptic activation of kinases are involved in central sensitization.
Most of the patients presenting with peripheral and all with central neuropathy show predominant synaptic facilitation contributing to allodynia and hypersensitivity (Campbell and Meyer, 2006). Peripheral nerve injury causes “pre-synaptic changes” including modifications in the synthesis of neurotransmitters, neuromodulators and in the density of calcium channels (Hendrich et al., 2008). On the contrary, increased receptor density as a result of enhanced synthesis of ion channels and scaffold proteins and phosphorylation of N-methyl-D-aspartate (NMDA) subunits occur because of “post-synaptic changes” (Cheng et al., 2008). These changes also contribute to an abnormal expression of Nav 1.3 (Hains et al., 2004)and the mitogen-activated protein kinase system (MAPK) (Ji and Woolf, 2001). The role of pathologically sensitized C-fibers which sensitizes neuropeptide substance P and spinal dorsal horn via glutamate release cannot be overlooked. The released glutamate shows an excitatory effect by acting upon post-synaptic NMDA leading to central sensitization (Ultenius et al., 2006). An appreciably abundant evidence reveal the involvement of loss of tonic GABA-conciliated inhibition and increased excitatory neurotransmitters resulted in an initiation of central sensitization, ending in peripheral hypersensitivity, that is, allodynia and hyperalgesia (Knabl, Witschi et al., 2008). Once this sensitivity is developed, the usually innocuous tactile stimuli could activate Aδ and Aβ low threshold mechanoceptors (Tal and Bennett, 1994).
ROLE OF GABA IN NEUROPATHIC PAIN
In the brain, γ-amino-butyric acid (GABA) tends to be the most abundant inhibitory neurotransmitter regulating different physiological characters like sleep, anxiety, memory formation, reward, etc (Zeilhofer et al., 2009)and also control the action of excitatory neurons in the CNS, facilitating a uniform flow of information and therefore, maintaining the homeostasis of neural circuits. Melzack and Wall (1967) previously reported the role of inhibitory neurons in the spinal dorsal horn involved in controlling pain transmission from the periphery to higher levels of the brain (Melzack and Wall, 1967). Later on, GABA was confirmed to be one of the foremost inhibitory neurotransmitters in the spinal dorsal horn (Yaksh, 1989).
After release from the pre-synaptic neurons, GABA act postsynaptically upon 3 major classes of receptors namely; GABAA and GABAC receptors that are ligand-gated ion channels and GABAB that are G protein-coupled channels (Gavande et al., 2011). Chemically ionotropic GABAA receptors are formed of transmembrane protein complexes and comprise 5 heteropentameric subunits. In the human brain, α1β2γ2 subunit is considered to be the most dominant one (Wafford, 2005). Upon activation by GABA, the membrane permeability to chloride and carbonate ions increases causing a net inward flow of anions and a consequent hyperpolarization. This hyperpolarizing post-synaptic response is termed as inhibitory post-synaptic potential (Semyanov et al., 2003).
Physiologically, GABA-releasing interneurons, imposes a strong inhibitory control over the dorsal horn neurons. The loss of these neurons could further increase the central sensitization, e.g. as reported in partial peripheral nerve injury models, in which the injury resulted in the reduced spinal GABA release along with decreased GABA-synthesizing enzyme glutamic acid decarboxylase in rodents (Moore et al., 2002), advancing into spontaneous pain and hyperexcitability manifested as allodynia and hyperalgesia.
In diseased states, an increased excitation occurs that is attributed to either a massive loss of GABAergic interneurons or deterioration of interneurons as consequence of depletion of their sensory excitatory inputs or receptors. This imbalance could culminate into many neurological and psychiatric disorders including epilepsy, schizophrenia, neuropathic pain, Alzheimer’s (AD) and Parkinson’s disease (PD) (Tyson and Anderson 2014).
Hence, the collaborative function of excitatory and inhibitory neurons plays a vital role in controlling various brain activities.
As discussed earlier, central and peripheral sensitization due to nerve injury could result in complex neuropathic pain. Besides other mechanisms, the loss of GABAergic interneurons is now considered to be one of the major contributor to such persistent pain states (Bráz et al., 2012). Recent evidence suggest that the deletion of particular GABA subunits or pharmacological blockade of GABAergic neurotransmission in the spinal cord resulted in hyperalgesia and allodynia (Gwak et al., 2006; Jergova et al., 2012). Similarly, electrophysiological studies have revealed that GABAA receptor inhibition could induce an embellished behavioral reaction to an innocuous mechanical stimulus (Hwang and Yaksh, 1997), consistent with the reported high levels of GABAergic receptors in the dorsal horn (Persohn et al., 1991). Moreover, the impaired GABAergic system in the animals presenting with chronic neuropathic pain propose the practical link between such pain and spinal inhibitory neurotransmission (Figure 2) (Zeilhofer, 2008). Therefore, restoration of spinal inhibitory neurotransmission can be a valuable pharmacological advancement in the treatment of neuropathic pain.

GABAA receptor agonists related antinociceptive effect has been attributed to the stimulation or blockade of other neurotransmitters as well (McCarson and Enna, 2014). Furthermore, the role of central GABA in opioid-mediated antinociception is well documented (Ossipov et al., 2010). Thus, GABA receptor agonists may play a vital role in treating acute and chronic pain (McCarson and Enna, 2014).
In this regard, GABAA receptor agonists muscimol and isoguvacine are reported to reverse nerve injury-induced tactile allodynia (Hwang and Yaksh, 1997). These receptors are closely linked to the large diameter afferents involved in innocuous sensation (Price et al., 1984; Sivilotti and Woolf, 1994; Reeve et al., 1998; Ataka et al., 2000; Riley et al., 2001; Turner, 2003). Great body of evidence suggests that a decreased spinal GABA plays a vital role in inducing and maintaining neuropathic pain. Behavioral and pharmacological studies have shown that a continuous or single intrathecal dose of GABA to the spinal cord or the implantation of GABA releasing cells decrease the signs of neuropathic pain (Eaton et al., 1998, 1999; Stubley et al., 2001; Malan et al., 2002). Moreover, blocking the spinal GABAA receptors results in aggravating peripheral nerve injury associated hyperalgesia (Yamamoto and Yaksh, 1993).
On the contrary, benzodiazepines, and positive allosteric modulators at GABAA receptors are widely used drugs not only in anxiety, convulsions and sleep disorders but also as analgesic if administered intrathecally (Tucker et al., 2004). Despite its analgesic potential, its use in pain relief is restricted due to the myriad effects like sedation. Hence, there is an increased need of research in the field of GABAergic modulators that could play a pronounced role in the attenuation of neuropathic pain.
FLAVONOIDS
Flavonoids are widely distributed in vegetables, grains, fruits, barks, roots, flowers, stems, tea, etc. Chemically, they are polyphenolic compounds comprising of a diphenyl propane skeleton (C-6-C-3-C-6), with two aromatic rings and an oxygen-containing heterocyclic benzopyran ring, labeled as fused aromatic ring A, the benzopyran ring adjacent to A is ring C and a phenyl ring B (Middleton, 1998).
Long before the isolation of flavonoids, the natural products bearing them were known for their healthful effects. The decreased mortality rate associated with the use of red wine (constitute flavonoids) was observed in Mediterranean populations that produced an inclination in the flavonoid research, that was further confirmed by epidemiological studies, portraying the cardio protective role of dietary flavonoids in coronary heart disease (Formica and Regelson, 1995; Groot and Rauen, 1998).
Similarly, a new substance isolated from oranges in 1930, was thought to be a vitamin and was called vitamin P, which was later on confirmed to be a flavonoid, that is, rutin which led to an increase in the isolation and study of the mechanisms of various individual flavonoids. A long history of plant species exists that constitute flavonoids and exhibit CNS effects and were used as folk medicines in Europe. Feverfew (Tanacetum parthenium L. Asteraceae) was used as prophylactic agent in migraine and Chamomile flowers (Matricaria recutica L. Asteraceae) as a tranquillizer for centuries, with both found to be containing apigenin as an active constituent via bioassay-guided fractionation studies (Jäger et al., 2009). Furthermore, Linden flowers (Tilia sp. Tiliaceae) were known for their sedative effects and Heather (Calluna vulgaris [L] Hull. Ericaceae) as nerve calming remedy, with both shown to have quercetin and kaempferol flavonoids as active components (Aguirre-Hernández et al., 2010).
Flavonoids can be categorized into ten groups (Table 2) (Peterson and Dwyer, 1998). Flavone, isoflavone and flavanol contains a double bond in the ring C, making the fused A-C ring system planar, whereas, the other classes lack this double bond and instead have C2 and C3 chiral centers that are located at each end of the plane ring A (Figure 3) (Jager and Saaby, 2011). Besides the isolation of natural flavonoids, various semi-synthetic and synthetic derivatives have been synthesized and screened for their therapeutic potential (Cushnie and Lamb, 2005).

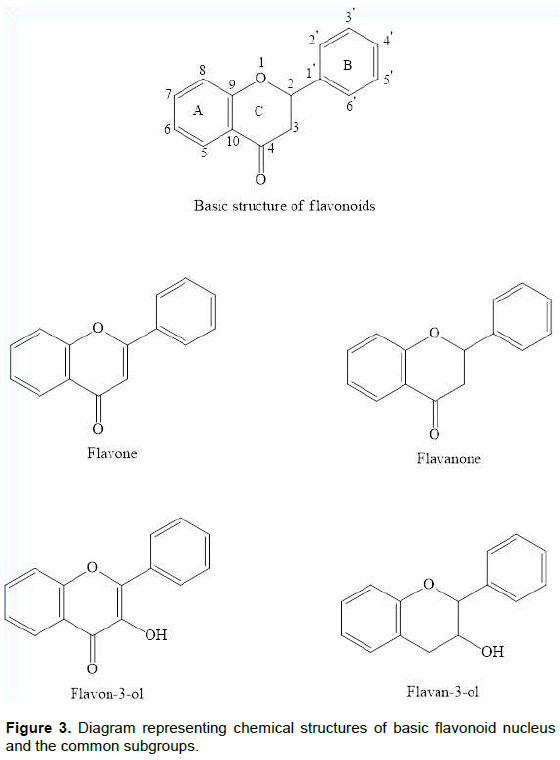
In plants, flavonoids not only provide UV-protection but also aid in the pollination by masking the flowers with attractive colors and patterns. Until now, more than 6000 varieties of flavonoids have been isolated. A variety of interest gasping pharmacological actions for these naturally occurring flavonoid compounds as well as synthetic derivatives have been found (Vidyalakshmi et al., 2010), including both peripheral and central nervous system effects (Figure 4) (Hall et al., 2005). These compounds attribute to a vast range of biological effects, like anticancer (Liu et al., 2010), anti-inflammatory (Wang et al., 2010), antioxidant (Heim et al., 2002), anxiolytic (Ognibene et al., 2008), cardio protective (Yu et al., 2005), antifungal (Ammar et al., 2013), antiviral (Orhan et al., 2010), antimicrobial (Cushnie and Lamb, 2005), neuroprotective (Cho et al., 2013)and antinociceptive activities (Wang et al., 2014).

FLAVONOIDS AS GABAA RECEPTOR MODULATORS
Flavonoids have been extensively focused for their peripheral events; however, more recently their selective affinity for GABAA receptors has been reported in studies using rat and bovine brain membrane binding assays (Hong and Hopfinger, 2003). In conjunction with binding studies, many behavioral studies have also been carried out, which indicate the anxiolytic effects of flavonoids in rodent anxiety models lacking many of the surplus side effects of BDZs (benzodiazepines) (Griebel et al., 1999). Interestingly, the positive, negative and neutralizing allosteric modulatory actions of flavonoids over a wide range of ionotropic GABA receptors has been focused and strongly supported via a large bulk of evidence.
The isolation of isoflavones from bovine urine that displaced 3[H]-diazepam binding in rat brain laid the foundation of the interaction of flavonoids with BDZ receptors (Luk et al., 1983). In 1990s, flavonoids were defined as a new family of BDZ receptor ligands (Medina et al., 1997; Marder and Paladini, 2002). Classically, they were considered to be acting upon BDZ receptors and a number of synthetic flavonoids having a significant affinity for BDZ binding site were developed accordingly (Yao et al., 2007); until they were reported to be insensitive to flumazenil that is BDZ receptor antagonist, thus highlighting a unique site of action for this class (Hanrahan et al., 2011).
The substitution at 6- or 3’-positions of flavones with an electronegative functional group enhanced the affinity towards the BDZ receptors (Paladini et al., 1999). Furthermore, the impact of ligand binding on the GABA binding was used to determine the GABA ratios. These ratios showed that flavones exhibited significant biological activities at BDZ receptors (Hanrahan et al., 2011). 6-Bromoflavone, 6-bromo-3’-nitroflavone and 6-chloro-3’-nitroflavone with a GABA ratio of 1.6-2.0, 1.38 and 2.0 were reported as full agonist (Marder et al., 1996), partial agonist (Wolfman et al., 1998)and an antagonist (Viola et al., 2000)at these receptors.
Positive ionotropic modulators of GABAA receptors enhance the chloride ion flux and provide a strong inhibitory effect. Therefore, for the management of CNS – allied diseases, including panic disorders, generalized anxiety, seizure disorders, muscle spasm, neuropathic pain and sleep disturbances; these modulators stand amongst the strongest candidates (Rudolph and Möhler, 2006). Moreover, with the discovery of the fact that flavonoids may act upon novel binding sites other than the classical benzodiazepine binding site, opportunities to search for new therapeutic agents with less adverse effects has been provided (Rudolph and Möhler, 2006). In this regard, 6-Methoxyflavonone has been reported to act as positive allosteric modulator at α1β2γ2L and α2β2γ2L subunits of GABAA receptors (Hall et al., 2014).
The substitution at 6-position on flavones is related to its effects on the recombinant GABAA receptors. 6-Hydroxyflavone depicted a significant effect at flumazenil-sensitive BDZ site (Ren et al., 2010). Moreover, 6-methoxyflavanone and 6-methoxyflavone have been recently reported to exhibit significant anti-allodynic effects in streptozotocin- and cisplatin-induced neuropathic pain models (Akbar et al., 2016; Shahid et al., 2017). These protective effects against neuropathic pain were attributed to the positive allosteric modulatory effects of these molecules on opioid and GABAA receptors respectively (Akbar et al., 2016).
Moreover, myrcitin and baicalin produced significant anti allodynic effects in sciatic nerve ligation models (Meotti et al., 2006; Cherng et al., 2014). Rutin and quercetin has been reported to inhibit oxaliplatin-induced chronic painful peripheral neuropathy (Azevedo et al., 2013). Naringin is also reported as exhibiting anti allodynic potential in the sreptozotocin-induced painful diabetic neuropathy (Kandhare et al., 2012).
OTHER NEUROPATHIC PAIN MODULATING MECHANISMS OF FLAVONOIDS
Besides its action on the GABAA receptors, flavonoids also exert antioxidant and anti-inflammatory effects. Almost all metabolic diseases are the consequence of oxidative stress. In addition to exogenous damage, the normal oxygen metabolism within the cells and tissues produce free radicals and reactive oxygen species that consistently endanger them. Flavonoids are very well known for their antioxidant potential and are proven to exert positive effects in diseases like atherosclerosis, diabetes, Alzheimer’s disease, cancer, etc (De Groot, 1994; Pal and Verma, 2013).
Although the underlying set of events behind the damage caused by the free radicals to the cellular functions is not fully understood, the contribution of lipid per oxidation leading to cellular membrane destruction and activation of inflammatory mediators by the free radicals culminating in ultimate tissue damage, can provide a lot of help in conceiving a pharmacological target. Nature has provided an inborn mechanism against these ROS, comprising of enzymes like superoxidase, glutathione peroxidase and catalase as well as non-enzymatic elements e.g. ascorbic acid, α-tochopherol, etc (Halliwell, 1995). However, these endogenous scavenging compounds are prone to depletion due to an increased oxidative stress caused by many diseased states including conditions that culminate in neuropathic pain e.g. diabetes mellitus (Schreiber et al., 2015).
Epicatechin and rutin are shown to have the ability to be oxidized themselves by the free radicals giving rise to a stable and less reactive species (Hanasaki et al., 1994). Similarly, quercetin inhibits the nitric oxide (NO) induced cell injury. Nitric oxide combines with the free radicals and produces an extremely harmful peroxynitrite that straightly oxidize LDL leading to a permanent destruction of the cell membrane. Hence, quercetin scavenge the free radicals, restraining them from reacting with NO (Shutenko et al., 1999), while silibin react directly with NO (Dehmlow et al., 1996).
Physiologically, the metabolism of xanthine to uric acid is carried out via xanthine dehydrogenase; however, in case of ischemic-reperfusion this enzyme changes into xanthine oxidase that acts as precursor of free radicals. Quercetin, luteolin and silibin flavonoids are known to act as antioxidant via inhibiting xanthine oxidase (Chang et al., 1992; Shoskes, 1998).
Similarly, reperfusion also results in the mobilization of leucocytes causing the consequent release of inflammatory mediators and cytotoxic oxidants provoking the complement system. A number of flavonoids are reportedly involved in the leucocytes immobilization ultimately causing a reduction in the serum complement system and inflammation (Friesenecker and Tsai, 1995; Ferrándiz et al., 1996). A pronounced body of evidence report the involvement of same pathophysiological mechanisms behind both inflammation and neuropathic pain of peripheral origin. Both types of pathologies manifest as allodynia and hyperalgesia (Clatworthy et al., 1995; DeLeo and Yezierski, 2001; Jin et al., 2003). Infiltration of inflammatory cells and their secretary products like arachidonic acid and cytokines (released for nerve regeneration) as a result of peripheral nerve injury is responsible for the generation and maintenance of the ongoing pain (Tracey and Walker, 1995; Cui et al., 2000; Ma and Eisenach, 2003). Cytokines like tumor necrosis factor-α (TNF-α), interleukin-1 (IL-1) and interleukin-6 (IL-6) when injected into the rat paw resulted in the induction of mechanical and thermal hyperalgesia (Cunha et al., 1992; Ferreira et al., 1993). The blockage of TNF–α in the rodent models of painful neuropathy has resulted in the attenuation of hyperalgesia (Sommer et al., 1998). Cytokines once released induce their own production and consequently activate COX–2 dependent prostanoid releases. The role of PGs in inducing inflammation accompanying enhanced sensitivity to pain is well documented. Intrathecal administration of PGs including PGE2 and PGF2α induced allodynia in conscious mice (Minami et al., 1992, 1994), whereas intrathecal injection of PGD2 and PGE2 resulted in the induction of hyperalgesia (Uda et al., 1990). Moreover, synthesis of PG and nitric oxide (NO) via COX-2 and inducible nitric oxide synthase (iNOS) is enhanced in the microglia as a result of peripheral nerve injury leading to hyper sensitization (Hanisch, 2002). Flavonoids exhibit an in vivo and in vitro anti-inflammatory activity. The in vivo anti-inflammatory is attributed to the inhibition of ecosanoid generating enzymes including COX, LOX and phospholipase A2 (Kim et al., 2004).
Flavonoids are known to act upon different enzyme systems, e.g. causes the inhibition of arachidonic acid and blocking the inflammatory response as arachidonic acid tends to start it. By this, antithrombotic and anti-inflammatory features are incorporated into the group (Ferrandiz and Alcaraz, 1991). They also cause decrease in the release of peroxidase and inhibition of ROS production by neutrophils. This inhibition is carried out via interference with α1-antitrypsin activation (Middleton and Kandaswami, 1992). Certain flavonoids are involved in iron chelation that causes lipid peroxidation, thereby abolishing an erratic factor for the free radicals development (Nelson et al., 1992; Ferrali et al., 1997).
CONCLUSION
Allosteric modulators at GABAA receptors modify either the efficacy or affinity of agonists like GABA, consequently regulating their activity. Over the last decade, these modulators have been focused extensively due to the advances in the understanding of the functions of GABAA receptor subtypes. Flavonoids being potent allosteric modulators may prove to be useful tools in the amelioration of such distressing painful conditions like neuropathic pain. However more studies are required to further elucidate the site of action of these bioactive molecules over the GABAA receptors.
CONFLICT OF INTERESTS
The author has not declared any conflict of interests.
REFERENCES
|
Aguirre-Hernández E, González-Trujano ME, Martínez AL, Moreno J, Kite G, Terrazas T, Soto-Hernández M (2010). HPLC/MS analysis and anxiolytic-like effect of quercetin and kaempferol flavonoids from Tilia americana var. mexicana. Journal of Ethnopharmacology 127(1): 91-97. |
|
|
Akbar S, Subhan F, Karim N, Shahid M, Ahmad N, Ali G, Mahmood W, Fawad K (2016). 6-Methoxyflavanone attenuates mechanical allodynia and vulvodynia in the streptozotocin-induced diabetic neuropathic pain. Biomedicine and Pharmacotherapy 84:962-971. |
|
|
Ammar MI, Nenaah GE, Mohamed AHH (2013).Antifungal activity of prenylated flavonoids isolated from Tephrosia apollinea L. against four phytopathogenic fungi. Crop Protection 49:21-25. |
|
|
Ataka T, Kumamoto E, Shimoji K, Yoshimura M (2000). Baclofen inhibits more effectively C-afferent than Aδ-afferent glutamatergic transmission in substantia gelatinosa neurons of adult rat spinal cord slices. Pain 86(3):273-282. |
|
|
Attal N (2001). Pharmacologic treatment of neuropathic pain. Acta Neurologica Belgica 101(1):53-64. |
|
|
Azevedo MI, Pereira AF, Nogueira RB, Rolim F E, Brito GA, Wong DVT, Lima-Júnior RC, de Albuquerque Ribeiro R, Vale ML (2013). The antioxidant effects of the flavonoids rutin and quercetin inhibit oxaliplatin-induced chronic painful peripheral neuropathy. Molecular Pain 9(1):53. |
|
|
Baron R (2006). Mechanisms of disease: neuropathic pain-a clinical perspective. Nature Clinical Practice Neurology 2(2):95-106. |
|
|
Baron R, Levine JD, Fields HL (1999). Causalgia and reflex sympathetic dystrophy: does the sympathetic nervous system contribute to the generation of pain? Muscle and nerve 22(6):678-695. |
|
|
Bráz JM, Sharif-Naeini R, Vogt D, Kriegstein A, Alvarez-Buylla A, Rubenstein JL, Basbaum AI (2012). Forebrain GABAergic neuron precursors integrate into adult spinal cord and reduce injury-induced neuropathic pain. Neuron 74(4):663-675. |
|
|
Campbell JN, Meyer RA (2006). Mechanisms of neuropathic pain. Neuron 52(1):77-92. |
|
|
Chang WS, Lee YJ, Lu F, Chiang HC (1992). Inhibitory effects of flavonoids on xanthine oxidase. Anticancer Research 13(6A):2165-2170. |
|
|
Cheng HT, Suzuki M, Hegarty DM, Xu Q, Weyerbacher AR, South SM, Ohata M, Inturrisi CE (2008). Inflammatory pain-induced signaling events following a conditional deletion of the N-methyl-D-aspartate receptor in spinal cord dorsal horn. Neuroscience 155(3):948-958. |
|
|
Cherng CH, Lee KC, Chien CC, Chou KY, Cheng YC, Hsin ST, Lee SO, Shen CH, RY Tsai, CS Wong (2014). Baicalin ameliorates neuropathic pain by suppressing HDAC1 expression in the spinal cord of spinal nerve ligation rats. Journal of the Formosan Medica l Association 113(8):513-520. |
|
|
Cho N, Lee KY, Huh J, Choi JH, Yang H, Jeong EJ, Kim HP, Sung SH (2013). Cognitive-enhancing effects of Rhus verniciflua bark extract and its active flavonoids with neuroprotective and anti-inflammatory activities. Food and chemical toxicology 58:355-361. |
|
|
Clatworthy A, Illich P, Castro G, Walters E (1995). Role of peri-axonal inflammation in the development of thermal hyperalgesia and guarding behavior in a rat model of neuropathic pain. Neuroscience letters 184(1):5-8. |
|
|
Cui JG, Holmin S, Mathiesen T, Meyerson BA, Linderoth B (2000). Possible role of inflammatory mediators in tactile hypersensitivity in rat models of mononeuropathy. Pain 88(3):239-248. |
|
|
Cunha F, Poole S, Lorenzetti B, Ferreira S (1992). The pivotal role of tumour necrosis factor α in the development of inflammatory hyperalgesia. British journal of pharmacology 107(3):660-664. |
|
|
Cushnie T, Lamb AJ (2005). Antimicrobial activity of flavonoids. International journal of antimicrobial agents 26(5):343-356. |
|
|
De Groot H (1994). Reactive oxygen species in tissue injury. Hepato-gastroenterology 41(4):328-332. |
|
|
Dehmlow C, Erhard J, de Groot H (1996). Inhibition of Kupffer cell functions as an explanation for the hepatoprotective properties of silibinin. Hepatology 23(4):749-754. |
|
|
De Leo JA, Yezierski RP (2001). The role of neuroinflammation and neuroimmune activation in persistent pain. Pain 90(1-2):1-6. |
|
|
Eaton MJ, Plunkett JA, Martinez MA, Lopez T, Karmally S, Cejas P, Whittemore SR (1998). Transplants of neuronal cells bioengineered to synthesize GABA alleviate chronic neuropathic pain. Cell transplantation 8(1):87-101. |
|
|
Eaton MJ, Martinez MA, Karmally S (1999). A single intrathecal injection of GABA permanently reverses neuropathic pain after nerve injury. Brain research 835(2):334-339. |
|
|
Ferrali M, Signorini C, Caciotti B, Sugherini L, Ciccoli L, Giachetti D, Comporti M (1997). Protection against oxidative damage of erythrocyte membrane by the flavonoid quercetin and its relation to iron chelating activity. FEBS letters 416(2):123-129. |
|
|
Ferrandiz M, Alcaraz M (1991). Anti-inflammatory activity and inhibition of arachidonic acid metabolism by flavonoids. Agents and actions 32(3-4):283-288. |
|
|
Ferrandiz ML, Gil B, Sanz MJ, Ubeda A, Erazo S, Gonzalez E, Negrete R, Pacheco S, Payaa M, Alcaraz MJ (1996). Effect of bakuchiol on leukocyte functions and some inflammatory responses in mice. Journal of pharmacy and pharmacology 48(9):975-980. |
|
|
Ferreira S, Lorenzetti B, Poole S (1993). Bradykinin initiates cytokineâ€mediated inflammatory hyperalgesia. British journal of pharmacology 110(3):1227-1231. |
|
|
Formica J, Regelson W (1995). Review of the biology of quercetin and related bioflavonoids. Food and chemical toxicology 33(12):1061-1080. |
|
|
Friesenecker B, Tsai A (1995). Cellular basis of inflammation, edema and the activity of Daflon 500 mg. International Journal of Microcirculation 15(Suppl. 1):17-21. |
|
|
Gavande N, Karim N, Johnston GA, Hanrahan JR, Chebib M (2011). Identification of Benzopyranâ€4â€one Derivatives (Isoflavones) as Positive Modulators of GABAA Receptors. ChemMedChem 6(8):1340-1346. |
|
|
Griebel G, Perrault G, Tan S, Schoemaker H, Sanger DJ (1999). Pharmacological studies on synthetic flavonoids: comparison with diazepam. Neuropharmacology 38(7):965-977. |
|
|
Groot Hd, Rauen U (1998). Tissue injury by reactive oxygen species and the protective effects of flavonoids. Fundamental & clinical pharmacology 12(3):249-255. |
|
|
Gwak YS, Tan HY, Nam TS, Paik KS, Hulsebosch CE, Leem JW (2006). Activation of spinal GABA receptors attenuates chronic central neuropathic pain after spinal cord injury. Journal of neurotrauma 23(7):1111-1124. |
|
|
Hains BC, Saab CY, Klein JP, Craner MJ, Waxman SG (2004). Altered sodium channel expression in second-order spinal sensory neurons contributes to pain after peripheral nerve injury. The Journal of neuroscience 24(20):4832-4839. |
|
|
Hall BJ, Chebib M, Hanrahan JR, Johnston GAR (2005). 6-Methylflavanone, a more efficacious positive allosteric modulator of γ-aminobutyric acid (GABA) action at human recombinant α2β2γ2L than at α1β2γ2L and α1β2 GABAA receptors expressed in Xenopus oocytes. European Journal of Pharmacology 512(2-3):97-104. |
|
|
Hall BJ, Karim N, Chebib M, Johnston GA, Hanrahan JR (2014). Modulation of ionotropic GABA receptors by 6-methoxyflavanone and 6-methoxyflavone. Neurochemical research 39(6):1068-1078. |
|
|
Halliwell B (1995). How to characterize an antioxidant: an update. In Biochemical Society Symposia. Portland Press Limited. 61:73-101. |
|
|
Hanasaki Y, Ogawa S, Fukui S (1994). The correlation between active oxygens scavenging and antioxidative effects of flavonoids. Free Radical Biology and Medicine 16(6):845-850. |
|
|
Hanisch UK (2002). Microglia as a source and target of cytokines. Glia 40(2):140-155. |
|
|
Hanrahan JR, Chebib M, Johnston GA (2011). Flavonoid modulation of GABAA receptors. British journal of pharmacology 163(2):234-245. |
|
|
Harden RN (1999). Gabapentin: a new tool in the treatment of neuropathic pain. Acta Neurologica Scandinavica 100:43-47. |
|
|
Heim KE, Tagliaferro AR, Bobilya DJ (2002). Flavonoid antioxidants: chemistry, metabolism and structure-activity relationships. The Journal of Nutritional Biochemistry 13(10):572-584. |
|
|
Hendrich J, Van Minh AT, Heblich F, Nieto-Rostro M, Watschinger K, Striessnig J, Wratten J, Davies A, Dolphin AC (2008). Pharmacological disruption of calcium channel trafficking by the α2δ ligand gabapentin. Proceedings of the National Academy of Sciences 105(9):3628-3633. |
|
|
Hong X, Hopfinger AJ (2003). 3D-pharmacophores of flavonoid binding at the benzodiazepine GABAA receptor site using 4D-QSAR analysis. Journal of chemical information and computer sciences 43(1):324-336. |
|
|
Hwang JH, Yaksh TL (1997). The effect of spinal GABA receptor agonists on tactile allodynia in a surgically-induced neuropathic pain model in the rat. Pain 70(1):15-22. |
|
|
Jäger AK, Krydsfeldt K, Rasmussen HB (2009). Bioassayâ€guided isolation of apigenin with GABAâ€benzodiazepine activity from Tanacetum parthenium. Phytotherapy Research 23(11):1642-1644. |
|
|
Jager AK, Saaby L (2011). Flavonoids and the CNS. Molecules 16(2):1471-1485. |
|
|
Jensen TS (2006). Translation of symptoms and signs into mechanisms in neuropathic pain. Frontiers in Headache Research 14:51. |
|
|
Jergova S, Hentall ID, Gajavelli S, Varghese MS, Sagen J (2012). Intraspinal transplantation of GABAergic neural progenitors attenuates neuropathic pain in rats: a pharmacologic and neurophysiological evaluation. Experimental neurology 234(1):39-49. |
|
|
Ji RR, Woolf CJ (2001). Neuronal plasticity and signal transduction in nociceptive neurons: implications for the initiation and maintenance of pathological pain. Neurobiology of disease 8(1):1-10. |
|
|
Jin SX, Zhuang ZY, Woolf CJ, Ji RR (2003). p38 mitogen-activated protein kinase is activated after a spinal nerve ligation in spinal cord microglia and dorsal root ganglion neurons and contributes to the generation of neuropathic pain. The Journal of neuroscience 23(10):4017-4022. |
|
|
Kandhare AD, Raygude KS, Ghosh P, Ghule AE, Bodhankar SL (2012). Neuroprotective effect of naringin by modulation of endogenous biomarkers in streptozotocin induced painful diabetic neuropathy. Fitoterapia 83(4):650-659. |
|
|
Kim HP, Son KH, Chang HW, Kang SS (2004). Anti-inflammatory plant flavonoids and cellular action mechanisms. Journal of pharmacological sciences 96(3):229-245. |
|
|
Knabl J, Witschi R, Hosl K, Reinold H, Zeilhofer UB, Ahmadi S, Brockhaus J, Sergejeva M, Hess A, Brune K (2008). Reversal of pathological pain through specific spinal GABAA receptor subtypes. Nature 451(7176):330-334. |
|
|
Lai J, Hunter JC, Porreca F (2003). The role of voltage-gated sodium channels in neuropathic pain. Current opinion in neurobiology 13(3):291-297. |
|
|
Lindenlaub T, Sommer C (2003). Cytokines in sural nerve biopsies from inflammatory and non-inflammatory neuropathies. Acta neuropathologica 105(6):593-602. |
|
|
Liu HL, WB Jiang, Xie MX (2010). Flavonoids: recent advances as anticancer drugs. Recent patents on anti-cancer drug discovery 5(2):152-164. |
|
|
Luk KC, Stern L, Weigele M, O'Brien RA, Spirt N (1983). Isolation and identification of" diazepam-like" compounds from bovine urine. Journal of natural products 46(6):852-861. |
|
|
Ma W, Eisenach J (2003). Cyclooxygenase 2 in infiltrating inflammatory cells in injured nerve is universally up-regulated following various types of peripheral nerve injury. Neuroscience 121(3):691-704. |
|
|
Malan TP, Mata HP, Porreca F (2002). Spinal GABA (A) and GABA (B) receptor pharmacology in a rat model of neuropathic pain. Anesthesiology 96(5):1161-1167. |
|
|
Marder M, Paladini AC (2002). GABA(A)-receptor ligands of flavonoid structure. Current topics in medicinal chemistry 2(8):853-867. |
|
|
Marder M, Viola H, Wasowski C, Wolfman C, Waterman PG, Cassels BK, Medina JH, Paladini AC (1996). 6-Bromoflavone, a high affinity ligand for the central benzodiazepine receptors is a member of a family of active flavonoids. Biochemical and biophysical research communications 223(2):384-389. |
|
|
McCarson KE, Enna S (2014). GABA pharmacology: The search for analgesics. Neurochemical Research 39(10):1948-1963. |
|
|
McKemy DD, Neuhausser WM, Julius D (2002). Identification of a cold receptor reveals a general role for TRP channels in thermosensation. Nature 416(6876):52-58. |
|
|
Medina JH, Viola H, Wolfman C, Marder M, Wasowski C, Calvo D, Paladini AC (1997). Overview-flavonoids: a new family of benzodiazepine receptor ligands. Neurochemical research 22(4):419-425. |
|
|
Melzack R, Wall PD (1967). Pain mechanisms: a new theory. Survey of Anesthesiology 11(2):89-90. |
|
|
Meotti FC, Missau FC, Ferreira J, Pizzolatti MG, Mizuzaki C, Nogueira CW, Santos AR (2006). Anti-allodynic property of flavonoid myricitrin in models of persistent inflammatory and neuropathic pain in mice. Biochemical Pharmacology 72(12):1707-1713. |
|
|
Middleton E, Kandaswami C (1992). Effects of flavonoids on immune and inflammatory cell functions. Biochemical Pharmacology 43(6):1167-1179. |
|
|
Middleton Jr E (1998). Effect of plant flavonoids on immune and inflammatory cell function. Flavonoids in the Living System. Springer pp. 175-182. |
|
|
Minami T, Uda R, Horiguchi S, Ito S, Hyodo M, Hayaishi O (1992). Allodynia evoked by intrathecal administration of prostaglandin F 2α to conscious mice. Pain 50(2):223-229. |
|
|
Minami T, Uda R, Horiguchi S, Ito S, Hyodo M, Hayaishi O (1994). Allodynia evoked by intrathecal administration of prostaglandin E2 to conscious mice. Pain 57(2):217-223. |
|
|
Moore KA, Kohno T, Karchewski LA, Scholz J, Baba H, Woolf CJ (2002). Partial peripheral nerve injury promotes a selective loss of GABAergic inhibition in the superficial dorsal horn of the spinal cord. The Journal of neuroscience 22(15):6724-6731. |
|
|
Nelson CW, Wei EP, Povlishock JT, Kontos HA, Moskowitz MA (1992). Oxygen radicals in cerebral ischemia. American Journal of Physiology-Heart and Circulatory Physiology 263(5):H1356-H1362. |
|
|
Ognibene E, Bovicelli P, Adriani W, Saso L, Laviola G (2008). Behavioral effects of 6-bromoflavanone and 5-methoxy-6, 8-dibromoflavanone as anxiolytic compounds. Progress in Neuro-Psychopharmacology and Biological Psychiatry 32(1):128-134. |
|
|
Orhan DD, Ozcelik B, Ozgen S, Ergun F (2010). Antibacterial, antifungal, and antiviral activities of some flavonoids. Microbiological research 165(6):496-504. |
|
|
Ossipov MH, Dussor GO, Porreca F (2010). Central modulation of pain. The Journal of clinical investigation 120(11):3779-3787. |
|
|
Pal D, Verma P (2013). Flavonoids: A powerful and abundant source of antioxidants. International Journal of Pharmacy and Pharmaceutical Sciences 5(3):95-98. |
|
|
Paladini A, Marder M, Viola H, Wolfman C, Wasowski C, Medina J (1999). Flavonoids and the central nervous system: from forgotten factors to potent anxiolytic compounds. Journal of Pharmacy and Pharmacology 51(5):519-526. |
|
|
Patapoutian A, Peier AM, Story GM, Viswanath V (2003). ThermoTRP channels and beyond: mechanisms of temperature sensation. Nature Reviews Neuroscience 4(7):529-539. |
|
|
Persohn E, Malherbe P, Richards J (1991). In situ hybridization histochemistry reveals a diversity of GABA A receptor subunitmRNAs in neurons of the rat spinal cord and dorsal root ganglia. Neuroscience 42(2):497-507. |
|
|
Peterson J, Dwyer J (1998). Flavonoids: dietary occurrence and biochemical activity. Nutrition Research 18(12):1995-2018. |
|
|
Price G, Wilkin G, Turnbull M, Bowery N (1984). Are baclofen-sensitive GABAB receptors present on primary afferent terminals of the spinal cord? Nature 304(5946):71. |
|
|
Price MP, McIlwrath SL, Xie J, Cheng C, Qiao J, Tarr DE, Sluka KA, Brennan TJ, Lewin GR, Welsh MJ (2001). The DRASIC cationchannel contributes to the detection of cutaneous touch and acid stimuli in mice. Neuron 32(6):1071-1083. |
|
|
Rasmussen PV, Sindrup SH, Jensen TS, Bach FW (2004). Symptoms and signs in patients with suspected neuropathic pain. Pain 110(1):461-469. |
|
|
Reeve AJ, Dickenson AH, Kerr NC (1998). Spinal effects of bicuculline: modulation of an allodynia-like state by an A1-receptor agonist, morphine, and an NMDA-receptor antagonist. Journal of neurophysiology 79(3):1494-1507. |
|
|
Ren L, Wang F, Xu Z, Chan WM, Zhao C, Xue H (2010). GABAA receptor subtype selectivity underlying anxiolytic effect of 6-hydroxyflavone. Biochemical Pharmacology 79(9):1337-1344. |
|
|
Riley R, Trafton J, Chi S, Basbaum A (2001). Presynaptic regulation of spinal cord tachykinin signaling via GABA B but not GABA A receptor activation. Neuroscience 103(3):725-737. |
|
|
Rudolph U, Mohler H (2006). GABA-based therapeutic approaches: GABAA receptor subtype functions. Current Opinion in Pharmacology 6(1):18-23. |
|
|
Schreiber AK, Nones CF, Reis RC, Chichorro JG, Cunha JM (2015). Diabetic neuropathic pain: physiopathology and treatment. World journal of diabetes 6(3):432. |
|
|
Semyanov A, Walker MC, Kullmann DM (2003). GABA uptake regulates cortical excitability via cell type-specific tonic inhibition. Nature neuroscience 6(5):484-490. |
|
|
Shahid M, Subhan F, Ahmad N, Sewell RD (2017). The flavonoid 6-methoxyflavone allays cisplatin-induced neuropathic allodynia and hypoalgesia. Biomedicine and Pharmacotherapy 95:1725-1733. |
|
|
Shoskes DA (1998). Effect of Bioflavonoids Quercetin and Curcumin on Ischemic Renal Injury: A New Class of Renoprotective Agents1. Transplantation 66(2):147-152. |
|
|
Shutenko Z, Henry Y, Pinard E, Seylaz J, Potier P, Berthet F, Girard P, Sercombe R (1999). Influence of the antioxidant quercetin in vivo on the level of nitric oxide determined by electron paramagnetic resonance in rat brain during global ischemia and reperfusion. Biochemical Pharmacology 57(2):199-208. |
|
|
Sivilotti L, Woolf CJ (1994). The contribution of GABAA and glycine receptors to central sensitization: disinhibition and touch-evoked allodynia in the spinal cord. Journal of Neurophysiology 72(1):169-179. |
|
|
Sommer C (2003). Painful neuropathies. Current Opinion in Neurology 16(5):623-628. |
|
|
Sommer C, Schmidt C, George A (1998). Hyperalgesia in experimental neuropathy is dependent on the TNF receptor 1. Experimental neurology 151(1):138-142. |
|
|
Stubley LA, Martinez MA, Karmally S, Lopez T, Cejas P, Eaton MJ (2001). Only early intervention with gamma-aminobutyric acid cell therapy is able to reverse neuropathic pain after partial nerve injury. Journal of neurotrauma 18(4):471-477. |
|
|
Tal M, Bennett GJ (1994). Extra-territorial pain in rats with a peripheral mononeuropathy: mechano-hyperalgesia and mechano-allodynia in the territory of an uninjured nerve. Pain 57(3):375-382. |
|
|
Tracey D, Walker J (1995). Pain due to nerve damage: are inflammatory mediators involved. Inflammation Research 44(10):407-411. |
|
|
Treede R, Jensen T, Campbell J, Cruccu G, Dostrovsky J, Griffin J, Hansson P, Hughes R, Nurmikko T, Serra J (2008). Redefinition of neuropathic pain and a grading system for clinical use: consensus statement on clinical and research diagnosis criteria. Neurology 70:1360-1365. |
|
|
Tucker AP, Mezzatesta J, Nadeson R, Goodchild CS (2004). Intrathecal midazolam II: combination with intrathecal fentanyl for labor pain. Anesthesia & Analgesia 98(6):1521-1527. |
|
|
Turner MS (2003). Intrathecal drug delivery 2002. Neurosurgical Re-Engineering of the Damaged Brain and Spinal Cord. Springer pp. 29-35. |
|
|
Tyson JA, Anderson SA (2014). GABAergic interneuron transplants to study development and treat disease. Trends in Neurosciences 37(3):169-177. |
|
|
Uda R, Horiguchi S, Ito S, Hyodo M, Hayaishi O (1990). Nociceptive effects induced by intrathecal administration of prostaglandin D2, E2, or F2α to conscious mice. Brain research 510(1):26-32. |
|
|
Ultenius C, Linderoth B, Meyerson BA, Wallin J (2006). Spinal NMDA receptor phosphorylation correlates with the presence of neuropathic signs following peripheral nerve injury in the rat. Neuroscience letters 399(1):85-90. |
|
|
Uphoff J, Binder A (2006). No adrenergic sensitization of afferent neurons in painful sensory polyneuropathy. Journal of Neurology 253(3):280-286. |
|
|
Vidyalakshmi K, Kamalakannan P, Viswanathan S, Ramaswamy S (2010). Antinociceptive effect of certain dihydroxy flavones in mice. Pharmacology Biochemistry and Behavior 96(1):1-6. |
|
|
Viola H, Wolfman C, Marder M, Goutman J, Bianchin M, Wasowski C, Calvo D, Izquierdo I, Paladini A, Medina J (2000). 6-Chloro-3′-nitroflavone is a potent ligand for the benzodiazepine binding site of the GABAA receptor devoid of intrinsic activity. Pharmacology Biochemistry and Behavior 65(2):313-320. |
|
|
Wafford KA. (2005). GABA A receptor subtypes: any clues to the mechanism of benzodiazepine dependence? Current Opinion in Pharmacology 5(1):47-52. |
|
|
Wang D, Tang W, Yang GM, Cai BC (2010). Anti-inflammatory, Antioxidant and Cytotoxic Activities of Flavonoids from Oxytropis falcata Bunge. Chinese Journal of Natural Medicines 8(6):461-465. |
|
|
Wang Y, Chen P, Tang C, Wang Y, Li Y, Zhang H (2014). Antinociceptive and anti-inflammatory activities of extract and two isolated flavonoids of Carthamus tinctorius L. Journal of Ethnopharmacology 151(2):944-950. |
|
|
Wolfman C, Viola H, Marder M, Ardenghi P, Wasowski C, Schroeder N, Izquierdo I, Ruveda E, Paladini A, Medina J (1998). Pharmacological characterization of 6-bromo-3′-nitroflavone, a synthetic flavonoid with high affinity for the benzodiazepine receptors. Pharmacology Biochemistry and Behavior 61(3):239-246. |
|
|
Wasner G, Schattschneider J, Binder A, Baron R (2004). Topical menthol-a human model for cold pain by activation and sensitization of C nociceptors. Brain 127(5):1159-1171. |
|
|
Woolf CJ (2011). Central sensitization: implications for the diagnosis and treatment of pain. Pain 152(3):S2-S15. |
|
|
Yaksh TL (1989). Behavioral and autonomic correlates of the tactile evoked allodynia produced by spinal glycine inhibition: effects of modulatory receptor systems and excitatory amino acid antagonists. Pain 37(1):111-123. |
|
|
Yamamoto T, Yaksh TL (1993). Effects of intrathecal strychnine and bicuculline on nerve compression-induced thermal hyperalgesia and selective antagonism by MK-801. Pain 54(1):79-84. |
|
|
Yao N, Song A, Wang X, Dixon S, Lam KS (2007). Synthesis of flavonoid analogues as scaffolds for natural product-based combinatorial libraries. Journal of Combinatorial Chemistry 9(4):668-676. |
|
|
Yu LL, Zhou KK, Parry J (2005). Antioxidant properties of cold-pressed black caraway, carrot, cranberry, and hemp seed oils. Food Chemistry 91(4):723-729. |
|
|
Zeilhofer HU (2008). Loss of glycinergic and GABAergic inhibition in chronic pain-contributions of inflammation and microglia. International immunopharmacology 8(2):182-187. |
|
|
Zeilhofer HU, Mohler H, Di Lio A (2009). GABAergic analgesia: new insights from mutant mice and subtype-selective agonists. Trends in Pharmacological Sciences 30(8):397-402. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


