Full Length Research Paper

ABSTRACT
Leaves of nine species of Ludwigia were collected from Southwestern Nigeria and the crude proteins were extracted and analysed by electrophoretic fractionation. The results shows that Ludwigia octovalvis var linearis, Ludwigia octovalvis var brevisepala, Ludwigia hyssopifolia and Ludwigia abyssinica are more closely related based on the position of protein bands. Also, Ludwigia decurrens A and Ludwigia adscendens subsp. diffusa are more closely related based on the number of band and the position of band. The band at 2.1 is taxonomic for all the species of Ludwigia studied. Similarly, the band at 1.0 delimits L. leptocarpa from the other species. Protein abundance sequence of the samples is in the order L. hyssopifolia ˃ L. octovalvis var linearis ˃ L. abyssinica ˃ L. octovalvis var brevisepala ˃ L. erecta ˃ L. decurrens B ˃ L. leptocarpa ˃ L. decurrens A ˃ L. adscendens subsp. diffusa. The occurrence of a new band in Ludwigia decurrence B separates it from Ludwigia decurrence A, a new hypothetical name is suggested for the delimitation of the two species. The protein bands are taxonomically distinct as no two species have the same band distribution; diagnostic bands which could be employed for the identification of each species are also reported.
Key words: Ludwigia, protein bands distribution, taxonomic implication, Nigeria.
INTRODUCTION
Ludwigia species have been classified among the 200 most aggressive world plant invaders (Cronk and Fuller, 1995). This is a genus of considerable economic importance; Ludwigia octovalvis as a traditional herbal medicine, has been used to treat gastrointestinal disorders such as diarrhoea and dysentery, a poultice of an entire plant is externally applied to heal dermatitis, boil, ulcer, impetigo and pimple (Kadum et al., 2012). The leaves of L. abyssinica A. Rich. are edible and used for dyeing of straw and fibres, and it is used in medicine applied for enhancing memory. The leaves of L. erecta (L.) H. Hara are edible as vegetables and used for treating fevers. Ludwigia hyssopifolia (G. Don) Exell is valuable as green manure and the leaves can be used to treat wounds (Kadiri and Olowokudejo, 2010).
Fluctuations in the taxonomic classifications of Ludwigia species have been reported (Dutartre et al., 2004). These have been attributed to their phenotypic plasticity; in other words, their growth forms vary under different environmental conditions which often complicate species identification. Most classification works on the genus has been hinged on evidences from leaf morphology, palynology, and flower and seed morphology. Similarly, Folorunso et al. (2014) reported the use of foliar and stem anatomical characters in the identification of Ludwigia species in Nigeria.
Electrophoretic techniques for identification and classification have become a very important tool in systematic research. This technique is particularly of taxonomic importance in separating varieties of plants (Folorunso et al., 2012) and also a useful tool in studies of genetic variability in plants (Oladipo et al., 2008). Electrophoretic technique has been employed on a number of plant groups to show that many isoenzymes or polymorphic proteins are widely distributed in higher plants and also to compare protein distribution of the wild relative of plants to the cultivated ones (Folorunso et al., 2012; Oladipo and Illoh, 2012).
This study therefore compares the electrophoretic pattern of protein bands distribution in the Ludwigia species with the aim of providing useful information on classification and identification of Ludwigia species and identifies both intraspecific and interspecific variations that exist among them and the highest protein richness in them.
MATERIALS AND METHODS
Fresh leaves of nine (9) species of Ludwigia common to Nigeria in the family Onagraceae were collected from the Southwestern Nigeria (Table 1). The altitudes and geographical coordinates of the localities were taken using a GPS device. Species identification and confirmation was done with the assistance of the curators at the Herbarium of Obafemi Awolowo University Campus (IFE) and Forestry Research Institute of Nigeria Herbarium (FHI), Ibadan. The reagents used include acrylamide, monobasic sodium phosphate, dibasic sodium phosphate, tetramethylenediamine (TEMED), sodium dodecyl sulphate (SDS), ammonium persulphate (APS), bromophenol blue, 2-mercaptoethanol, glycerol, distilled water, Coomasic brilliant blue, methanol, glacial acetic acid.
.png)
Protein extraction was carried out by homogenizing 0.5 g of the species leaf sample in a porcelain mortar in 5 ml of 0.85% NaCl. The leaves extracts were centrifuged for 15 min at 3,000 revolutions per minute. The supernatant was removed after the precipitate had settled down and poured in sampling bottles, this was kept in the refrigerator till use.
Four to six (4 – 6) drops of the sample buffer was added to 10 – 12 drops of the protein sample, 0.2% bromophenol blue was added, and the sample was heated in a boiling water bath for 5 min. After boiling, the samples were introduced into each gel tube. 10% 2-mercaptoethanol was added to the sample to reduce the disulphide bonds in the protein so as to expose the protein to SDS action. The 5% SDS in an anionic detergent that distrupts nearly all non-covalent interactions in native proteins, thus the SDS forms a rod-like material with proteins that gives them uniform movement along the electrophoretic column according to their molecular weights. The glycerol added makes the protein sample denser so that it does not mix within the running buffer. The buffers are employed to maintain polyionic character of protein. Bromophenol blue was added to act as a tracer. The level of similarity of protein profile of the species was used to construct the dendrogram showing the coefficient of similarity.
RESULTS
The pattern of protein distribution in the species of Ludwigia studied and their diagrammatic representation are as shown in Figures 1 and 2, respectively. A keen examination of the bands shows that there are different patterns of band distribution in the genus. Marked differences were recorded for number, combination of bands and intensity of bands between species. The relationship between all the species of Ludwigia on the basis of band distribution is as shown in Table 2. The bands range from one to four (Figure 2). Most of the bands were found to be fast moving bands (2.0 to 3.5 cm), followed by the intermediate moving bands (1.0 to 1.9 cm), and slow moving bands (0.0 to 0.9 cm), respectively. Table 3 shows the common band relationship between all the Ludwigia species studied. L. octovalvis var linearis and Ludwigia hysopifolia have similar bands with the same intensity at 1.6 cm; L. octovalvis var linearis, Ludwigia hysopifolia and Ludwigia abysssinica have similar bands at 0.5 cm but with varying intensity.
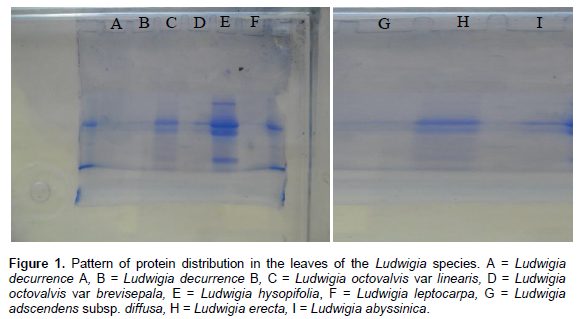
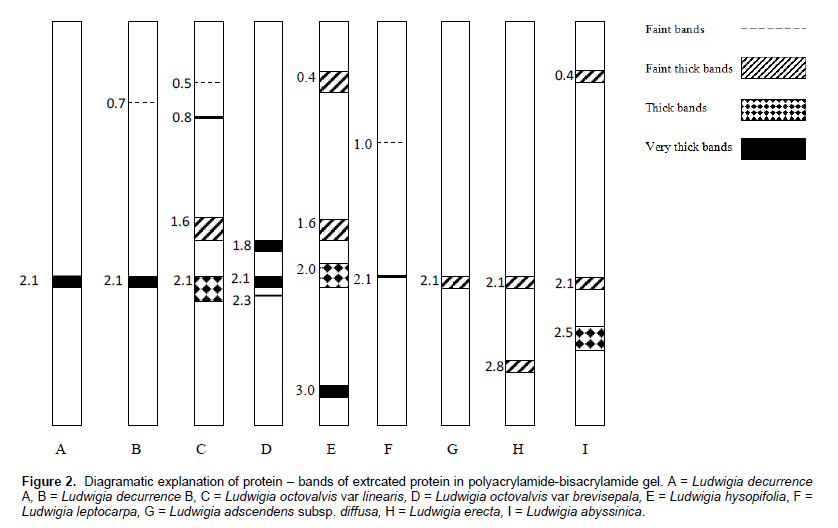
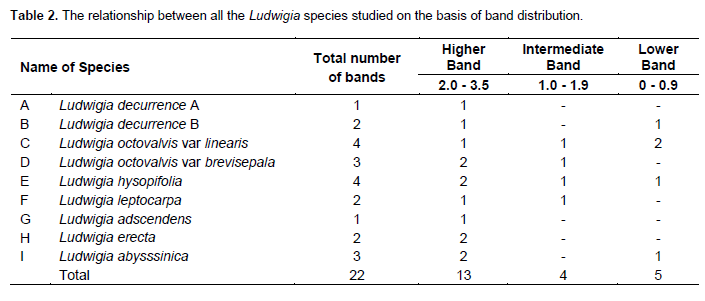
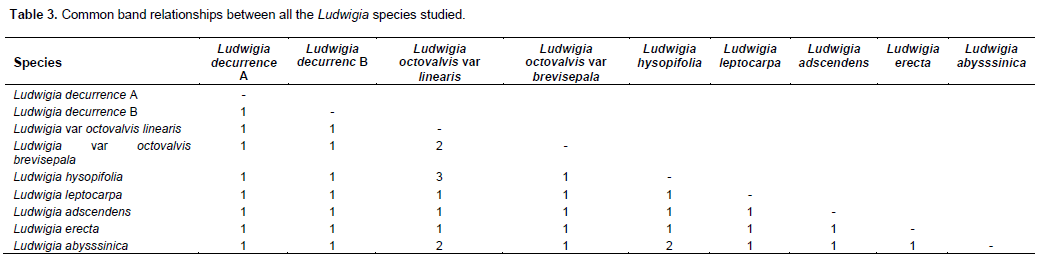
There are diagnostic bands peculiar to each of the species which makes individuals different from one another: Ludwigia decurrence B shows band at 0.7 cm, L. octovalvis var linearis at 0.8 cm, Ludwigia leptocarpa at 1.0 cm, Ludwigia octovalvis var brevisepala at 1.8 cm, Ludwigia abysssinica at 2.5 cm, Ludwigia erecta, at 2.8 cm and Ludwigia hysopifolia at 3.0 cm. The band at 2.1 cm was found to be common to all the species and occur at different intensities. Inter specific bands were observed between pairs of species in the genus. L. octovalvis var linearis and Ludwigia hysopifolia were found to have the highest number of bands (four), followed by Ludwigia octovalvis var brevisepala and Ludwigia abysssinica with three bands. Ludwigia decurrence B, Ludwigia leptocarpa and Ludwigia erecta have two bands while Ludwigia decurrence A and Ludwigia adscendens have one band (Figure 2).
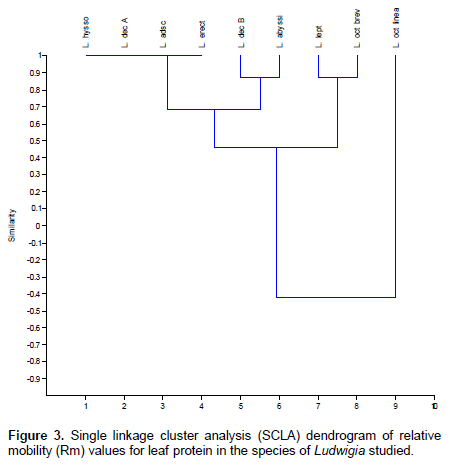
The dendrogram depicts the similarity coefficient for the Ludwigia species based on protein banding patterns (Figure 3). Three major clusters were formed, the first cluster comprises of L. hyssopifolia, L. decurrence A, L. adscendens and L. erecta at 1.0 similarity level. The second cluster comprises L. decurrence B and L. abyssinica at 0.85 similarity level. These two clusters are similar at 0.7 similarity level. The third cluster comprises of L. leptocarpa and L. octovalvis brevisepala, they are similar to the first two clusters at 0.45 similarity level. L.octovalvis linearis is delimited from the other species of Ludwigia studied at -0.4 similarity level similarity level. An artificial key for the identification of the Ludwigia species studied based on their band relationships is as shown in Table 4.

DISCUSSION
The result shows that no two species of Ludwigia have the same number and intensity of protein band. This affirms the morphological identification of the species studied, that is, the protein banding pattern is a reflection of their morphological characteristics. Protein variation in the species of Ludwigia studied is an indication of protein polymorphism; this depicts the genetic divergence in them and at the same time forms the basis of the separation of individuals in a particular population into different taxa (Ladizinsky, 1983). This genetic divergence is reported for Ludwigia decurrence A and B, with Ludwigia decurrence B a novel band in its gel which was not reported for Ludwigia decurrence A. The presence of this new band in Ludwigia decurrence B therefore separates it from Ludwigia decurrence A and a new hypothetical name may be given to delimit the species. Similarly, the diagnostic bands that separate Ludwigia octovalvis varieties are reported, these are basically the bands at 0.5 and 0.8. The Single Linkage Cluster Analysis reveals the true similarity position of the Ludwigia species studied.
Bands with identical electrophoretic mobilities represent proteins with identical amino acid sequences and are therefore potentially homologous in their derivations (Scogin, 1972). L. hyssopifolia and L. abyssinica have proteins with higher molecular weights which informs why they have slow moving band at 0.4. This band therefore delimits them from the other species; other diagnostic bands for the identification of each species are reported in the results.
The protein bands are taxonomically distinct as no two species have the same band distribution. This agrees with the opinion of Olsson (1967) as reported by Folorunso et al. (2012) that biogenetic relationships can best be indicated by quantitative results using chemo-taxonomic methods. The band at 2.1 is taxonomic for all the species of Ludwigia studied; this shows evidence of common evolutionary origin in them. Coming from the same parental stock, their evolution is convergent thereby making it possible for character traits to be shared in common. This support the assertion of Gottlieb (1971) that when a band appears in all individuals in a population, it is assumed that the gene which codes enzyme or protein does not vary. Based on the position of protein bands L. octovalvis var linearis, L. octovalvis var brevisepala, L. hyssopifolia and L. abyssinica are more closely related. The band at 1.0 delimits L. leptocarpa from the other species. L. decurrens A and L. adscendens subsp. diffusa are more closely related based on the number of band and the position of band.
CONCLUSION
Taxonomic bands for Ludwigia species studied have been reported together with the diagnostic bands for their identification. From the Dendrogram, interspecific relationships as well as their intraspecific relationships based on protein bands were reported. An artificial key was generated for the identification of the Ludwigia species studied based on their protein band relationship.
CONFLICT OF INTEREST
The authors have not declared any conflict of interest.
REFERENCES
| Cronk QCB, Fuller JL (1995). Plant Invaders: The Threat to National Ecosystems. Chapman and Hall: London; 241 | ||||
| Dutartre A, Dandelot S, Haury J, Lambert E, Le Goff P, Menozzi MJ (2004). Les jussies: characterisation des relations entre sites, populations et activites humaines. In: Implications pour la gestion. Rapport intermediaire programme INVABIO, Cemagref, Bordeaux. p. 44. | ||||
|
Folorunso AE, Adelalu KF, Oziegbe M (2014). Use of foliar and stem anatomical characters in the identification of Ludwigia species in Nigeria. Int. J. Biol. Chem. Sci. 8(5): 2232-2243. Crossref |
||||
| Folorunso AE, Akinwumi KF, Okonji RE (2012). Comparative studies of the Biochemical parameters of the leaves and seeds of Moringa oleifera. J. Agric. Sci. Tech. B 2:671-677. | ||||
|
Folorunso AE, Awelewa OA, Adewale IO (2006) not cited. Comparative study of protein profiles of the leaves of wild Manihot glaziovii Mueller and the cultivated species, Manihot esculenta. Int. J. Agric. Res. 1(1):53-57. Crossref |
||||
|
Gottlieb LD (1971). Gel electrophoresis: New approach to the study of evolution. Biosci. 21(18):939-944. Crossref |
||||
| Kadiri AB, Olowokudejo JD (2010). Systematic significant of foliar epidermal morphology in the West African species of Ludwigia (Onagraceae). Phytologia Balcanica, 16(1):57-64. | ||||
| Kadum YH, Manaf UA, Fariza SS (2012). Toxicological evaluation of 80% methanol extract of Ludwigia octovalvis (Jacq.) P.H. Raven leaves (Onagraceae) in BALB/c mice. National Centre for Biotechnology Information, U.S. National Library of Medicine 8600 Rockville Pike, Bethesda MD, 20894 USA. | ||||
| Ladizinsky G (1983). Study of Evolutionary Problems by Means of Seed Protein Electrophoresis. In Seed Protein: Biochemistry, genetics and nutritive value, (Werner Gottschalk and Herman P. Muller (editors), Martins NJHOff/DR.W Junk Publishers. The Hague/Boston/London, pp. 487. | ||||
| Oladipo OT, Illoh HC, Odekanyin OO (2008). Crude protein electrophoresis of seeds of four Nigerian species of Jatropha Linn. (Euphorbiaceae). Ife J. Sci. 10(2):263-267. | ||||
| Oladipo OT, Illoh HC (2012). Leaf protein electrophoresis and Taxonomy of Species of Jatropha L. (Euphorbiaceae). Not. Sci. Biol. 4(3):92-96. | ||||
| Olsson U (1967). Chemotaxonomic analysis of some cytopes in Methta X verticallate complex (Labiatae). Botanisca Notiser 20:255-267. | ||||
| Scogin R (1972). Protein in the genus Lithops (Aizoaccar): Developmental and comparative studies. J. South Afric. Bot. 39(1):55-61. | ||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


