Full Length Research Paper

ABSTRACT
Annona senegalensis Pers. is a plant used in Burkina folkloric medicine for the treatment of epilepsy. The stem-bark, roots and leaves of A. senegalensis are used for an array of human disorders, including epilepsy. Previous studies have demonstrated some genuine effects of A. senegalensis fractions in vivo models of temporal lobe epilepsy. In the present work, we have compared the pharmacological profile of the most promising A. senegalensis fraction obtained up-to-date with anticonvulsant drugs targeting voltage-activated channels namely Carbamazepine (Na+ channels) and Retigabine (KCNQ Channels). In an in vitro model of temporal lobe epilepsy, we could observe a dose-dependent reduction of electrically evoked epileptiform bursts in the CA1 region of the rat hippocampus when applying carbamazepine, retigabine or the F4 fraction of A. senegalensis. The intrinsic excitability of CA1 pyramidal neurons was dose-dependently reduced by A. senegalensis. Since A. senegalensis effects could not anymore be observed when endogenous voltage-dependent sodium channels were blocked and intrinsic excitability restored with a dynamic clamp amplifier, it was concluded that A. senegalensis contains molecules that target voltage-dependent sodium channels.
Key words: Hippocampus, epilepsy, Annona senegalensis, Burkina Faso plant extracts, sodium channel, KCNQ channel, M-current, retigabine, carbamazepine.
Abbreviation: PTX, Picrotoxin; Vm, membrane potential.INTRODUCTION
Epilepsy is a disorder of the brain defined by at least two unprovoked seizures occurring more than 24 h apart (Fisher et al., 2014). It is characterised by episodes of excessive synchronised neuronal activity (Stafstrom and Carmant, 2015). Seizures result from sustained firing of Na+ dependent action potentials riding on a slow depolarised potential, mainly generated by ionotropic glutamate receptor currents (McCormick and Contreras, 2001). Disease management can require costly treatments (Bruno et al., 2013, Privitera, 2011; Yemadje et al., 2011). In addition, the lack of social coverage and qualified personnel makes the situation of epilepsy difficult. Finally, most of the currently used antiepileptic drugs have significant safety and efficacy deficits (Britton and So, 1995; Malagon-Valdez, 2004; Perucca and Mula, 2013). Indeed, these available drugs are associated with drug interaction making it difficult to attain seizure control (Yemitan and Adeyemi, 2013; Perucca and Mula, 2013). Therefore, development of new and reliable therapeutic treatments with better safety, efficacy profile and low cost is necessary. Traditional African medicine has been used for many years for the prevention and treatment of various diseases with few adverse effects. One of such medicinal plants used in the traditional management of epilepsy as the most popular plant for the treatment of various disorders is Annona senegalensis Pers. It has also been successfully used against febrile convulsions by the local people thereby giving this shrub prominence in ethnomedicine or folk medicine. The therapeutic strategy in countering epilepsy involves reducing neuronal excitability through different mechanistic pathways. Most therapeutics currently used in the treatment of epilepsy is either directed towards blocking voltage-gated sodium and calcium channels or potentiating GABA mediated neurotransmission. More recently a potassium channel (KCNQ) opener, retigabine, has successfully been approved as an alternative target and thus expanded the spectrum of ion channels targets by Anti-Epileptic Drugs (AEDs) (Rostock et al., 1996). Many studies have been reported the anticonvulsant effects of roots barks and fractions of this plant (Konate et al., 2012a, b; Okoli et al., 2010).
In our previous studies, one fraction (F4) was obtained from roots bark of this plant, and identified significant anticonvulsant effects in in vivo models of temporal lobe epilepsy (Konate et al., 2012a, b). Therefore, in the present study, we investigated the possible mechanism of action of this fraction (F4), offering a scientific basis of the traditional use of this plant for treatment of epilepsy.
MATERIALS AND METHODS
Plant material
Root barks of A. senegalensis Pers used in this study were harvested in the rain season (August, 2010) from Ouagadougou (District of Kadiogo, Burkina Faso). The plant was identified at the National Herbarium of Bukina under the reference number HNBU 8713.
Reagent setup
Experimental animals, anesthetics, slice handling, dissection dish, and other tools for dissection and vibratome, like:
Artificial cerebrospinal fluid (ACSF)
125 mM NaCl, 2.5 mM KCl, 0.8 mM NaH2PO4, 26 mM NaHCO3, 2 mM CaCl2, 1.3 mM MgCl2, and 10 mM D-glucose, pH 7.4., bubbled with 95% O2-5% CO2.
Hippocampal slice cultures solution
The culture medium contained (in ml) 25 MEM, 12.5 HBSS, 12.5 horse serum, 0.5 penicillin/streptomycin, 0.8 glucose (1 M), 0.1 ascorbic acid (1 mg/ml), 0.4 HEPES (1 M), 0.5 B27, and 8.95 sterile H2O.
Microelectrode solution
For whole-cell recordings: 140 KCl, 5 HEPES, 2 EGTA, 2 Na2ATP, 0.3 NaGTP, and 2 MgCl2, pH 7.4 or with a low chloride solution containing (in mM): 140 Kgluconate, 10 HEPES, 2 EGTA, 2 Na2ATP, 0.3 NaGTP, and 2 MgCl2, pH 7.4
Equipment setup
Platinum wire U-piece with nylon threads, inverted suction pipette, syringes, microscope with fiber optic light source, computer with monitor and software, patch clamp amplifier, vibration isolation stage microelectrodes, microelectrode holder, micromanipulator, recording chamber, and drug application system.
Electrophysiological recordings were performed in whole-cell configuration with a Multiclamp 700B amplifier (Axon Instruments), filtered at 5 kHz and digitized at 20 kHz via a PCI-6220 National Instrument interface controlled by IgorPro (Wavemetrics) and/or Digidata 1322A interface controlled by PClamp software (Axon Instruments).
Sample preparation
Root barks of A. senegalensis Pers were cleaned, shade-dried, crushed and sieved. The powder (100.0 g) was mixed in 500.0 ml of methanol in a beaker before being introduced into a glass column (2 L). After 24 h of maceration, a leaching was done. The methanolic extract collected was concentrated under reduced pressure using a rotary evaporator. The first part of the concentrated extract (F1) was stored and the second part was used for liquid-liquid fractionation with n-hexane. The n-hexane fraction (F2) was subsequently concentrated under rota vapor. A concentrated part of the obtained extract (F2) was kept and the second part was used for liquid-liquid separation with ethyl acetate. After this further separation, two fractions were collected: the ethyl acetate fraction (F3) and the residual aqueous fraction. The ethyl acetate fraction (F3) was evaporated to dryness and the residual aqueous fraction (F4) was lyophilized and then stored in a desiccator for phytochemical and pharmacological analyses. The extractions were repeated two to three times and the extracts are then combined.
Animals
Male Wistar rats (postnatal day 14-20) provided from Institut National de la Santé et de la Recherche Médicale (INSERM UMR_S 1072) (Marseille/France) were used in the present study. The rats were kept under conventional conditions on a 12:12 h light/dark cycle and had ad libitum access to feed pellets and water. The experimental protocols were approved by the local direction of veterinary services (Direction Départementale des Services Vétérinaires – Préfecture des Bouches du Rhône, France (O. Caillard, licence number A13-493). All experiments were carried out according to the European and institutional guidelines for the care and use of laboratory animals (Council Directive 86/609/EEC and French National Research Council).
Hippocampal slice preparation
Transverse hippocampal slices were prepared according to Debanne et al. (2008) described method. Briefly, rats (postnatal day 7-8) were decapitated under chloral hydrate anesthesia and their hippocampi were dissected, and immersed into cutting solution (ACSF). Transverse hippocampal slices 350 µm in thickness were prepared using a Leica VT1000S micro-tome (Leica, Nussloch, Germany). Olympus BX51WI microscope equipped with differential interference contrast optics under infrared illumination and a water immersion lens (×60, 0.9 NA; Olympus) were used to visualize CA1 pyramidal cells.
Electrophysiological recordings
Recordings of the electrophysiological in whole-cell configuration were made using a Multiclamp 700B amplifier (Axon Instruments), filtered at 5 kHz and digitized at 20 kHz via a PCI-6220 National Instrument interface controlled by IgorPro (Wavemetrics) and/or Digidata 1322A interface controlled by PClamp software (Axon Instruments) for the electrophysiological. The reversal potential for GABAA receptors, determined by local pressure ejection of isoguvacine (0.5 mM) at different holding membrane potentials, was 3.7 ± 2.9 mV (n=4). With the low chloride-based solution, the reversal potential for GABAA receptors, determined by the same method was ‑73.7±1.7 mV (n=9). Cells were recorded if the series resistance, measured throughout experiments, remained stable and <40 MΩ. Stimulating pipettes were filled with extracellular saline. Picrotoxin (PTX, 100 mM) was prepared in ethanol and stored at -80°C. Kynurenate (200 mM) was prepared in distilled water and stored at -20°C. Retigabine (10 mM), carbamazepine (100 mM) and the F4 fraction of A. senegalensis (200 mg/ml) were prepared in DMSO and stored at -20°C. Drugs were thawed and diluted into the extracellular solution before use. DMSO was diluted in the extracellular solution of all control experiments such that the same final concentration of DMSO was present in the extracellular medium when applying any drug.
Hippocampal slice cultures
Hippocampal slice cultures were prepared by the method described by Debanne et al. (2008). Briefly, rats (postnatal day 7-8) were anesthetized with chloral hydrate and decapitated. Hippocampal slice cultures are prepared by dissecting slices from the brain region of interest. Hippocampal slices (350 µm) were maintained in appropriate culture conditions for up to 21 days in an incubator at 34°C, 95% O2-5% CO2. To limit glial proliferation, 5 µM Ara-C was added to the culture medium starting at 3 DIV. LGI1 or heat-denatured LGI1 (100 nM) was applied to slice-cultures for 24 to 72 h. These cultures have been prepared from a variety of brain regions and maintain general morphology of the originating brain region during the culture period.
Electrophysiology
Individual slice cultures were transferred to a recording chamber perfused at 2 to 3 ml/min with artificial cerebrospinal fluid (ACSF). Whole cell recordings were obtained from CA3 pyramidal cells in organotypic cultures. Recordings of excitatory postsynaptic current and potentials (EPSC and EPSPs) were made using a Multiclamp 700B amplifier (Axon Instruments, Molecular Devices). Only neurons with a resting membrane potential more negative than -60 mV were kept for final analysis.
Dynamic clamp recordings
To add artificial Nav channel, the allosteric model proposed by Kuo and Bean (1994)was used. For this purpose, a fast dynamic-clamp system was used with National Instruments data acquisition cards (PCI-6052E & PCI-6251) and NI-DAQmx 9.21, controlled by the QuB software (http://www.qub.buffalo.edu/; Milescu et al., 2010, 2008). The dynamic-clamp system was running on an independent personal computer mounted with a dual eight-core Xeon processor at 2.67 GHz. The feedback loop (90 kHz) continuously read the Vm from the MultiClamp 700B, and the DC command from the PCI-6723 NIDAQ card controlled by IGOR Pro software on a separate computer and generated an output according to the equation Isum = gNa·(Vm-ENa) + DC. ENa was set at +62 mV. The other parameters used in the simulation were the same as the one used by Milescu et al. (2010).
Stimuli
Direct-Current (DC) steps were generated using IGOR Pro software (WaveMetrics) and converted to analog signals via a PCI-6723 National Instruments.
Electrical stimulations (20-80 µs; 10-50 V) were delivered with a bipolar tungsten electrode placed in the stratum radiatum of the CA3 region.
Anticonvulsant properties of the F4 fraction
In order to test the anticonvulsant properties of A. senegalensis, epileptiform activity was induced by blocking GABAergic inhibition with Picrotoxin (PTX, 100 µM). Spontaneous epileptiform discharges could be observed after extracellular perfusion of PTX.
Epileptiform discharges could also be evoked when placing an electrode in the CA3 region of the hippocampus, in order to activate the CA3 recurrent collaterals and the mossy fibers of the hippocampus. To validate this model of epilepsy in vitro and the ability of anticonvulsant agents to reduce this type of neuronal events, the effects of carbamazepine (CBZ, 10 µM), an active drug used in the treatment of partial were tested and generalised epilepsy in humans (Beydoun et al., 2020).
Modulation of intrinsic excitability by the Annona F4 fraction
In the following experiments, the second of these two assumptions were evaluated, namely the possibility that the extracellular presence of the F4 fraction of A. senegalensis affects the genesis of action potentials. We therefore evaluated the intrinsic excitability of pyramidal cells in the CA1 region of the hippocampus. Experiments were performed in the presence of kynurenate (200 µM) and PTX (100 µm) in order to block inotropic glutamatergic and GABAergic synaptic activity, respectively. In current-clamp mode, depolarizing current pulses of various amplitudes was injected into the recorded cells.
Effects of the Annona F4 fraction on voltage-gated sodium channels
In the next experiments, it was reasoned that if the F4 fraction was targeting voltage-dependent TTX sensitive sodium channels, its effects should not be any more detected if these channels were blocked by the presence of TTX, but the intrinsic excitability restored by the Dynamic Clamp injection of currents mimicking sodium currents simulated according to the voltage fluctuations overtime of the recorded cell.
Analysis
Synaptic activity and spike analysis were performed with Igor Pro (Wavemetrics, Lake Oswego, OR, USA). A collection of Igor Pro functions (Neuromatic, Jason Rothman, http://www.neuromatic.thinkrandom.com) was used in addition to homemade functions.
Spike threshold was detected when dV/dt exceeded 10 mV/ms. Changes in excitability were evaluated for equivalent DC steps and then averaged for each cell. In Figures 1 and 2, dose response curves were fitted with a Hill equation.
Pooled data are presented as mean ± SE and statistical analysis was performed using the Mann-Whitney U test or Wilcoxon rank signed test.
RESULTS
Phytochemical test of the extract
Phytochemical screening of the F4 fraction indicated the presence of the following constituents: terpenoids, flavonoids, and diterpenoids.
Anticonvulsant properties of the F4 fraction in an in vitro model of epilepsy
After a control period during which the epileptiform discharges were evoked in the presence of PTX (100 µM), CBZ was added to the extracellular medium. After 10 min, a decrease in the duration of discharges and the number of action potentials emitted during each discharge was observed (Figure 1A). CBZ reduced in a dose-dependent manner the duration of evoked epileptiform bursts (Figure 1B). At 1, 10 and 100 and 300 µM epileptiform activity was reduced to 94 (n=3), 79 (n=7), 49 (n=4) and 12 (n=3)% of control, respectively.
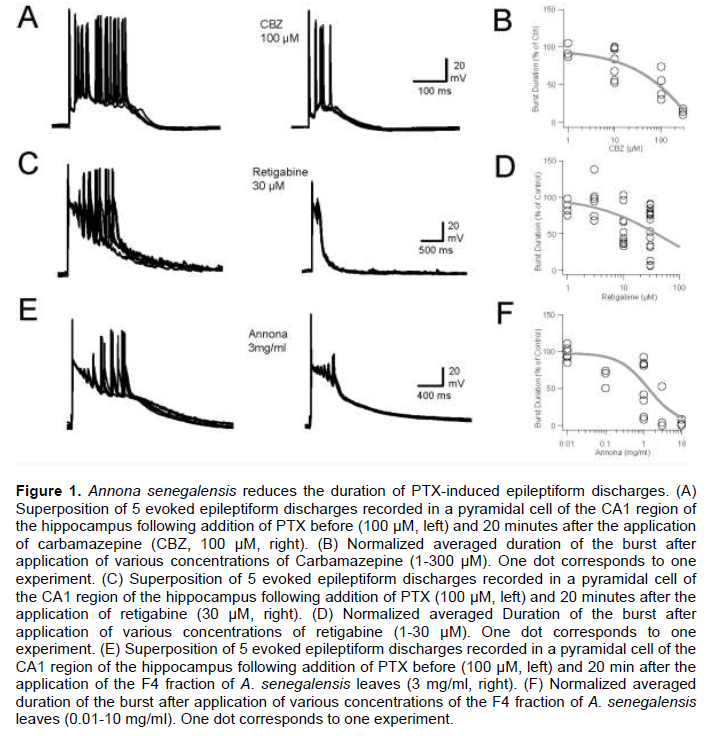
The effects of retigabine were also tested on epileptiform activity (Figure 1C). Retigabine reduced in a dose dependent-manner the duration and the number of spikes emitted during epileptiform discharges (Figure 1C and D). At 1, 3, 10 and 30 µM, epileptiform activity was reduced to 86 (n=4), 95 (n=6), 69 (n=13) and 55 (n=15)% of control, respectively.
Different concentrations of the Annona F4 fraction were tested on the control of epileptiform discharges induced in vitro by PTX (100 µm, Figure 1E). Like CBZ and retigabine, the effects of the Annona F4 fraction on epileptiform activity were dose-dependent. At 0.01, 0.1, 1, 3 and 10 mg/ml, epileptiform activity was reduced to 97 (n=6), 65 (n=3), 54 (n=8), 14 (n=4) and 3 (n=3)% of control, respectively.
These results showed that the Annona F4 fraction has interesting anticonvulsant properties, given the fact that at 0.1 to 1 mg/ml, the reduction of epileptiform activity was quantitatively and qualitatively similar to CBZ (100 µM) or retigabine (30 µM) in vitro. At larger doses of epileptiform, discharges were not detected in the CA1 region of the hippocampus, suggesting that the spread of network activity was disrupted, either due to a blockage of synaptic glutamatergic transmission and/or that the genesis of action potentials was fully abolished.
Modulation of intrinsic excitability by the Annona F4 fraction
Thus, input-output curves were determined under control conditions and after application of various concentrations of CBZ, retigabine or the Annona F4 fraction (Figure 2).

CBZ reduced intrinsic excitability in a dose-dependent manner. Indeed, CBZ reduced intrinsic excitability to 100% (n=3), 92% (n=8), 46% (n=5) and 9% (n=3) at concentrations of 1, 10, 100 and 300 µM, respectively (Figure 2A). Next, the effects of retigabine on intrinsic excitability were evaluated. Extracellular application of retigabine decreased the intrinsic excitability of CA1 pyramidal cells in a dose-dependent manner. At the lowest concentration tested, 1 µM retigabine increased the intrinsic excitability to 108% of control (n=10). At 3, 10 and 30 µM, retigabine reduced intrinsic excitability to 67% (n=7), 29% (n=10) and 33% (n=10), respectively (Figure 2C and D).
Similar experiments were performed with A. senegalensis (Figure 2E and F). Annona F4 fraction reduced CA1 pyramidal cell excitability in a dose-dependent manner. At the lower concentration tested (0.01 mg/ml) Annona F4 fraction reduced the intrinsic excitability to 82% of control (n=6). At 0.1, 1, 3 and 10 mg/ml, the F4 fraction reduced intrinsic excitability to 87% (n=8), 76% (n=5), 42% (n=6) and 2% (n=3) of control.
Thus A. senegalensis F4 fraction has the ability to reduce intrinsic excitability of CA1 pyramidal neurones. As demonstrated by CBZ and retigabine, which modulate different ion channel targets, namely sodium and KCNQ potassium channels. The acute effects of the F4 fraction could be explained by affecting at least one of these two channels. Thus in the next series of experiments, attempt was made to decipher between these two putative targets.
Effects of the Annona F4 fraction on voltage-gated sodium channels
In the next experiments, we reasoned that if the F4 fraction was targeting voltage-dependent TTX sensitive sodium channels, its effects should not be any more detected if these channels were blocked by the presence of TTX, but the intrinsic excitability restored by the Dynamic Clamp injection of currents mimicking sodium currents simulated according to the voltage fluctuations overtime of the recorded cell (see methods). Thus, it was possible to record and measure the intrinsic excitability of a CA1 pyramidal neuron where endogenous TTX sensitive sodium channels were pharmacologically blocked (Figure 3A). In these conditions, application of the F4 fraction of A. senegalensis (3 mg/ml) did not alter intrinsic excitability (Figure 3B and C).

On average, when compared to control (that is, absence of TTX; 42 ± 13 n=6), the excitability was 91 ± 10 (n=5). In order to validate the use of Dynamic Clamp as a method to specifically address TTX sensitive sodium channels as a pharmacological target, we tested and compared the effects of retigabine on intrinsic excitability of Control and TTX-dynamic Clamped neurones. Retigabine (10 µM) reduced to a same extent intrinsic excitability in both conditions (P>0.05).
Thus, when TTX sensitive sodium channels were blocked, the F4 fraction of A. senegalensis did not alter anymore the intrinsic excitability of CA1 pyramidal neurons restored with a dynamic clamp amplifier for TTX sensitive sodium channels whilst pharmacological compounds targeting other types of channels could still affect intrinsic excitability. Our data therefore suggest that the F4 fraction of A. Senegalensis targets TTX sensitive sodium channels.
DISCUSSION
Traditional medicine has always been used in Burkina Faso and other African countries for the prevention and treatment of various diseases with few adverse effects (Gurib-Fakim, 2006). Among several traditional claims, the usefulness of A. senegalensis in the management of febrile convulsions has been manifest (Konate et al., 2012a, b; Okoli et al., 2010).
The result of preliminary phytochemical screening revealed the presence of various secondary metabolites including, alkaloids, tannins, flavonoid, saponins, glycosides, anthocyanins, triterpenes steroids and volatile oil (Jaiya et al., 2014; Mpiana, 2012; Yisa et al., 2010). These phytochemical constituents have been reported to be associated with different pharmacological activities (Bukhari et al., 2013; Chen et al., 2013). Flavonoid, triterpenes and steroids, among other phytochemicals have been reported to possess anticonvulsant activity (Sayyah et al., 2004). Bioactivity-guided fractionation of A. senegalensis using pentylenetetrazole (PTZ) induced seizures in mice resulted in the characterization of kaur-16-en-19-oic acid (KA), a diterpenoid. KA dose-dependently delayed onset of myoclonic spasms and tonic-clonic phases of PTZ-induced seizures and maximal electroshock seizures (Theophine et al., 2013; Nemesio et al., 2016). In previous investigations, extracts of A. senegalensis have been reported to have significant anticonvulsant effects; however, no related active compounds have been reported (Konate et al., 2012a, b; Okoli et al., 2010).
In the present study, we explored the mechanisms of action of the F4 fraction from the extract of A. senegalensis which was reported to have significant anticonvulsant activity (Konate et al., 2012a, b). The data presented in this study provided scientific evidence that this fraction significantly reduced the hippocampal neuronal excitability at the network level.
Conventional antiepileptic drugs may improve GABA function or block sodium channels. K+ channels play a significant role in controlling all aspect of neuronal excitability (Wickenden, 2002). Voltage-gated Na+ channels blockers such as carbamazepine, phenytoin and lamotrigine which prevent seizure spread effectively antagonize seizures induced by K+ channel blocker such as 4-AP (Subramaniam et al., 1995; Ragsdale and Avoli, 1998).
The effects of A. senegalensis on epileptiform activity were similar to carbamazepine and retigabine, two anticonvulsant drugs that target sodium (McLean and MacDonald, 1986; Ragsdale et al., 1991)and KCNQ channels (Tatulian et al., 2001), respectively. While sodium channels are directly involved in the genesis of action potential, KCNQ channels have been shown to modulate both neuronal excitability (Devaux et al., 2004)and neurotransmitter release (Martire et al., 2004). While in our two first sets of experiments, we could not conclude on the channels that are modulated by the Annona F4 fraction, the experiments performed with a dynamic clamp amplifier programmed to simulate the voltage-dependent sodium currents suggested strongly that Annona F4 fraction may target and inhibit the voltage-dependent sodium channels.
From the results presented earlier, we can conclude that, as suggested in vivo (Konate et al., 2012a, b; Okoli et al., 2010), A. senegalensis possess interesting anticonvulsant properties that seem to be mediated through at least the modulation of voltage-dependent sodium channel. Piperine, a natural compound isolated from the fruits of Piper, delays the onset of seizures in various drug-induced convulsions in rodents and associated mortality (Bukhari et al., 2013; Chen et al., 2013). Different mechanisms have been proposed for piperine. Among them, a recent study strongly suggested that Na+ channel antagonist activity may be an important contributor to anticonvulsant mechanisms of piperine (Mishra et al., 2015). Voltage-gated Na+ channels are key players in cellular excitability and they are the therapeutic target of antiepileptic drugs (AEDs) like carbamazepine, phenytoin and lamotrigine. These drugs modulate voltage-gated Na+ channels in a use-and voltage-dependent manner which allows them to selectively prevent high frequency firing, with little effect on a single action potential. Carbamazepine, lamotrigine and phenytoin all have a much higher affinity for the inactivated state than for the closed and open states of the Na+ channel. Therefore, they stabilize the inactivated state, effectively blocking the Na+ conductance and delaying recovery from inactivation, which prevents synchronized high frequency firing (Ragsdale and Avoli, 1998).
In order to conclude that sodium channel antagonist activity mays act as a major contributor to the anticonvulsant mechanisms of senegalensis, like piperine (Mishra et al., 2015), some voltage-clamp experiments of pharmacologically isolated sodium channels in CA1 pyramidal cells will be required. Cell lines expressing sodium channels could also be used to confirm the voltage-dependent channels targeted by A. senegalensis and the biophysical mechanisms by which these channels are antagonized.
CONCLUSION
In the present study, the pharmacological profile of the most promising A. senegalensis fraction obtained up-to-date with anticonvulsant drugs targeting voltage-activated channels namely Carbamazepine (Na+ channels) and Retigabine (KCNQ Channels) were compared.
Using an in vitro model of temporal lobe epilepsy, the dose-dependent effects of A. senegalensis on electrically evoked epileptiform bursts in the CA1 region of the rat hippocampus with carbamazepine and retigabine were compared. The dose-dependent effects of A. senegalensis on intrinsic excitability of CA1 pyramidal neurons were also evaluated and compared to carbamazepine and retigabine. Using a dynamic clamp amplifier, a last series of experiments were designed in order to evaluate the hypothesis that anti-epileptic effects of A. senegalensis may be due to a sodium channel antagonism activity. The data suggest that A. senegalensis contains molecules that target voltage-dependent sodium channels.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
This work was supported by the Institut National de la Santé et de la Recherche Médicale (INSERM), the Centre National pour la Recherche Scientifique (CNRS), with a grant from the Fondation Française pour la Recherche sur l’Epilepsie (FFRE), the French Ministry for Europe and Foreign Affairs. Parts of the equipment used in this study were funded by European Community (LSHM-CT-2004-511995), Agence Nationale pour la Recherche (ANR 06-Neuro-014-01) and région PACA (APO ‘Plexin') grants attributed to D. Debanne. They thank the members of INSERM UMR_S 1072 for helpful comments and suggestions during completion of this work, and other for critical reading of the manuscript.
REFERENCES
|
Beydoun A, DuPont S, Zhou D, Matta M, Nagire V, Lagae L. (2020). Current role of carbamazepine and oxcarbazepine in the management of epilepsy. Seizure 83:251-263. |
|
|
Bukhari IA, Pivac N, Alhumayyd MS, Mahesar AL,Gilani AH. (2013). The analgesic and anticonvulsant effects ofpiperine in mice. Journal of Physiology and Pharmacology 64(6):789-94. |
|
|
Bruno E, Bartoloni A, Zammarchi L, Strohmeyer M, Bartalesi F, Bustos JA (2013). COHEMI Project Study Group. (Epilepsy and neurocysticercosis in Latin America: a systematic review and meta-analysis. PLoS Neglected Tropical Diseases 7(10):e2480. |
|
|
Britton JW, So EL (1995). New antiepileptic drugs: prospects for the future. Journal of Epilepsy 8:267-81 |
|
|
Chen CY, Li W, Qu KP, Chen CR (2013). Piperine exertsanti-seizure effects via the TRPV1 receptor inmice. European Journal of Pharmacology 714:288-94. |
|
|
Debanne D, Boudkkazi S, Campanac E, Cudmore RH, Giraud P. (2008). Paired-recordings from synaptically coupled cortical and hippocampal neurons in acute and cultured brain slices. Nature Protocols 3:1559-68 |
|
|
Devaux JJ, Kleopa KA, Cooper EC, Scherer SS (2004). KCNQ2 is a nodal K+ channel. Journal of Neuroscience 24:1236-44. |
|
|
Fisher RS, Acevedo C, Arzimanoglou A, Bogacz A, Cross JH, Elger CE, Engel J, Forsgren L, French JA, Glynn M, Hesdorffer DC, Lee BI, Mathern GW, Moshe SL, Perucca E, Scheffer IE, Tomson T, Watanabe M, Wiebe S (2014). ILAE official report: a practical clinical definition of epilepsy. Epilepsia 55:475-482. |
|
|
Gurib-Fakim A (2006). Medicinal plants: traditions of yesterday and drugsof tomorrow. Molecular Aspects of Medicine 27(1): 1-93 |
|
|
Jaiya I, Arzika S, Abdulkadir M (2014). Extraction and Phytochemical Screening of the Root and Leave of Annona senegalesis (Wild Custad Apple). Academic Journal of Interdisciplinary Studies 3(7):9-15. |
|
|
Konate A, Sawadogo WR, Dubruc F, Caillard O, Guissou IP. (2012°). Anticonvulsant effects of the Stem bark extract of Annona senegalensis Pers. Molecular and Clinical pharmacology 3:62-72 |
|
|
Konate A, Sawadogo WR, Dubruc F, Caillard O, Ouedraogo M, Guissou IP (2012b). Phytochemical and Anticonvulsant Properties of Annona senegalensis Pers.(Annonaceae), Plant Used in Burkina Folk Medicine to Treat Epilepsy and Convulsions. British Journal of Pharmacology and Toxicology 3:245-250 |
|
|
Kuo CC, Bean BP (1994). Na+ channels must deactivate to recover from inactivation. Neuron 12:819-829 |
|
|
Malagon-Valdez J (2004). The new antiepileptic drugs: their indications and side effects. Revue Neurologique 39:570-575 |
|
|
Martire M, Castaldo P, D'Amico M, Preziosi P, Annunziato L, Taglialatela M (2004). M channels containing KCNQ2 subunits modulate norepinephrine, aspartate, and GABA release from hippocampal nerve terminals. Journal of Neuroscience 24:592-97 |
|
|
McCormick DA, Contreras D (2001). On the cellular and network bases of epileptic seizures. Annual Review of Physiology 63:815-846 |
|
|
McLean MJ, MacDonald RL (1986). Carbamazepine and 10,11-epoxycarbamazepine produce use- and voltage-dependent limitation of rapidly firing action potentials of mouse central neurons in cell culture. Journal of Pharmacology and Experimental Therapeutics 238:727-38 |
|
|
Milescu LS, Bean BP, Smith JC (2010). Isolation of somatic Na+ currents by selective inactivation of axonal channels with a voltage prepulse. Journal of Neuroscience 30:7740-7748 |
|
|
Milescu LS, Yamanishi T, Ptak K, Mogri MZ, Smith JC (2008). Real-time kinetic modeling of voltage-gated ion channels using dynamic clamp. Biophysical Journal 95: 66-87 |
|
|
Mishra A, Punia JK, Bladen C, Zamponi GW, Goel RK (2015). Anticonvulsant mechanisms of piperine, a piperidine alkaloid. Channels (Austin) 9:317-23 |
|
|
Mpiana P (2012). Antisickling properties, thermal and photochemical degradations of anthocyanin extracts from Annona senegalensis (Annonaceae) International Journal of Biological and Chemical Sciences 6(5):2241-2251. |
|
|
Nemesio VR, Edmundo LG,Yesenia PH (2016). Anticonvulsant effect of kaurenoic acid isolated from the root bark of Annona senegalensis Pharmacology Biochemistry and Behavior 109:38-43. |
|
|
Okoli CO, Onyeto CA, Akpa BP, Ezike AC, Akah PA, Okoye TC (2010). Neuropharmacological evaluation of Annona senegalensis leaves. African Journal of Biotechnology 9:8435-8444. |
|
|
Perucca P, Mula M (2013) Antiepileptic drug effects on mood and behavior: molecular targets. Epilepsy and Behavior 26(3):440-449. |
|
|
Privitera M (2011). Current challenges in the management of epilepsy, American Journal of Managed Care 17(Suppl. 7):S195-203. |
|
|
Ragsdale DS, Scheuer T, Catterall WA (1991). Frequency and voltage-dependent inhibition of type IIA Na+ channels, expressed in a mammalian cell line, by local anesthetic, antiarrhythmic, and anticonvulsant drugs. Molecular Pharmacology 40:756-65. |
|
|
Ragsdale DS, Avoli M (1998). Sodium channels as molecular targets for antiepileptic drugs. Brain Research. Brain Research Reviews. 26:16-28. |
|
|
Rostock A, Tober C, Rundfeldt C, Bartsch R, Engel J (1996). D-23129: a new anticonvulsant with a broad spectrum activity in animal models of epileptic seizures. Epilepsy Research 23:211-23. |
|
|
Stafstrom CE, Carmant L (2015). Seizures and epilepsy: an overview for neuroscientists. Cold Spring Harb Perspect Medicine 5(6):a022426. |
|
|
Sayyah M, Nadjafnia L, Kamalinejad M (2004). Anticonvulsant activity and chemical composition of Artemisia dracunculus L. essential oil. Journal of Ethnopharmacology 94(2-3):283-287. |
|
|
Subramaniam S, Rho JM, Penix L, Donevan SD, Fielding RP, Rogawski MA (1995). Felbamate block of the N-methyl-D-aspartate receptor. Journal of Pharmacology and Experimental Therapeutics 273(2):878-886. |
|
|
Theophine C, Okoye,Peter A, Akah,Edwin O, Omeje, Festus BC, Okoye Chukwuemeka Nworu S (2013). Anticonvulsant effect of kaurenoic acid isolated from the root bark of Annona senegalensis Pharmacology Biochemistry and Behavior 109.38-43. |
|
|
Tatulian L, Delmas P, Abogadie FC, Brown DA (2001). Activation of expressed KCNQ potassium currents and native neuronal M-type potassium currents by the anti-convulsant drug retigabine. Journal of Neuroscience 21(15):5535-5545. |
|
|
Yemadje LP, Houinato D, Quet F, Druet-Cabanac M, Preux PM (2011). Understanding the differences in prevalence of epilepsy in tropical regions. Epilepsia 52(8):1376 1381. |
|
|
Yemitan OK, Adeyemi OO (2013). Antiepileptogenic and anticonvulsant actions of Dalbergia saxatilis (Hook, F.) in sub-toxic chemical kindling and toxic convulsant models. European Journal of Medicinal Plants 3(2):288-296. |
|
|
Yisa J, Egila J, Darlinton A (2010). Chemical composition of Annona senegalensis from Nupe land, Nigeria. African Journal of Biotechnology 9(26):4106-4109. |
|
|
Wickenden AD (2002). Potassium channels as antiepileptic drug targets. Neuropharmacology; 43(7):1055-1060. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


