Full Length Research Paper

ABSTRACT
Insulin-degrading enzyme (IDE) is the major zinc-metalloprotease involved in the cleavage of insulin, β-amyloid protein and mainly contributes to the pathophysiology of type 2 diabetes and Alzheimer’s disease. Therefore, this enzyme is expressed as candidate target for drugs used in the management of diabetes and peptide hydroxamates have been reported recently as inhibitors for IDE. Novel synthesized peptide hydroxamic acid II containing tryptophan and a sulfonamide bond has been prepared in our laboratory. The aim of this study was to determine whether this drug could be of value in modulating diabetic states in rats. In this study, forty adult male Wistar albino rats received 20% fructose in drinking water (HFW) for six weeks to induce diabetes. Administration of the prepared compound at two dose level (5 and 10 mg/kg body weight, p.o) to diabetic rats significantly reduced IDE protein, glucagon levels, improved insulin receptor signaling (phosphorylation), insulin sensitivity and lipid profile. However, it induced certain up-regulation of IDE mRNA expression. These findings may confirm its role in the modulation of glucose homeostasis through IDE and insulin receptor signaling.
Key words: Insulin degrading enzyme, insulin receptors phosphorylation, insulin resistance, glucagon, hyperlipidemia.
INTRODUCTION
Pancreatic beta cells[C31] failure is the major contributing factor to the development of type 2 diabetes since insulin resistance of target tissues is usually associated with abnormal insulin secretion. On cellular level, the inadequate insulin signaling from insulin receptor downstream to the final substrates of insulin action may lead to multiple metabolic pathways (Iliya et al., 2016).
Insulin receptor is a hetero tetramer consisting of 2 alpha subunits and 2 beta subunits that are linked by disulphide bonds. Insulin binds to the 2 alpha subunits of insulin receptors and activates tyrosine kinase in the beta subunits. Once it is activated, tyrosine kinase promotes autophosphorylation of the beta subunits where phosphorylation of tyrosine residues is required for amplification of the kinase activity. Most of the metabolic and antiapoptotic effects of insulin are mediated by the signaling pathway involving the phosphorylation of insulin receptor substrate protein and the activation of protein kinase B. In[C32] humans, rare mutation of the insulin receptor substrate-1 protein is associated with insulin resistance (Junlan and Gangjian, 2012).
Insulin-degrading enzyme (IDE) is a structurally distinctive zinc metalloprotease responsible for catabolizing insulin and other intermediate-sized peptide substrates. In principle, it should be possible to enhance the activity of insulin by inhibiting its catabolism (Deprez-Poulain et al., 2015). Thus, suitable pharmacological inhibitors of IDE may hold therapeutic value, particularly for type 2 diabetes mellitus (T2DM) and other disorders involving impaired insulin signaling (Maianti et al., 2014).
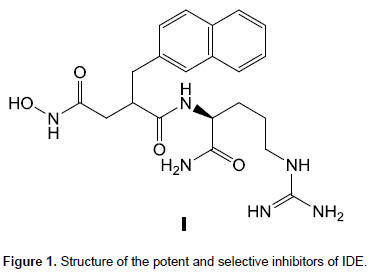
Peptide hydroxamic acids were previously reported as potent and selective inhibitors of IDE, for example, compound I showed excellent potency (ki = 2.96 ± 0.20 nM) and good selectivity (∼104-fold) vis-aÌ€-vis other zinc-metalloproteases and representative member of other protease classes (Abdul-Hay et al., 2013) (Figure 1).
Herein, the synthesis and antidiabetic activity of a novel peptide hydroxamic acid II containing typtophane and a sulfonamide bond through examination of IDE gene expression and protein level as well as insulin receptor phosphorylation were reported (Figure 2).

MATERIALS AND METHODS
Chemistry
Synthesis of [C31] (S)-N-(1-(1H-benzo[d][1,2,3]triazol-1-yl)-3-(1H-indol-3-yl)-1-oxopropan-2-yl)-4- methylbenzenesulfonamide (III)
A mixture of p-toluene sulfonyl chloride (0.95 g, 5 mmol) and DMAP (0.08 g, 0.65 mmol) was stirred in methylene chloride (5 ml) for 10 min. Compound II (1.8 g., 5 mmol) was dissolved in methylene chloride (5 ml) containing trimethylamine (TEA; 0.21 ml, 1.5 mmol) and the resulting solution was added to the reaction mixture. After 20 min, benzotriazole (0.72 g, 6 mmol) was added and the reaction was allowed to stir for additional 1.5 h at 25°C. Upon completion of the reaction (monitored by thin layet chromatography [TLC]), CH2Cl2 (50 ml) was added and the organic layer was washed with saturated Na2CO3 (10 ml, 3×), water (10 ml, 2×) and brine[C32] (10 ml, 1×). The organic layer was dried over anhydrous sodium sulfate and hexane (20 ml) was added. The solid separated was filtered and dried under vacuum to give the target N-acylbenzotriazoles (2.1 g, 92%).
Synthesis of (S)-N-hydroxy-3-(1H-indol-3-yl)-2-(4-methylphenyl-sulfonamido) propanamide (II)
To a solution of compound III (1.37.g, 3 mmol) in methylene chloride (20 ml), hydroxylamine HCl (0.42 g, 6 mmol[C33] ) and TEA (0.84 ml, 6 mmol) were added. The mixture was stirred at 25°C for 1 h. The reaction was diluted with methylene chloride (40 ml), and then was washed with 6N HCl[C34] (3 × 10 ml), water (2 × 10 ml) and brine (1 × 10 ml). The organic layer was dried over anhydrous sodium sulfate and was filtered. The filtrate was evaporated under vacuum to afford compound II (yield 1.00 g, 90%).
Animals and ethics
Adult male Wistar albino rats (weighing 160±20 g) were housed in stainless steel cages in a controlled environment (23°C and with a 12-h light/dark cycle). All experimental procedures were performed according to the guidelines of the Animal Care and Use Committee of Zagazig University, Egypt.
Experimental design
Forty rats were randomly assigned into the following four groups, n=10, for 45 days. The first one was kept on normal rat chow [C35] diet (El-Nasr Pharmaceuticals and Chemicals Industry, Egypt), normal tap water and no treatment (Normal control group). The other three groups received normal rat chow diet, 20% fructose in drinking water (HFW) daily to induce type 2 diabetes (Mamikutty et al., 2014). It has been reported that long term fructose feeding to genetically selected albino rats result in the development of diabetes mellitus and glomerulosclerosis [6]. One of these groups received no treatment and served as diabetic control group, while the others were co-treated orally with the synthesized peptide hydroxamic acid II containing typtophane and a sulfonamide bond in dose levels of 5 and 10 mg/kg body weight/day (Hydroxamic acid 5 group, Hydroxamic acid 10 group) for 45 days. Selection was based on a pilot study conducted in our laboratory where they produced optimal effects on the selected biomarkers without side effects. However, a higher dose of 15 mg/kg body weight/day, p.o produced deleterious effects and marked mortality.
Blood and tissue sampling
The rats were fasted overnight and supplemented with only tap water[C36] , blood samples were obtained at the baseline and at end of the experiment via orbital vein of anaesthetized rats. Animals were weighed before collection of blood samples. Blood was centrifuged at 4500 rpm and serum samples were immediately analyzed for glucose. The remaining serum was frozen at -80°C and stored for further analysis of lipid profile, insulin and glucagon. Rats were then killed by decapitation and the liver was excised, washed with cold saline, kept in liquid nitrogen and stored at -80°C for measurement of IDE gene expression and protein level as well as insulin receptor phosphorylation in cell lysate..
Biochemical measurements
Colorimetric kits (Spinreact, Spain) were used for determination of glucose, total cholesterol (TC), triacylglycerol (TAG) and high density lipoprotein cholesterol (HDL-C) levels. Low density lipoprotein cholesterol (LDL-C) was calculated from Friedewald formula: LDL-C= TC-(TAG/5+HDL-C). Enzyme-linked immunosorbent assay (ELISA) kits for rat insulin, glucagon, insulin receptor phosphorylation, and IDE were purchased from SPI Bio, Montigny Le Bretonneux, France; CUSABIO, China; Sigma Aldrich, MO, USA and Cloud-Clone Crop, assembled by USCN Life Science Inc., Wuhan, China, respectively. All biochemical analyses were done following the manufacturer’s protocols.
Quantitative real time polymerase chain reaction (q-PCR) for IDE
Total RNA was isolated from the livers of animals in the studied groups using Qiagen tissue extraction kit (Qiagen, USA) and RNA was converted into cDNA using high capacity cDNA reverse transcription kit (Fermentas, USA). The primers sequence for IDE and GAPDH genes are shown in Table 1. The cDNA was subjected to amplification and analysis using the TaqMan Master Mix (Applied Biosystems, Foster City, CA, USA) for quantitative RT-PCR. GAPDH was used as the reference gene. The assay was run on Step One™ Real-Time PCR Systemversion 3.1 (Applied Biosystems, USA) according to the manufacturer's instructions.

Statistical analysis
Statistical analysis was done[C31] using the InStat 2.04 statistical package (Graph Pad InStat). Data are presented as means ± standard deviation (SD) for six rats/group. Student’s t test was used to compare means between each two groups. P<0.05 was considered to be statistically significant.
[C31]Delete space
[C31]Remove space
[C32]Space
[C33]Space
[C34]Space
[C35]specify type, brand and manufacturer
[C36]Delete space
RESULTS
Chemistry
The target compound II was prepared according to the synthetic pathway, which is depicted in scheme 1. L-Tryptophan 1 was sulphonylated using tosyl chloride in the presence of potassium carbonate to give N-tosyl-L-tryptophan (2). Compound 2 was converted to its corresponding benzotriazolide 3 (90% yield) via reaction with 1H-benzotriazole and tosyl chloride in the presence of equivalent amount of TEA (Agha et al., 2016). Reaction of 3 with hydroxylamine HCl in the presence of TEA afforded the target compound II in 95% yield. Intermediates 2 and 3 as well as the target II were characterized by 1HNMR, 13CNMR, and elemental analysis.
Biochemical markers
Intake of fructose solution (HFW, 20%) induced higher adiposity, hyperglycemia, dyslipidemia, and insulin resistance. Serum glucagon, hepatic content of IDE showed significant increase along[C31] with down-regulation of IDE gene expression and a significant decrease of insulin receptor phosphorylation. Concomitant administration of hydroxamic acid peptide II at dose levels (low and high) greatly ameliorated these changes by improving insulin receptor phosphorylation and insulin sensitivity as well as IDE reduction[C32] (Tables 2 and 3). Unexpectedly, IDE mRNA level was significantly increased (Figure 3).



DISCUSSION
Rats fed on HFW in the current study developed a state of metabolic syndrome as manifested by hyperglycemia, dyslipidemia, and body weight gain (obesity). The excess calorie intake here is finally stored in the abdominal adipose tissues as TAG (Karamohamed et al., 2003) leading to the development of dyslipidemia and hyperglycemia. Adipose tissue usually acts as an endocrine organ which involves the metabolism of glucocorticoid hormone where deregulation of glucocorticoid metabolism as reported earlier may lead to obesity, dyslipidemia, hypertension, and diabetes (Kershaw and Flier, 2004).
It is generally known that sensitivity of insulin is reduced with the presence of TAG, inducing a reduction of glucose uptake by the insulin sensitive tissues leading to activation of lipolysis process, more free fatty acids, glycerol formation indeed TAG accumulation within the adipose tissue (Paschos and Paletas, 2009). These viscous cycles are repeated and more TAG is formed, leading to a hypertriglyceridemic state along with insulin resistance (elevated glucose and insulin).
Patients suffering from insulin resistance and type 2 diabetes frequently display signs of abnormal lipid metabolism, increased circulatory concentration, and elevated deposition of lipids in the skeletal muscles (McGarry, 2001). Increase in plasma free fatty acids reduces insulin-stimulated glucose uptake, whereas a decrease in plasma lipid content improves insulin activity in the skeletal muscle cells, adipocytes, and liver (Moller, 2001). Studies have shown that raising plasma fatty acids in both rodents (Kim et al., 2004) and humans (Dresner et al., 1999) abolishes insulin activation of insulin receptor substrate-1 associated PI3-kinase activity in skeletal muscles where insulin receptor substrate-1 is the most prevalent lipid associated insulin resistance and has been shown to be linked to GLUT4 translocation defects (Pessin et al., 1999).
Hyperglycemia observed here may be attributed to the fact that unlike glucose, fructose does not stimulate insulin secretion from pancreatic β-cell (Bray et al., 2004); additionally, the reduction of insulin sensitivity during hypertriglyceridemia may induce hyperglycemia. The present study demonstrated an increase of liver IDE protein level in rats fed on HFW (20%). This may be attributed to a direct response for a negative feedback mechanism due to an alteration of circulated insulin. However, there is no sufficient information about IDE regulation in obesity (Duckworth et al., 1998; Hulse et al., 2009). Inactivation of IDE by gene knockout induced hyperinsulinemia and insulin resistance in mice[C31] as previously reported may add further support[C32] [C33] (Wei et al., 2014). Increased[C34] glucagon level recorded[C35] in the diabetic rats may[C36] [C37] induce IDE protein as previously reported (Wei et al., 2014).
Probable biochemical effects of our compound may be mediated through different mechanisms. The first one is through enhancing glucose uptake by liver, muscle, and adipose tissue; although, this is not done in most studies. The second one may be attributed to parathyroid hormone (PTH) role where several studies indicated that[C38] PTH may elicit insulin resistance by reducing the number of glucose transporter available in cell membranes responsible for glucose uptake (Sung et al., 2012; Teegarden and Donkin, 2009). Additionally, it suppresses insulin release (Perna et al., 1990) and promotes insulin resistance in adipocytes (Teegarden and Donkin, 2009).
It may have a beneficial effect on insulin action either directly by stimulating the expression of insulin receptor, enhancing in turn insulin responsiveness for glucose transport or indirectly via its role in regulating extracellular calcium (Maestro et al., 2000), ensuring its influx through cell membranes and an adequate intracellular calcium pool because calcium is essential for insulin-mediated intracellular processes in insulin responsive tissues such as skeletal muscles and adipose tissues (Williams et al., 1990).
Generally[C39] , insulinemia is a result of the balance between the insulin produced and secreted by pancreatic islets β-cells and insulin removal from the plasma (insulin clearance) (Duckworth et al., 1998). Insulin clearance is fundamentally controlled by the liver and[C310] more than 50% of insulin secreted is removed by the liver after the second passage through the portal vein (Butterfield, 1970; Kotronen et al., 2008; Mittelman et al., 2000). Hepatic removal and degradation of insulin is mainly controlled by IDE (Amata et al., 2009; Duckworth et al., 1998). Every[C311] insulin-responsive cell expresses IDE mainly to oppose insulin signaling by uncoupling insulin from insulin receptor (IR) by removing or degrading insulin (Amata et al., 2009). Accordingly[C312] , hepatic IDE inhibition increased insulin sensitivity coupled with increased IR activity (Li et al., 2002; Leissring et al., 2010). Therefore, IDE expression and its action have certain role in glucose hemostasis, since IDE reduces insulinemia by activation or increasing insulin removal in the liver. Additionally, it reduces insulin sensitivity by interrupting the IR signaling pathway.
Many[C313] studies indicated that IDE expression and insulin clearance are reduced in diabetic individuals especially those having type 2 diabetes (Karamohamed et al., 2003; Kotronen et al., 2008; SlominskiÄ et al., 2009) and also in diabetic and obese rodents (Pessin et al., 1999; Moller, 2001; Matveyenko et al., 2008).
Taken in consideration that[C314] IDE expression and activity represent the major contributing factors to alterations in insulin clearance; the latter can also be affected by certain factors like the changes in renal blood flow or other physiological conditions (Duckworth et al., 1998). Finally, the[C315] observed decrease in hepatic IDE expression in the present study is mostly attributed to hypertriglyceridemia produced in subsequent to high-fat diet (HFD) intake since TAG and non esterified fatty acids are inhibitors[C316] for insulin clearance and hepatic IDE expression (Hamel et al., 2003; Kotronen et al., 2008; Yoshii et al., 2006). Another explanation may be through certain cytokines like interleukin-6 (IL-6) which are released by oversized adipose tissue might control insulin clearance and hepatic IDE expression (Rezende et al., 2012).
[C31]Delete space
[C33]It is relevant, reference update.
Look for other more current
[C34]Delete space
[C35]Delete space
[C36]Delete space
[C37]Delete space
[C38]Delete space
[C39]Delete space
[C310]Delete space
[C311]Delete space
[C312]Delete space
[C313]Delete space
[C314]Delete space
[C315]Delete space
[C316]Delete space
CONCLUSION
Rats received HFD developed insulin resistance, hyperglycemia, and dyslipidemia. Hydroxamic acid peptide administration improved such metabolic disturbances as manifested by an inhibition of hepatic IDE level, increased insulin sensitivity coupled with increased insulin receptor phosphorylation.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Abdul-Hay SO, Lane AL, Caulfield TR, Claussin C, Bertrand J, Masson A, Leissring MA. (2013). Optimization of Peptide Hydroxamate Inhibitors of Insulin-Degrading Enzyme Reveals Marked Substrate-Selectivity. J. Med. Chem. 56(6):2246-2255. |
|
|
Agha KA, Abo-Dya NE, Ibrahim TS, Abdel-Aal EH (2016). Efficient synthesis of N-acylbenzotriazoles using tosyl chloride: en route to suberoylanilide hydroxamic acid (SAHA). ARKIVOC 3:161-70. |
|
|
Amata O, Marino T, Russo N, Toscano M (2009). Human insulin-degrading enzyme working mechanism. J. Am. Chem. Soc. 131(41):14804-14811. |
|
|
Bray GA, Nielsen SJ, Popkin BM (2004). Consumption of high-fructose corn syrup in beverages may play a role in the epidemic of obesity. Am. J. Clin. Nutr. 79(4):537-543. |
|
|
Butterfield WJ (1970). Insulin clearance in nondiabetic and diabetic subjects. Panminerva Med. 12(6):233-235. |
|
|
Deprez-Poulain R, Hennuyer N, Bosc D, Liang WG, Enée E, Marechal X, Verdelet T (2015). Catalytic site inhibition of insulin-degrading enzyme by a small molecule induces glucose intolerance in mice. Nat. Commun. 6:1-6. |
|
|
Dresner A, Laurent D, Marcucci M, Griffin ME, Dufour S, Cline GW, Slezak LA, Andersen DK, Hundal RS, Rothman DL, Petersen KF, Shulman GI. (1999). Effects of free fatty acids on glucose transport and IRS-1-associated phosphatidyl inositol 3-kinase activity. J. Clin. Invest. 103:253-259. |
|
|
Duckworth WC, Bennett RG, Hamel FG (1998). Insulin degradation: progress and potential. Endocr. Rev. 19(5):608-24. |
|
|
Hamel FG, Upward JL, Bennett RG (2003). In vitro inhibition of insulin-degrading enzyme by long-chain fatty acids and their coenzyme A thioesters. Endocrinology 144(6):2404-2408. |
|
|
Hulse RE, Ralat LA, Wei-Jen T. (2009). Structure, function, and regulation of insulin-degrading enzyme. Vitam. Horm. 80:635-648. |
|
|
Miriam ET (2015). IDF Atlas: About 415 Million Adults Worldwide Have Diabetes. Medscape. |
|
|
Iliya A, Mohammed B, Akuyam SA, Nok AJ, Bauchi ZM, Tanko M, Timbuak JA, Yusuf B (2016). Molecular mechanisms of insulin signaling. Sub-Saharan Afr. J. Med. 3:3-7. |
|
|
Junlan Z, Gangjian Q (2012). Adipocyte dysfunction and hypertension. Am. J. Cardiovasc. Dis. 2(2):143-149. |
|
|
Karamohamed S, Demissie S, Volcjak J, Liu C, Heard-Costa N, Liu J, Shoemaker CM, Panhuysen CI, Meigs JB, Wilson P, Atwood LD, Cupples LA, Herbert A. (2003). Polymorphisms in the insulin-degrading enzyme gene are associated with type 2 diabetes in men from the NHLBI Framingham Heart Study. Diabetes 52(6): 1562-1567. |
|
|
Kershaw EE, Flier JS (2004). Adipose tissue as an endocrine organ. J. Clin. Endocrinol. Metab. 89(6):2548-2556. |
|
|
Kim JK, Fillmore JJ, Sunshine MJ, Albrecht B, Higashimori T, Kim DW, Liu ZX, Soos TJ, Cline GW, O'Brien WR, Littman DR, Shulman GI. (2004). PKC-theta knockout mice are protected from fat-induced insulin resistance. J. Clin. Invest. 114(6):823-827. |
|
|
Kotronen A, Juurinen L, Tiikkainen M, Vehkavaara S, Yki-Järvinen H. (2008). Increased liver fat, impaired insulin clearance, and hepatic and adipose tissue insulin resistance in type 2 diabetes. Gastroenterology 135(1):122-130. |
|
|
Leissring MA, Malito E, Hedouin S, Reinstatler L, Sahara T, Abdul-Hay SO, Choudhry S, Maharvi GM, Fauq AH, Huzarska M, May PS, Choi S, Logan TP, Turk BE, Cantley LC, Manolopoulou M, Tang WJ, Stein RL, Cuny GD, Selkoe DJ (2010). Designed inhibitors of insulin-degrading enzyme regulate the catabolism and activity of insulin. PLoS ONE 5(5): e10504. |
|
|
Li CZ, Zhang SH, Shu CD, Ren W (2002). [Relationship between insulin-degrading enzyme activity and insulin sensitivity in cell model of insulin-resistance]. Di 1 jun yi da xue xue bao= Academic journal of the first medical college of PLA 22(2):151-154. |
|
|
Maestro B, Campión J, Dávila N, Calle C (2000). Stimulation by 1,25-dihydroxyvitamin D3 of insulin receptor expression and insulin responsiveness for glucose transport in U-937 human promonocytic cells. Endocr. J. 47(4):383-91. |
|
|
Maianti JP, McFedries A, Foda ZH, Kleiner RE, Du XQ, Leissring MA, Liu, DR (2014). Anti-diabetic activity of insulin-degrading enzyme inhibitors mediated by multiple hormones. Nature 511(7507): 94-98. |
|
|
Mamikutty N, Thent ZC, Sapri SR, Sahruddin NN, MohdYusof MR, Haji SF (2014). The establishment of metabolic syndrome model by induction of fructose drinking water in male Wistar rats. Biomed. Res. Int. 2014:263897. |
|
|
Matveyenko AV, Veldhuis JD, Butler PC (2008). Adaptations in pulsatile insulin secretion, hepatic insulin clearance, and β-cell mass to age-related insulin resistance in rats. Am. J. Physiol. Endocrinol. Metab. 295(4):E832-E841. |
|
|
McGarry JD (2001). Dysregulation of fatty acid metabolism in the etiology of type 2 diabetes. Diabetes. 51(1):7-18. |
|
|
Mittelman SD, Van Citters GW, Kim SP, Davis DA, Dea MK, Hamilton-Wessler M, Bergman RN. (2000). Longitudinal compensation for fat-induced insulin resistance includes reduced insulin clearance and enhanced β-cell response. Diabetes 49(12):2116-2125. |
|
|
Moller DE (2001). New drug targets for type 2 diabetes and the metabolic syndrome. Nature 414(6865):821-827. |
|
|
Paschos P, Paletas K (2009). Non alcoholic fatty liver disease and metabolic syndrome. Hippokratia 13(1):9-19. |
|
|
Perna AF, Fadda GZ, Zhou XJ, Massry SG (1990). Mechanisms of impaired insulin secretion after chronic excess of parathyroid hormone. Am. J. Physiol. 259(2 Pt 2):F210-6. |
|
|
Pessin JE, Thurmond DC, Elmendorf JS, Coker KJ, Okada S (1999). Molecular basis of insulin-stimulated GLUT4 vesicle trafficking. J. Biol. Chem. 274(5):2593-2596. |
|
|
Rezende LF, Santos GJ, Santos-Silva JC, Carneiro EM, Boschero AC (2012). Ciliaryneurotrophic factor (CNTF) protects non-obese Swiss mice against type 2 diabetes by increasing β cell mass and reducing insulin clearance. Diabetologia 55(5):1495-504. |
|
|
SlominskiÄ PA, Pivovarova OV, Shadrina MI, Artem'eva AV, Pfaipffer FG, Rudovich NN, Agadzhanian SE, Pronin VS, Limborskaia SA (2009). Association of insulinase gene polymorphisms with type 2 diabetes mellitus in patients from the Moscow population. Genetika 45(1):127-131. |
|
|
Sung CC, Liao MT, Lu KC, Wu CC (2012). Role of vitamin D in insulin resistance. J. Biomed. Biotechnol. 2012. |
|
|
Teegarden D, Donkin SS (2009). Vitamin D: emerging new roles in insulin sensitivity. Nutr. Res. Rev. 22(1):82-92. |
|
|
Wei X, Bilun Ke, Zhiyun Zhao, Xin Ye, Zhanguo Gao, Jianping Ye (2014). Regulation of insulin degrading enzyme activity by obesity-associated factors and pioglitazone in liver of diet-induced obese mice. PLoS One 9(4):e95399. |
|
|
Williams PF, Caterson ID, Cooney GJ, Zilkens RR, Turtle JR (1990). High affinity insulin binding and insulin receptor-effector coupling: modulation by Ca2+. Cell Calcium 11(8):547-556. |
|
|
Yoshii H, Lam TK, Gupta N, Goh T, Haber CA, Uchino H, Kim TT, Chong VZ, Shah K, Fantus IG, Mari A, Kawamori R, Giacca A (2006). Effects of portal free fatty acid elevation on insulin clearance and hepatic glucose flux. Am. J. Physiol. Endocrinol. Metab. 290(6):E1089-E1097. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


