Full Length Research Paper

ABSTRACT
The potency and competitive ability of indigenous rhizobia population in soil to nodulate a non-native legume host has been contentious especially in tropical Africa. This study examined the symbiotic compatibility between the indigenous rhizobia population and a non-native legume species. Soil samples were randomly collected with soil auger at 0 - 30 cm depth from agricultural fields southeast, Nigeria. The control soil samples were obtained from the Badagry beach which had no previous history of legume cultivation at the same depth. Mucuna pruriens seeds were aseptically sown in plastic pots containing these soils kept in the greenhouse for 10 weeks. The test pots were watered with sterile distilled water while the control pots received inorganic nitrogen (N) as source of N nutrient. The agronomic features of the host legume were evaluated after 10 weeks and the presence of appropriate Rhizobium inoculant contributed positively to biomass production in M. pruriens. The Mucuna microsymbiont was found to thrive under favorable climatic and edaphic factors for effective nodulation of the host legume. The microsymbiont was identified on yeast-extract mannitol salt agar (YEMA) containing bromothymol blue (BTB) as Bradyrhizobium and Rhizobium species. This study convincingly showed the importance of rhizobia to soil fertility maintenance in sustainable agricultural practice as well as the adaptability of the legume microsymbiont to different ecological zones particularly the tropical humid environment.
Key words: Inoculation, Mucuna pruriens, nodulation, organic agriculture, Rhizobium, soil fertility.
INTRODUCTION
The challenge facing agriculture in sub-Saharan Africa is decreasing soil fertility and decreasing productivity. This is due to excessive cultivation of farmlands which results in low fertility in most of the soils in sub-Saharan Africa and the limited use of fertilizers which often are not readily available (Anonymous, 1999).
Leguminous cover crops such as in alley cropping have the potential to contribute to soil nitrogen and increase the yields of subsequent or associated cereal crops through symbiotic N2-fixation. Although, it has often been wrong-fully assumed that Mucuna pruriens will freely nodulate, fix N2 and therefore contribute to soil N. Recent findings suggested M. pruriens failure to nodulate in some farmer’s field in the derived Savanna in Republic of Benin. Although, Mucuna could play an important role as a source of N, it may also depend solely on soil N and therefore may deplete soil N, hence behaving as non-fixers when either not effectively nodulated or there is too low nutrient availability particularly phosphorous (P) (Hougnandan et al., 2000; Jemo et al., 2007).
M. pruriens has been reported to be adequately established and well nodulated in soil in the presence of its microsymbiont, favorable climatic and edaphic factors which enhances its establishment in soil within the humid and moist savanna zone of Nigeria (Ojo, 2001; Jemo et al., 2007).
The severity of the spear grass (Imperata cylindrical) problem, the decline of the traditional fallow system as a means of soil fertility management and lack of inorganic fertilizer, necessitated the adoption of M. pruriens technology by small holders in some areas in the derived Savanna of West Africa (Akobundu and Okigbo, 1984; Versteeg and Koudokpon, 1993; Sanginga et al., 1996). In West Africa, crop productivity declines rapidly when the fallow period is shortened below a minimal period of time needed to restore soil fertility to a level accepted for sustainable agricultural practice. Hence, soils are degrading rapidly and nutrient balances are negative (Van der Pol et al., 1993). In order to tackle this problem in West Africa, the development of cropping systems that enhances sustainable crop production at an economically accepted level with minimal external inputs has long been proposed. Alley cropping systems (Kang et al., 1981) and other soil fertility regenerating techniques such as intercropping of maize and Pigeon pea (Cajanus cajan), relay planting of M. pruriens one month after planting maize or planting tree fallows of Acacia auriculiformis on severely degraded soils have since been tested in the Mono province of Benin Republic by Versteeg and Koudokpon (1993). In alley cropping systems, trees are intercropped with crops and cut back regularly to avoid tree-crop competition for water, light or nutrients. The tree pruning is subsequently applied to soil surface thereby supplying nutrients. Although, alley cropping systems have proven to sustain crop productivity on soils of high base saturation with minimal supplies of mineral fertilizer (Kang et al., 1995), the use of M. pruriens as green manure for intensive maize (Zea mays L.) production in West Africa has given much positive yield effect to suggest its integration into sustainable agriculture practice within the region (Hauser and Nolte, 2002). The socio-economic conditions of the peasant farmers have prevented the absolute adoption of this system as well as reports that N2 released from decomposing residues is poorly recovered by food crops (Vanlauwe et al., 1996b, 1998).
The selection of adequately adapted inoculants strains and understanding of changes in rhizobia population as well as the factors precipitating such changes would enhance the application of appropriate indigenous rhizobial strains in the desired low-input farming system and subsequently enhance both M. pruriens biomass and N uptake (Sanginga et al., 1994; Jemo et al., 2007). The challenge of decreasing agricultural productivity in tropical Africa would be eradicated by synchronization of low-input systems such as Rhizobium/legume association into farming systems under favorable climatic and edaphic factors, which are not only sustainable but also economically viable (Anonymous, 1991). The potency and competitive ability of introduced rhizobia inoculants in soil for over 10 years have been reported (Ojo and Fagade, 2002). However, in many soils adequate nodulation and N2-fixation by legumes require inoculation with the appropriate and specific rhizobia inoculants particularly where such legumes has not been previously cultivated (Sanginga et al., 1988; Jemo et al., 2007).
The objectives of the current study were to assess the ability of promiscuous native rhizobia population in a humid tropical environment to adequately nodulate M. pruriens as well as their potential to produce biomass in spite of the prevailing climatic and edaphic factors in the environment under investigation.
MATERIALS AND METHODS
Soils
Soil samples were randomly collected from agricultural field in Aba, Calabar, Enugu, Onitsha, Lokoja, Owerri and Benin within the tropical humid zone, Southeast of Nigeria at a depth of 0-30 cm using a soil auger (5 cm diameter). Ten core soil samples were randomly collected with the soil auger from each of the agricultural fields. Soil samples with no previous history of being inoculated with Mucuna microsymbiont as well as not having been cultivated with legumes were collected at same depth from another location (Badagry beach) to serve as the control. The composite soil samples from each of the experimental fields sampled in each of the ecological zones were then used for the pot experiment in the green house using the randomized complete block design.
Seeds
Legume seeds were obtained from the Genetic Resources Unit (GRU), International Institute of Tropical Agriculture (IITA) Ibadan, Nigeria.
Media and plant nutrients
Yeast-extract mannitol agar (YEMA), Jensen’s Nutrient solution and potassium nitrate (KNO3) solution at 0.05% N were used (Vincent, 1970).
Soil analysis procedures
Preparation of the soil samples
The composite soil sample from each location was passed through a 6-mm mesh sieve and mix thoroughly (Ocio and Brookes, 1990b). Then, three portions of the soil were weighed out, 15-50 g, one portion into a weighing container for water content determination and two portions into 100 ml glass bottles, one sample to be fumigated for 24 h and then extracted as well as one control sample to be extracted immediately.
Extraction of microbial biomass C and N
The fumigation was carried out using CHCl3. The bottles containing the unfumigated control and fumigated subsamples had 0.5 M K2SO4 dispensed into them using the equivalent oven-dry soil weigh (g): extractant volume (ml) ratio 1:2 to 1:5. The jars with their caps tightened were placed on a rotatory shaker for 1 h. After shaking, the soil suspension was pass through the Whatman No.5 filter paper; excessive evaporation was avoided during the filtration process. Thereafter, measurements of organic C and total N were done using 20 ml of extracts in each case. Then, the filtrate was capped and stored at 4°C for not more than 2 -3 days (Voroney et al., 1993).
Determination of organic C, organic N and exchangeable NH4+ in the extract
Organic C, organic N and exchangeable NH4+ dissolved in the K2SO4 extracts were determined colorimetrically using automated equipment as described by Voroney et al. (1993).
Nodulation experiment
The composite soil samples were tested with scarified M. pruriens seeds to evaluate their potential at nodulating with promiscuous native rhizobia population in the soils. Seven plastic pots, 20 mm diameter were used to cultivate the legume and replicated twice. Two plastic pots of the same diameter were used for the control experiment and this was replicated twice. A total of sixteen plastic pots were used for the experiment, randomly arranged on a platform in the greenhouse. Four scarified M. pruriens seeds were sown aseptically into each pot and later thinned to two plants per pot with sterile forceps 2 weeks after planting (WAP). Each of the pot for the test experiment was watered with sterile distilled water while one pair of the control pot received Jensen’s nutrient solution and the other pair 0.05% KNO3 solution (Vincent, 1970).
The experiment was monitored for 10 weeks. The above-ground plant materials were harvested with sterile surgical blades and then oven-dried at 65°C for 48 h. Roots were carefully removed and examined for nodulation. Fresh nodules were counted, cleaned of soil particles and then used for strain identification (Sanginga et al., 1994).
Nodule-typing on YEMA
Aseptic procedures were practiced in typing the harvested nodules on YEMA containing bromothymol blue (BTB). The nodules were crushed with sterile glass rods in sterile Petri dishes and then inoculated on YEMA containing BTB (0.0005%) incubated at 28°C for 48 h. Pure cultures were prepared from the isolates and used for plant re-infection experiment as well as for biochemical characterization (Somasegaran and Hoben, 1994; Ojo, 2001).
Morphological and biochemical tests
The morphological and biochemical characterization of rhizobia from harvested nodules of M. pruriens were executed following the methods described by Somasegaran and Hoben (1994) and Gerhardt et al. (1981).
Gram reaction
A thin film of each of the isolate was prepared on a grease-free slide and it was air-dried as well as heat-fixed by passing it horizontally over the Bunsen flame. The smear was stained with crystal violet for 60 s and rinsed off under slow running tap water. The resulting smear was thereafter stained with Gram’s iodine and left for 30 s. This was rinsed off with tap water. The decolorizing agent (75% Ethanol) was used to flush the smear on the slide and was immediately rinsed with water. Thereafter, few drops of the counter stain Safranine was applied on the smear for 30 s and then rinsed with tap water. The slide was left to dry and later examined under oil immersion lens (100x) of the microscope. Gram positive organisms appear purple in color while Gram negative organisms appear pink or red under the microscope (Gerhardt et al., 1981).
Catalase test
This was perfected by adding 3% hydrogen peroxide to 18-24 h culture on agar slant or glass slide. The culture was observed for immediate appearance of bubbles (Alexander and Strete, 2001). Presence of bubbles is considered a positive test.
Oxidase test
This was performed by adding oxidase reagent (tetramethyl–p–phenylenediamine dihydrochloride) to a bacterial growth on agar plate. Color change was observed for up to 60 s (Alexander and Strete, 2001). A dark purple color represents a positive test.
Indole test
Suspected isolates were inoculated into test-tubes containing peptone water with a sterile transfer needle. Test-tubes were incubated at 28±2°C for 24 - 48 h. Thereafter, five drops of Kovac’s reagent (p-dimethylaminobenzaldehyde) was added to the medium (Alexander and Strete, 2001). A red color on medium represents a positive test for indole production.
Growth on YEMA supplemented with BTB
The growth reactions in the standard yeast-extract medium (YM) containing bromthymol blue (BTB) acted as pH indicator. Fast-growing rhizobia produce an acid reaction in the YM medium (yellow color) containing BTB (pH 6.8) while slow-growing rhizobia produce an alkaline reaction (blue or purple color) (Somasegaran and Hoben, 1994).
Motility test
This was used to detect the presence of flagella. Suspected isolates were inoculated into nutrient agar slant with a sterile transfer needle inserted and withdrawn in a straight line in the centre. The inoculated slants were incubated at 28±2°C for 24 - 48 h (Alexander and Strete, 2001). Growth away from line of inoculation indicates the organism is motile.
Statistics
Statistical Package for Social Sciences (SPSS) software model (Version 16) was used for the statistical analysis and analysis of variance (ANOVA).
RESULTS
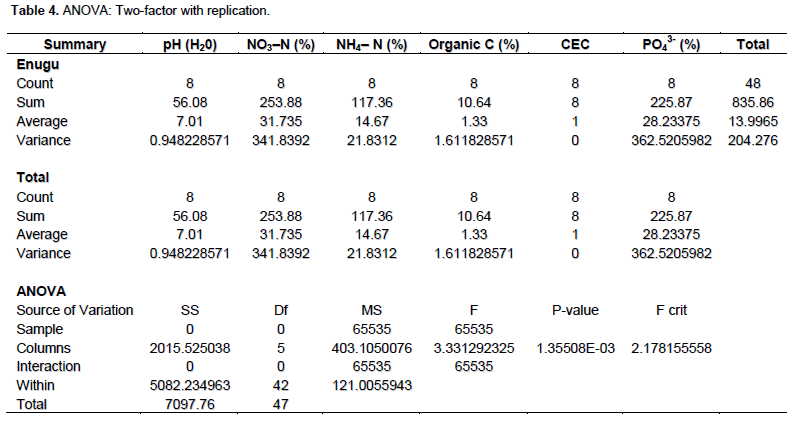
The mean physico-chemical properties of the soil samples showed that all the locations had some level of fertility that could support sustainable agricultural practice. The soil samples from Benin, Enugu, Owerri and Onitsha were slightly acidic while other locations were slightly alkaline (Table 1). The general presentation of soil pH from the southeast ecological zone was neutral (7.01) (P> 0.05) as shown in Table 4. The mean nitrogen (N) content showed that the soil had more NO3–N than NH4–N (Tables 1 and 4). Generally, with the quantity of soil N in the soil, it is expected that N2- fixation should complement the soil N to facilitate the establishment of the legume host.
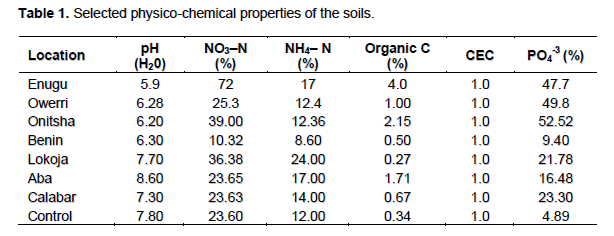
The mean PO43- nutrient was low (28.2%) (P>0.05) but this is adequate for M. pruriens establishment in the soil (Tables 1 and 4). Phosphorus (P) and nitrogen are essential mineral nutrient for the survival of rhizobia population in soil which had been provided by the soil samples. The mean organic carbon (C) content (1.33%) though extremely low, the CEC property of the soils were expected to support the proliferation of M. pruriens microsymbiont as well as cation exchange in the soil (Tables 1 and 4). The control soil samples had mineral nutrient in them but they were sterile soils hence no M. pruriens microsymbiont in them. The rhizobia count using yeast-extract mannitol salt agar (YEMA) was above 300 cells/g of soil at the commencement of this study from each location. The mean agronomic features of M. pruriens var cochinensis 10 weeks after planting (WAP) showed that the legume was established in all soil samples. The moisture accumulation was relatively high in all pots, this was as a result of quantity of watering process applied in the greenhouse and the experiment was conducted during rainy season. It was obvious that nodulation failed with soil samples from Onitsha despite the firm establishment of the legume host and its biomass production. The nodule number (#) was highest with Enugu, Calabar, Benin, Owerri, Lokoja and Aba in a descending order.
Root and shoot dry weights which are pointers to the quality of biomass production were highest in Calabar and Lokoja, respectively (Table 2). The mean root and shoot dry weight were 1.27 and 3.61 g, respectively while mean nodule number (6) suggested poor nodulation 10 WAP (P> 0.05) as shown in Table 5.
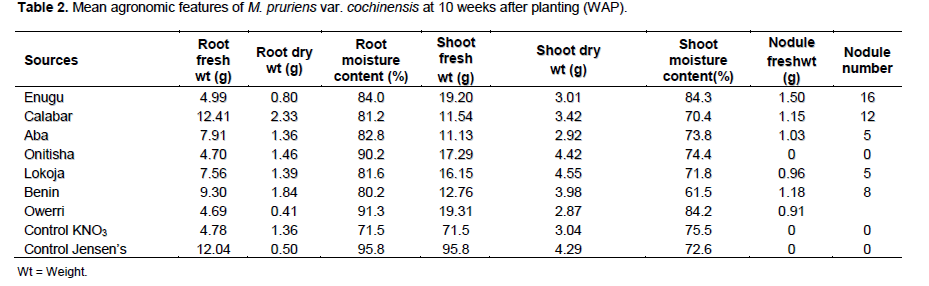
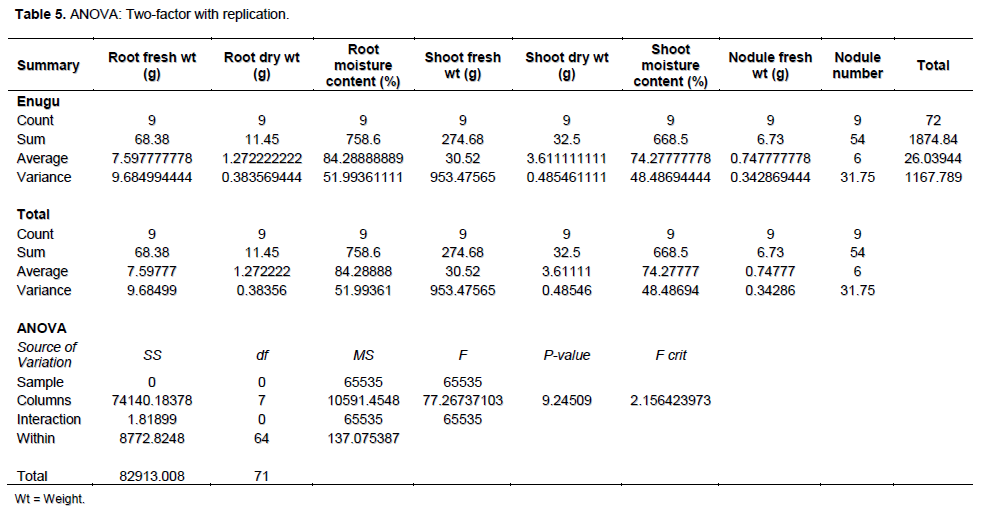
Generally, nodulation of M. pruriens was relatively poor in all locations 10 WAP because only few of the indigenous rhizobia strains were specific for this M. pruriens species which is of Asian origin.
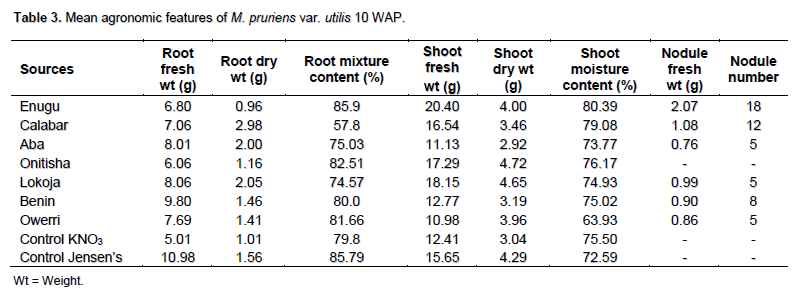
The mean agronomic features of M. pruriens var utilis showed firm establishment of the legume host but it failed to nodulate with native rhizobia species specifically in Onitsha soil samples (Table 3). Biomass production 10 WAP in terms of root and shoot dry weight was highest in Calabar and Onitsha, respectively. The mean root and shoot dry weight were 1.62 and 3.80 g, respectively with mean nodule number 5.88 (P> 0.05) as shown in Table 6. In both varieties of legume used, the uninoculated controls were not comparable to the inoculated pots in terms of biomass production. There was no nodulation in the control pots since they were uninoculated. The presence of Rhizobium species in the legume roots could only have been the factor that enhanced the biomass production in most of the inoculated pots. Furthermore, the contents of the nodules aseptically inoculated on YEMA containing BTB showed yellow (acidic) and blue (alkaline) coloration which suggested the presence of Rhizobium and Bradyrhizobium species, respectively. The results of this experiment validates the null hypothesis (H0); M. pruriens microsymbiont are quite scanty in population southeast, Nigeria while the native rhizobia population are incompatible and non-specific for M. pruriens varieties cultivated. The alternative hypothesis (H1); the native rhizobia population in soils southeast, Nigeria is compatible and specific for the introduced M. pruriens varieties (Table 7).
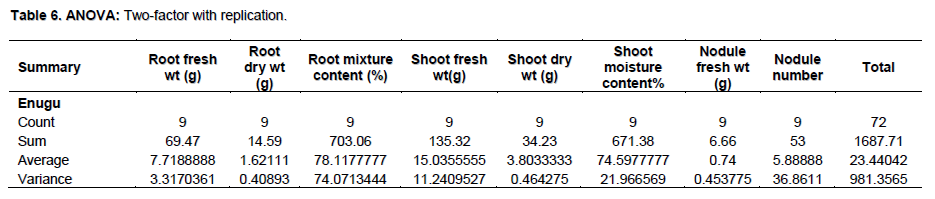
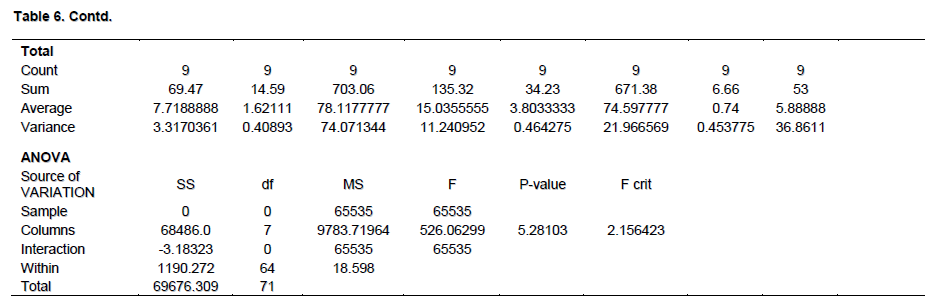

Therefore, statistical analysis results showed that native rhizobia species specific for nodulation of M. pruriens in southeast Nigeria are scanty, however, the legume host was well established.
DISCUSSION
The root-nodule bacteria have been studied for long but little of its potential contribution to organic farming as well as sustainable agriculture has been explored. The detection of nodules showed presence of indigenous promiscuous M. pruriens microsymbiont with the competitive edge over other rhizobia strains in the soil hence its ability to nodulate M. pruriens. The absence of nodulation of M. pruriens in control pots showed the effectiveness of sterilization procedures applied to control soils as well as the fact that rhizobia species specific for M. pruriens were absent in the control pots.
M. pruriens varieties planted were well established in all the ecological zones. The two varieties (M. pruriens var. utilis and M. pruriens var. cochinensis) failed to nodulate with native rhizobia population in soil samples from Onitsha, Southeast Nigeria. Soils from Onitsha, Owerri, Benin and Enugu were acidic (Table 1). Compara-tively, Onitsha soil was the best in terms of root moisture retention while in terms of shoot biomass production, Lokoja soil was the best. The percentage (%) moisture in the root/shoot showed that more water absorption occurred in the soils from the core east (Enugu, Aba, Onitsha and Owerri) due to the soil texture (Table 2). The presence of rhizobia species as biological N fertilizer contributed to the improvement of soil fertility thus enhancing the establishment of M. pruriens in the soil environment southeast, Nigeria; although, soil acidity has been reported to adversely affect agricultural production and limits legume productivity (Bordeleau and Prevost, 1994; Correa and Barneix, 1997). This observation suggested that certain environmental stress may be responsible for the absence of nodulation in Onitsha soil. In this case, indigenous strains were unable to adapt to acid stress and probably some other environmental stress not investigated. While this same soil samples had relatively adequate N and P nutrient supply as well as organic C, these findings corroborated earlier report by Walsh (1995) and Zahran (1999).
Although, at the commencement of this study, the mean indigenous rhizobia population was found to be above 300 cells/g of soil for these locations, the presence of these rhizobia species was confirmed using standard and conventional methods as reported by Vincent (1970).
Typical environmental stress earlier discovered by previous researcher include photosynthetic deprivation, water stress, poor water-holding capacity (Bottomley, 1991), salinity, soil nitrate, temperature, presence of heavy metals, acidic soil and biocides to militate against the legume-microsymbiont systems in soil which further corroborated the deductions of Walsh (1995). More so, the population of Rhizobium and Bradyrhizobium species vary in their level of tolerance to adverse environmental changes. However, Enugu soil which is the most acidic (Table 1) had the highest nodulation. One physico-chemical factor that was outstanding with this location was the relatively high NO3–N and PO4-3 nutrient in this location. The tolerance of the M. pruriens microsymbiont (Rhizobium sp.) to the prevailing climatic and edaphic factors might have enhanced the success of the symbiotic association in Enugu soil as well as its water-holding capacity (Table 2). Screening for the tolerant strains of M. pruriens microsymbiont indigenous to Onitsha and well adapted to its climatic and edaphic conditions failed because nodulation did not take place at this location. Poor nodulation is also possible in soils where a viable rhizobia population can be demonstrated (Graham et al., 1994). However, M. pruriens microsymbiont (Bradyrhizobium sp.) isolated from Lokoja, Aba and Calabar soils were influenced by the alkaline soil pH. The pH of the rhizosphere thus deter-mines the type microsymbiont that occupies the nodules of the host legume.
High soil temperature in tropical and sub-tropical areas has been identified to be inhibitory or delay nodulation (Graham, 1992), particularly in creeping legumes nodulation which is affected by high tropical temperatures which may be as high as between 28 and 35°C in the January – April period. The optimum temperature for effective nodulation is between 25 and 30°C (Michiels et al., 1994; Arayankoon et al., 1990). Furthermore, the presence of heavy metals contaminants on cultivated plots as a result of the use of sewage sludge as organic fertilizer which often contains materials potentially toxic to soil microorganisms have been reported to have adverse effects on nodulation and N2-fixation in legumes (McGrath et al., 1988). Other chemical contaminants such as polycyclic aromatic hydrocarbons (PAHs) which occur as ubiquitous environmental contaminants due to the combustion of fossil fuels can affect nodulation and N2–fixation in legumes (Wetzel and Werner, 1995). The presence of rhizobia as N fertilizer contributed to the improvement of soil fertility thus enhancing the establishment of M. pruriens in the soil environment southeast, Nigeria.
The nodule number profile (Table 2) suggested the relative presence of more M. pruriens microsymbiont in the soil from Enugu than in all other locations. Considering both the root and shoot dry weight (Table 2) which suggested the degree of soil fertility available to the legume particularly in collaboration with the microsymbiont, the agronomic parameters of M. pruriens var cochinensis showed that soils from Benin, Onitsha and Lokoja relatively supported biomass production more than in other locations. Although, nodulation failure was observed on Onitsha soil, this calls for further investigation of probable causes of this apart from climatic and edaphic reasons. Whereas M. pruriens var utilis (Table 3) showed excellent agronomic features with soils from Calabar, Lokoja and Aba in terms of root dry weight. However, considering shoot dry weight, soil from Onitsha, Lokoja, Enugu and Owerri performed excellently (Table 3). This performance can be attributed to the level of fertility of these soils. These findings are in agreement with the conclusion of Alhou et al. (1999) and Jemo et al. (2007).
The statistical analysis was critical with ANOVA using two-factor replication. Some ecological zones in this study have compatible and specific M. pruriens microsymbiont which have adapted to the introduced Mucuna varieties.
CONCLUSION
Biological nitrogen fixation (BNF) is an efficient source of N for increasing agricultural productivity. However, its success is inhibited by various adverse environmental conditions which subsequently affect crop yields. The M. pruriens seeds used in this study were of Asian origin but were perfectly adapted to the tropical humid climate in this study. The use of rhizobia as inoculants could be a successful biological process capable of improving crop yield while simultaneously providing biomass for other industry use.
The Rhizobium-legume symbiosis is hereby suggested as an ideal solution to the improvement of soil fertility as well as the rehabilitation of adversely degraded soil and this is worthy of priority for future research.
Natural selection of stress-tolerant rhizobia strains would be a non-polluting and cost-effective way to improve soil fertility as compared to other common ways such as inorganic NPK fertilizer and sewage sludge application. This will improve international trade volume because the crop yield from this type of agricultural practice would have conformed to the specifications of the federation of international organic agriculture standards.
CONFLICT OF INTEREST
The authors did not declare any conflict of interest.
ACKNOWLEDGEMENT
The authors gratefully appreciate Oluremi Adeyemo, Health Services Unit (LASU) for her assistance in the preparation of the manuscript.
REFERENCES
|
Akobundu IO, Okigbo BN (1984). Preliminary evaluation of ground covers for use as live-mulch in maize production. Field Crops Res. 8:177-186. Crossref |
||||
| Alexander SK, Strete D (2001). Microbiology In: a photographic atlas for the Laboratory (B. Cummings Ed.) An imprint of Addison Wesley Longman Inc. pp. 69-92. | ||||
| Alhou K, Sanginga N, Vanlauwe B, Lyasse O, Diels J, Merckx R (1999). Alley cropping in the moist savanna of West Africa / Restoration and Maintenance of soil fetility on 'terre de barre' soils in Benin Republic. Agric. For. Syst. 42:213-227. | ||||
| Anonymous (1991). Harnessing BNF for sustainable agriculture IITA Res. 12:25-26. | ||||
| Anonymous (1999). Agricultural research and Development in Sub-Saharan Africa In : Development of sustainable Agriculture in Sub-Saharan Africa. IITA Report pp. 13-20. | ||||
|
Arayankoon T, Schomberg HH, Weaver RW (1990). Nodulation and N2-fixation of guar at high root temperature. Plant Soil 126:209-213. Crossref |
||||
| Bottomley P (1991). Ecology of Rhizobium and Bradyrhizobium In: Stacey et al. (eds) Biological nitrogen fixation. Chapman & Hall, New York, N.Y. pp. 292-347. | ||||
|
Bordeleau LM, Prevost D (1994). Nodulation and Nitrogen fixation in extreme environments. Plant Soil 16:115-124. Crossref |
||||
|
Correa OS, Barneix AJ (1997). Cellular Mechanisms of pH tolerance in Rhizobium loti. World J. Microbiol. Biotech. 13:153-157. Crossref |
||||
| Gerhardt P, Murray RGE, Costilow RN, Nester EW, Wood WA, Krieg NR, Phillips GB (1981). Manual of Methods for general Bacteriology. Am. Soc. Microbiol. pp. 413- 419. | ||||
|
Graham PH (1992). Stress tolerance in Rhizobium and Bradyrhizobium, and nodulation under adverse soil conditions. Can. J. Microbiol. 38:475-484. Crossref |
||||
|
Graham PH, Draeger K, Ferrey ML, Conroy MJ, Hammer BE, Martinez E, Aarons SR, Quint C (1994). Acid pH tolerance in strains of Rhizobium and Bradyrhizobium and initial studies on the basis for acid tolerance of Rhizobium tropicalis UMR 1899. Can. J. Microbiol. 40: 198-207. Crossref |
||||
| Hougnandan P, Vanlauwe B, Diels J (2000). Participative farm trials with M. pruriens. Soil Biol. Biochem. 34:512-516. | ||||
|
Hauser S, Nolte C (2002). Biomass production and N fixation of five Mucuna pruriens varieties and their effect on maize yields in the forest zone of Cameroon. J. Plant Nutr. Soil Sci. 165(1):101-109. Crossref |
||||
| Jemo M, Nolte C, Nwaga D (2007). Biomass production, N and P uptake of Mucuna after Bradyrhizobia and Arbuscular mycorrhizal fungi inoculation and P- application on acid soil of southern Cameroon In: Advances in integrated soil fertility management in sub- Saharan Africa: challenges and opportunities pp. 855- 864. | ||||
| Kang BT, Hauser S, Vanlauwe B, Sanginga N, Atta-Krah AN (1995). Alley farming research on high base status soils. In: Kang et al. (eds.) Alley Farming Research and Development IITA. Nigeria, pp. 25-39. | ||||
|
Kang BT, Wilson GF, Sipkens L (1981). Alley cropping Maize (Zea mays L.) and Leucaena (Leucaena leucocephala) in Southern Nigeria. Plant Soil 63:165-179. Crossref |
||||
|
McGrath SP, Brooks PC, Giller KE (1988). Effects of potentially toxic metals in soil derived from past applications of sewage sludge on nitrogen fixation by Trifolium repens L. Soil Biol. Biochem. 20:415- 424. Crossref |
||||
|
Michiels J, Verreth C, Vander leyden J (1994). Effects of temperature stress on bean nodulating Rhizobium strains. Appl. Environ. Microbiol. 60:1206-1212. Pubmed |
||||
|
Ocio JA, Brookes PC (1990b). Soil microbial biomass measurements in sieved and unsieved soil. Soil Biol. Biochem. 22:999-1000. Crossref |
||||
|
Ojo OA (2001). Assessment of nodulation of Mucuna pruriens by promiscuous indigenous rhizobia in the moist savanna zone of Nigeria. World J. Microbiol. Biotech. 17(4):429-431. Crossref |
||||
|
Ojo OA, Fagade OE (2002). Persistence of Rhizobium inoculants originating from Leucaena leucocephala fallowed plots in southwest Nigeria. Afr. J. Biotechnol. 1(1):2327. |
||||
|
Sanginga N, Danso SKA, Mulongoy K, Ojeifo AA (1994). Persistence and recovery of introduced Rhizobium ten years after inoculation on L. leucocephala grown on an Alfisol in Southwestern Nigeria. Plant Soil 159:199-204. Crossref |
||||
|
Sanginga N, Mulongoy K, Ayanba A (1988). Nitrogen contribution of Leucaena/ Rhizobium symbiosis to soil and a subsequent maize crop. Plant Soil 112:137-141. Crossref |
||||
|
Sanginga N, Ibewiro B, Hougnandan P, Vanlauwe B, Okogun JA, Akobundu IO, Versteeg M (1996). Evaluation of symbiotic properties and nitrogen contribution of Mucuna to maize grown in the derivedSavanna of West Africa. Plant Soil, 179:119-129. Crossref |
||||
|
Somasegaran P, Hoben HJ (1994). Handbook for Rhizobia. In: Methods in Legume-Rhizobium Technology, New York Springer-Verlag. pp 450. ISBN 0-38794134-7. Statistical Package for Social Sciences (SPSS) version 16.0. Crossref |
||||
| Van der Pol F, Gogan AC, Dagbenombakin G (1993). L'Epuisement des sols et sa Valeur Economique dans le Department du Mono Benin. RAMAR. Cotonou, Benin. 48pp. | ||||
|
Vanlauwe B, Swift MJ, Merckx R (1996b). Soil litter dynamics and N use in a Leucaena (Leucaena leucocephala (Lam. de Wit.) alley cropping system in Southwestern Nigeria. Soil Biol. Biochem. 28:739-749. Crossref |
||||
|
Vanlauwe B, Diels J, Duchateau L, Sanginga N, Merckx R (1998). Mineral N dynamics in bare and cropped Leucaena leucocephala and Dactyladenia barteri alley cropping systems after the addition of 15N – labeled leaf residues. Eur. J. Biol. Sci. 49:417-425. Crossref |
||||
|
Versteeg MN, Koudokpon V (1993). Participatory farmer testing of four low external input technologies to address soil fertility decline in Mono Province (Benin). Agric. Syst. 42:265-276. Crossref |
||||
| Vincent JM (1970). A Manual for the Practical Study of Root Nodule Bacteria. IBP Handbook No. 15 pp. Oxford; Blackwell. ISBN 0- 63206410-2. | ||||
| Voroney RP, Winter JP, Beyaert RP (1993). Soil microbial biomass C and N In: Soil sampling and methods of analysis. (Ed.) Martin R. Carter. Can. Soc. Soil Sci. pp. 277-282. | ||||
|
Walsh KB (1995). Physiology of the legume nodule and its response to stress. Soil Biol /Biochem. 27:637-655. Crossref |
||||
|
Wetzel A, Werner D (1995). Ecotoxicological evaluation of contaminated soil using the legume root nodule symbiosis as effect parameter. Environ. Toxicol. Water Qual. 10:127-134. Crossref |
||||
| Zahran HH (1999). Rhizobium-legume symbiosis and Nitrogen fixation under severe conditions and in an arid climate. Mol. Biol. Rev. 63(4): 968-989. | ||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


