Full Length Research Paper

ABSTRACT
In this study the microbiota of Clarias gariepinus tissues harvested from e-waste-soil polluted vials were assessed. Soil samples contaminated with e-waste were analyzed using standard analytical protocol while the microbial study was obtained using standard conventional microbiological techniques. The results revealed that the soil was sandy-loamy and blackish in colour. High organic matter (17.60%) and organic carbon (10.17%) were obtained. Also, higher calcium (182.00 mg/kg) and phosphorus (146.65 mg/kg) contents compared to other mineral constituents were recorded while, the heavy metals ranged from 0.32- 64.90 mg/kg. Bacterial count ranged from 9.0 × 102 to 4.0 × 103 cfu/L while the fungal count from 4.0 × 102 to 2.3 ×103 sfu/L. The genera of bacteria isolate were identified as Staphylococcus, Proteus, Bacillus, Listeria, Salmonella, Enterobacter, Pseudomonas, Lactobacillus and Corynebacterium and fungal isolates were Penicilliun, Candida, Articulospora, Aspergillus, Rhizopus, Mucor, Zoopage, Varcosporium and Rhodotorula. Microbial species richness of fish tissues from polluted vials indicates a more poly-diverse microbial community compared to those from unpolluted vial. Differences were observed in the occurrence of fungi on the surfaces of catfish tissues in control vial compare to those on fishes from polluted vials. Hence, there is a need for proper water management for fish culturing and caution in the exploration of fishes from polluted natural waters for human consumption.
Key words: Clarias gariepinus, e-waste, fish tissues, microbiota, pollution, microbial species richness.
INTRODUCTION
The United Nations Food and Agricultural Organization predicts that the world’s food and feed supply need to grow by 0.7-fold (70%) to sustain the increasing human populace by mid of next century (Bruijn et al., 2018). Currently, the cultivation of aquatic animals is the fastest growing animal food international sector (Bruijn et al., 2018). However, fish production has been hindered by numerous diseases caused by microorganisms and other ectoparasites. Fish tissues house different microbes whose communities is affected by physicochemical water parameter (such as pH, temperature, nutrient availability, dissolve O2 and salinity), environmental factors (pollution and season), host age, genotype, feeding approach and rearing conditions (Bruijn et al., 2018). There is growing evidence that microbial consortia rather than single species are linked to fish well-being and diseases (Gilbert et al., 2016). Constituents within man’s gut microbiota can substantially promote or suppress disease progress (Gilbert et al., 2016), whereas ecological changes or infections can significantly influence man’s gut microbiota by promoting proliferation of opportunistic microbes (Stecher et al., 2013). Similarly, plants’ microbial floral plays significant role in the defense against non-living physical and chemical elements (abiotic factors) and the living organisms (biotic factors) stress (Mendes and Raaijmakers, 2015). Likewise, fish microbial flora may possess considerable potential to influences well-being and disease. Owing to the intricate composition of microbial communities, disentangling relationships and identifying species for precise functions is hugely challenging, particularly when environmental impacts on population dynamics and processes are considered (Bruijn et al., 2018). Taxonomical or functional fluxes in the microbiota are fundamental factor for disease propagation or disease defense (Raaijmakers and Mazzola, 2016, Bruijn et al., 2018).
Fish tissues such as the fillet/skin, sensory system (olfactory system), gill system and the gut have contact directly with the surrounding and consequently are the initial points of microbial interaction with their host. More aerobic rather than anaerobic microbes are harbored by the mucus of the fish fillet and gill system (Merrifield and Rodiles, 2015). Microbiota composition of the gills’ and skin differs; the protected niches of the gill membrane have more microorganisms that putitatively favor gas exchange (Lowrey et al., 2015). Fish organs such as kidney, brain, liver and muscle under healthy condition are sterile, nonetheless there are scarcity of reports that have studied this in fish in detail (Bruijn et al., 2018). Likewise, there is a dearth of literature on the microbiota and functions of the communities of the fungal fish (Bruijn et al., 2018). Host genetics is also an important influential factor that determine the microbial populations of fish (Ghanbari et al., 2015). The composition of the gut microbiota is similarly depended on the microbiota composition in the environments (water and sediment), in spite of the importance of host genetical make up and feeding approach (Kashinskaya et al., 2015). A recent study shows the gut microbiota of salmon cultured in an aquarium and an open freshwater inlet cage system shared 1:97 operational taxonomic units (OTUs), demonstrating the environment substantially influences the composition of the gut microbiota (Dehler et al., 2017). Another environmental influence shaping the fish microbiota is water component or chemistry, this can be influenced by in flow of substances such as pollutants, into the water environment (Sylvain et al., 2016).
Evidently, these findings show that water condition has vital effects on fish microbiota. Microbiota is a complex and dynamic population of microorganisms that colonies the tissues of higher organisms, which exert influence on the host during homeostasis and disease (Thursby and Juge, 2017). The collection of microbes, colonising the tissue of higher organism has co-evolved with the host over the years to form an intricate and symbiotic relationship (Thursby and Juge, 2017; Bruijn et al., 2018). Modern techniques for sequencing DNA help to find most of these microbes because many of them cannot be cultured in the laboratory using conventional methods. Microbiota may have a role in auto-immune diseases (Bruijn et al., 2018). Imbalance in the microbial population of the gut may also aggravate digestive or intestinal problems (Willyard, 2011). Since some of the microbes in the body can modify the production of neurotransmitters known to occur in the brain, it may also relieve neuro-chemical imbalances (Bravo et al., 2011; Madigan, 2012). The microorganisms (non-pathogenic) in question, generally they do not cause illness except they grow abnormally. They exist symbiotically with their hosts such as fish (Salvucci, 2014). Disease causing microbes (pathogenic microorganisms) are basic component of fish microbiota, nonetheless their presence does not often lead to illness. On the other hand, when fish mutualistic or commensal microbial community balance is altered, pathogens proliferate and cause infection and disease, this process is known as dysbiosis (Moya and Ferrer, 2016). The imbalances that occur in the defensive symbiotic microbial community could result from changes in the surrounding (environment), including climate and seasonal changes, H2O conditions, temperature, changes in culturing parameters. Similarly, infection by a primary pathogen can lead to distortion in the microbial balance, this subsequently permits opportunistic disease-causing microbes (pathogens) to invade the host system (Llewellyn et al., 2017). In addition, symbiotic microbes play vital roles that contribute to host health and defense against disease-causing microbes (Xu et al., 2016). Some of these roles among others are direct defensive effects against disease-causing microbes by antibiosis and struggle for limited resources (niche exclusion or omission). Furthermore, these functions can be performed indirectly by inducing the host immune response then nutrient uptake thus promoting fish well-being. Symbiotic microorganisms could stop pathogen infection via several mechanisms, such as niche omission, which involves colonizing mucosal tissues then occupied infection sites and struggling for essential resources (Banerjee and Ray, 2017). The fish microbiota can induce a conserved host response in invasion (colonization) and development, and the fish microbiota communities evidently influences the inflammatory response (Bruijn et al., 2018).
Clarias gariepinus is freshwater fish species of African origin, is one of the most cultured species in the world (EmiroÄŸlu et al., 2018). This fish species is valuable for fish farming (aquaculture) due to its easy to farm especially in warm climates, fast growth rate, resistance to diseases, and high stocking density (EmiroÄŸlu et al., 2018). Additionally, many catfish species are important human food source. Monitoring of fishes’ microbiota such as that of C. gariepinus can be useful to evaluate their health status and fitness for human consumption.
Many of the water body in various parts of Nigeria receive huge quantities of untreated effluents and solid waste which contain substances that are harmful not only, to human but also to aquatic biota (Ekpo, 2012). In recognition of deleterious effects of the destruction and loss of habitat caused by solid waste, chemical pollution, eutrophication and climatic alterations on the aquatic organisms, as a result of human activities, combined with an urgent need of a more environmentally sensitive and ecologically sustainable management of water bodies in Nigeria, gingered the assessment of catfish microbiota harvested from polluted vials. Therefore, the main aim of this study is to create awareness on the multi-species diversity and probable health implication of the microorganisms found on the tissue of fish from polluted aquatic environment.
MATERIALS AND METHODS
Collection of samples
Electronic waste (e-waste) contaminated soil samples were collected from Alaba International Market, Lagos, Lagos State (Latitude: 6°27′14″ N, Longitude: 3°23′40″ E, Elevation above sea level: 11 m (36 ft)), Nigeria. E-waste is appliance consuming electricity and reaching the end of its life cycle. E-waste includes a wide range of electronic appliances ranging from large household appliances, such as refrigerators, air conditioners, cell phones, stereo systems and consumable electronic items to computers discarded by their users. E-waste generally contains valued metals such as copper, platinum group as well as potential environmental contaminants such as lead, mercury, nickel, selenium, cadmium, polybrominated diphenyl ethers (PBDEs) and polychlorinated biphenyls (PCBs). Most e-waste is disposed in landfills (Sanusi, 2015). The e-waste burning site is close to the Lagos water body; washing of surface materials from land surface to the sea or leaching of chemicals substances into it can be experienced.
Set up and pollution of vials
Four vials each containing six juvenile catfish were set up and polluted with three different concentrations of soil sample from e-waste dumpsite (0.025, 0.050 and 0.075 kg). This was carried out in the ratio of 1:1, 1:2, 1:3 of water to soil sample (25 L: 0.025 kg, 25 L: 0.050 kg, 25 L: 0.075 kg) after acclimatization of the fishes for 42 days. The fourth vial was not polluted it serves as the control. At the end of five weeks (35 days) bacteria and fungi were then isolated from both the vials and harvested fish tissues (skin, gill, intestine, kidney and liver).
Physiochemical parameters of soil sample
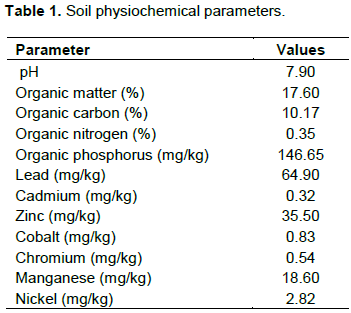
The pH of the soil samples and water samples in the aquaria were determined by the method described by Hendershot et al. (1993). From the soil samples, 10 g were weighted into a 100 ml beaker and 20 ml of distilled water was added to it. The mixture was stirred properly and allowed to stand for 30 min. 20 ml of the water samples were measured into a beaker and the electrode of the calibrated pH meter was dipped into sample water, the observed pH was read and recorded.
Extraction of heavy metals from soil samples was by acid digestion (Yusuf et al., 2015). The digestion was carried out with 20 ml of mixture of concentrate HClO4 and HNO3 at ratio 2:1 (v/v) on a hot plate and the mixture heated to almost dryness. 20 ml of 0.5 M HNO3 were added and the solution filtered into 50 ml volumetric flask through Whatman No.42 filter paper. The filtrate obtained was made up 50 ml mark with distilled water and used for heavy metal determination against those of the blank and calibration standards using a flame atomic absorption spectrophotometer (AAS), Apha 4 model. Phosphate concentrations in soil sample was determined using the phosphovanadomolydate colorimetric method (AOAC, 2019). Nitrogen determination was determined by direct nesslerisation method (AOAC, 2019). Organic carbon and organic matter determination were obtained using the methods describes by AOAC (2019).
Total plate count
Samples were taken from fish tissues using sterile swab sticks which were then placed in appropriate diluent for further studies. Plates in triplicates from harvested catfish tissues were observed for their bacteria and fungi loads. Colony and spore counting were carried out by counting the number of visible colonies and spores that appears on the plates for bacteria and fungi respectively. Calculation of colony forming unit (cfu) per ml for bacteria and spore forming unit (sfu) per ml for fungi were based on the dilution factor used.
Tentative microbial identification
Biochemical and morphological identification of bacteria isolates
Individual colonies were identified morphologically and by biochemical test using techniques described by Jesumirhewe et al. (2016). The biochemical tests performed to tentatively identified the bacterial isolates were; gram staining, catalase test, spore staining, motility test, starch hydrolysis, coagulase test and sugar fermentation test.
Identification of fungi
Probable fungi isolates were identified based on the cultural, morphological and microscopic examination of the colonies grown on potato dextrose agar (Onions et al., 1981). The parameters such as colony color, features of hyphae and shape of mature fruiting bodies were microscopically determined. The microscopic observations made were used in identifying the fungi isolate.
RESULTS AND DISCUSSION
The bacterial counts from the harvested tissues were observed to be highest on the skin (3.7 × 103) and the gills (4.0 × 103) (Figure 1). This is due to the constant interaction of these two tissues with the water environment. Although difficult to estimate and compare, studies have shown bacterial population in similar ranges (Austin, 2006; Merrifield and Rodiles, 2015). Bruijn et al. (2018), reported fish skin typically harbors 102-104 bacteria per cm2, whereas the gills harbour 103-106 bacteria per gram of tissue based on fish species, environment and cultivation methods. The lowest bacterial counts of 9.0 × 102 was observed in the intestine. The low bacterial population in the intestine can be attributed to the anaerobic condition around the gastrointestinal tract that will only support the growth or proliferation of anaerobic or facultative bacteria only (Ashlee et al., 2008). The fungal count followed a pattern which showed fungal population reduces with increase in the concentration of pollutant in the vials (Figure 2). This suggests that fungal proliferations were largely influenced by the presence of the pollutant. Sylvain et al. (2016), reported water chemistry, an environmental factor that can be influenced by pollution, also determines the fish microbiome composition. Microbial communities in natural aquatic environments respond rapidly to changes in their immediate environment (Bentzon-Tilia et al., 2016). For instance, the gut microbiota of salmon cultured in an aquarium facility and in an open freshwater inlet birdcage system, indicates the environment substantially influences the composition of the gut microbiome (Dehler et al., 2017).
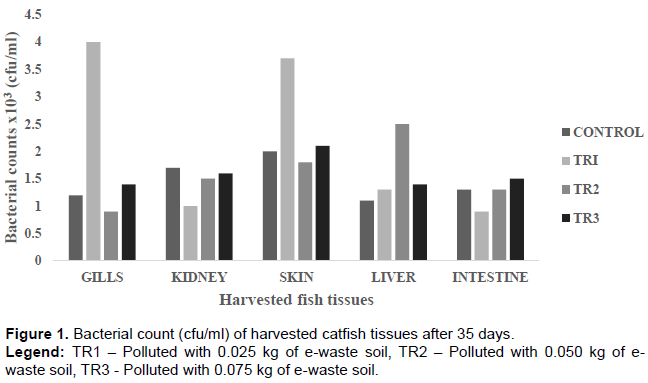
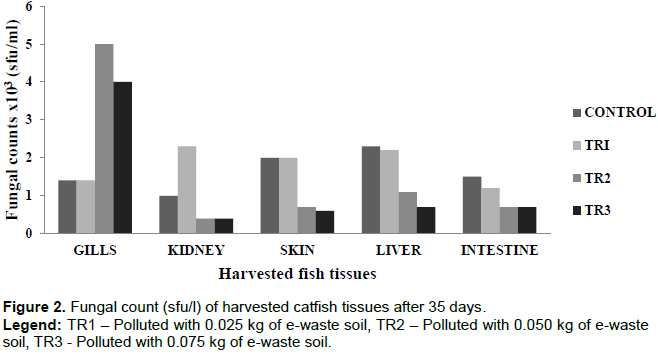
Electronic waste (e-waste) soil polluted vials had higher number of microbial isolates comparison to the unpolluted vials. This trend was the same for microbes from fish tissues harvested from the polluted vials; where fish tissues from polluted vials showed more species richness (diversity of microbial species) than those from unpolluted vials. These could be attributed to the organic contents of the e-waste soil sample (Table 1), which promoted and supported the growth of those microbes in the polluted vials (Bentzon-Tilia et al., 2016). This increased species poly-diversity of microbial community of the polluted vials and fish tissues from polluted vials might impact microbial balance in the polluted vials, on the fish and its tissues. The microbial imbalance can lead to unhealthy relationships within the microbes’ community (such as parasitism and inhibition effect) and between the fish and the microbes. The relationship between the fish and these microbes could also be pathogenic. Fungus such as Rhodotorula has been reported to cause infections in aquatic organisms. Fermanda and Luciano (2012), documented that Rhodotorula causes skin infections in both terrestrial and aquatic animals such as chickens, sea animals and lung infections and otitis in sheep and cattle. Dantigny et al. (2005), reported that some of these fungi secrete aflatoxins and other substances that can inhibit the growth of other microorganisms which could lead to declination of bacteria or fungi population. Also, the microbial colonization in and around the gastrointestinal tract of the fish could influence the microbial composition of the digestive tract. This might be of positive effect such as probiotic benefits. Symbiotic gut microorganisms similarly aid the fish in nutrient acquisition (Borrelli et al., 2016). The gut microbes can release exogenous enzymes to promote the digestion of food and degradation of chitin, protein, starch and other large and complex molecules (Montalban-Arques et al., 2015).
Similarly, they produce vitamins and eicosapentaenoic acid that is vital for metabolism to improve the well-being of the host. On the other hand, the effect might be negative, which can lead to poor feeding, impact digestibility rate and fish weight loss. Imbalance in the mix of microbial populations of the gut can also promote or aggravate intestinal problems (Ashlee et al., 2008; Willyard, 2011).
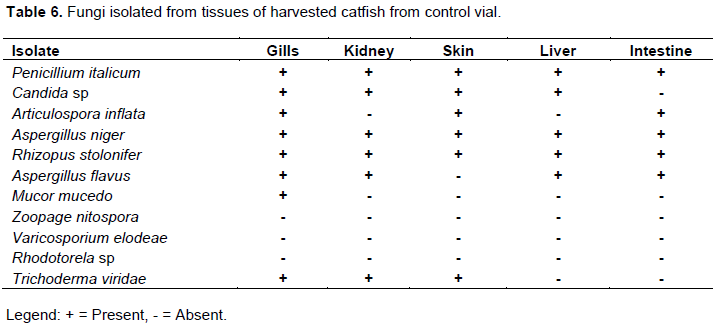
Bacteria isolates from the fish environment (water) were similar to those isolated from the fish tissues (Tables 2 to 3). The microorganisms associated with fish or their tissues are usually dictated by the environment the fish lives (Ekpo, 2012; Dehler et al., 2017). Many researchers (Sugita et al., 1997; Shewan, 2000; Okaeme, 2006, Bruijn et al., 2018) have isolated different species of bacteria from the skin of fresh water fish including Bacillus species from the skin of sea water fish. Sugita et al. (1997), reported that Staphylococcus spp, Escherichia coli were isolated frequently from the skin of fresh water fish while some researchers concluded that predominant genera are Pseudomonas, Staphylococcus and the member of the family Enterobacteriaceae on the skin of fresh water fishes, these agrees with their implication in this study.
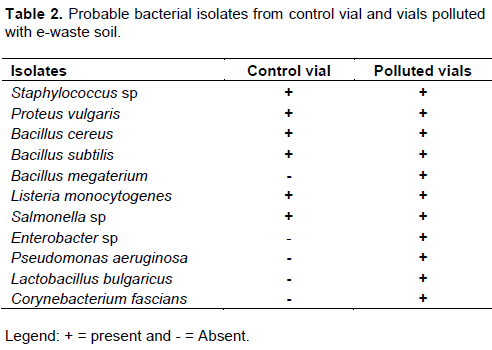
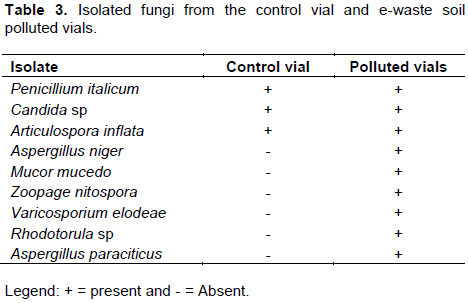
Eleven individual isolates of bacteria and twelve fungi (Tables 2 to 7) were isolated from polluted fish aquaria and harvested fish tissues. Staphylococcus aureus, Bacillus cereus, Bacillus subtilis, Bacillus megaterium, Listeria monocytogenes, Lactobacillus bulgaricus, Pseudomonas aeruginosa and Corynebacterium fascians have been implicated as normal fish floral which are dependent on the environment in which the fish lives, cultured or fish feed (Gram et al., 2000). S. aureus, Salmonella sp, P. aeruginosa, Aspergillus flavus, Mucor mucedo are linked to fish spoilage (Ashie et al., 1996, Gram et al., 2000, Doyle, 2007; Krijgsheld et al., 2013). Their presence can be attributed to the mortality of some of the catfish at the early stage of the study. Furthermore, Bacillus spp, P. aeruginosa, Articulospora inflata, Zoopage nitospora, Varicosporium elodeae, Penicillium italicum, Aspergillus niger, Aspergillus flavus, Rhizopus stolonifer, Mucor mucedo, Candida sp and Rhodotorula sp have been implicated in other environments such as petroleum polluted environment, gastrointestinal tract, agrarian soil, thus their isolation in the current study demonstrates or suggests their ability to survival under different environmental conditions and substrate utilization versatility (Ashlee et al., 2008; Alfreda and Ekene, 2012; Krijgsheld et al., 2013; Fermanda and Luciano, 2012).



CONCLUSION
This study demonstrates the diversity of catfish microbial populations elucidated with laboratory scale polluted aquarium system and discusses the potential implications of these microbes. Microbial species richness of fish tissues from polluted vials indicate that pollution increased the microbial colonization of these fish tissues. There are also differences in the occurrence of the microbes on the tissues of fish from the control vial and the polluted vials. These might not be of wholesome benefit to the fish. Overall, fish microbiota has enormous potential for fish health and disease, helping the host in its defense against pathogen colonization and infection or promote disease development leading to its immune defense being overwhelmed.
Therefore, proper management of waste disposal, in this case e-waste is necessary. E-waste contamination in aquatic environment can affect the quality of aquatic environment as well as promote microbial proliferation, which could be unhealthy to fishes and also reduce their quality for human consumption.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENT
The research was financially supported by Costech Canada Inc. and this is gratefully appreciated.
REFERENCES
|
Alfreda ON, Ekene GO (2012). Bioremediation of crude oil polluted soil using bacteria and poultry manure monitored through soybean productivity. Polish Journal of Environmental Studies 21(1):171-176. |
|
|
Association of Analytical Chemists (AOAC) (2019). Official Method of Analysis: Association of Analytical Chemists. 21st Edition. Washington DC. |
|
|
Ashie INA, Smith JP, Simpson BK (1996). Spoilage and shelf life extension of fresh and shell fish. Critical Reviews in Food Science and Nutrition 36:87-121. |
|
|
Ashlee ME, Richard L, Roberto K (2008). Ecology and genomics of Bacillus subtilis. Trends in Microbiology 16(6). |
|
|
Austin B (2006). The bacterial microflora of fish, revised. Science World Journal 6:931-945. |
|
|
Banerjee G, Ray AK (2017). The advancement of probiotics research and its application in fish farming industries. Research in Veterinary Science 115:66-77. |
|
|
Bentzon-Tilia M, Sonnenschein EC, Gram L (2016). Monitoring and managing microbes in aquaculture -Towards a sustainable industry. Microbial Biotechnology. 9(5):576-584. |
|
|
Borrelli L, Aceto S, Agnisola C (2016). Probiotic modulation of the microbiota-gut-brain axis and behaviour in zebrafish. Scientific Reports 6:30046. |
|
|
Bravo JA, Forsythe P, Chew MV, Escaravage E, Savignac HM, Dinan TG, Bienenstock J, Cryan JF (2011). Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proceedings of the National Academy of Sciences 108(38): 16050-16055. |
|
|
Bruijn I, Liu Y, Wiegertjes GF, Raaijmakers JM (2018). Exploring fish microbial communities to mitigate emerging diseases in aquaculture. FEMS Microbiology Ecology 94(1):1-12. |
|
|
Dantigny P, Guilamart A, Bensoussan M (2005). Basis of predictive Mycology. International Journal of Food Microbiology 100:187-196. |
|
|
Dehler CE, Secombes CJ, Martin SA (2017). Environmental and physiological factors shape the gut microbiota of Atlantic salmon parr (Salmo salar L.). Aquaculture 467:149-57. |
|
|
Doyle EM (2007). Microbial food spoilage - losses and control strategies. Food Research Institute. University of Wisconsin-Madison. |
|
|
Ekpo IE (2012). Diversity and condition factor of fish species of Ikpa river at Nwaniba in Niger Delta, Nigeria, FUTA Journal of Research in Sciences 1(1):36-47. |
|
|
Thursby E, Juge N (2017). Introduction to the human gut microbiota. Biochemical Journal 474:1823-1836. |
|
|
EmiroÄŸlu O, Baskurt S, Aksu S, Giannetto D, Tarkan AS (2018). Standard weight equations of two sub-/tropic non-native freshwater fish, Clarias gariepinus and Oreochromis niloticus, in the Sakarya River Basin (NW Turkey). Turkish Journal of Zoology 42(6):694-699. |
|
|
Fermanda W, Luciano ZG (2012). Epidemiology of Rhodotorula: An Emerging Pathogen. Interdisciplinary Perspectives on Infectious Diseases 2012:D 465717, 7 p. |
|
|
Ghanbari M, Kneifel W, Domig KJ (2015). A new view of the fish gutmicrobiome: advances from next-generation sequencing. Aquaculture 448:464-475. |
|
|
Gilbert JA, Quinn RA, Debelius J (2016). Microbiome-wide association studies link dynamic microbial consortia to disease. Nature 535:94-103. |
|
|
Gram LL, Oundo JO, Bon J (2000). Shelf life of fish depends on storage temperature and initial bacteria load. Tropical Science 25:28-30. |
|
|
Hendershot WH, Lalande H, Duquette M (1993). Soil reaction and exchangeable acidity. In: Soil Sampling and Methods of Analysis (Carter M. R., ed.). Boca Raton, FL, USA: Lewis Publisher. pp 141-165. |
|
|
Jesumirhewe C, Ogunlowo PO, Olley M, Springer B, Allerberger F, Ruppitsch W (2016). Accuracy of conventional identification methods used for Enterobacteriaceae isolates in three Nigerian hospitals. Peer-Reviewed Journal 4:e2511 |
|
|
Kashinskaya EN, Belkova NL, Izvekova GI (2015). A comparative study on microbiota from the intestine of Prussian carp (Carassius gibelio) and their aquatic environmental compartments, using different molecular methods. Journal of Applied Microbiology 119:948-961. |
|
|
Krijgsheld P, Bleichrodt R, Van Veluw GJ, Wang F, Muller WH, Dijksterhuis J, Wosten HAB (2013). Development in Aspergillus. Studies in Mycology 74:1-29. |
|
|
Llewellyn MS, Leadbeater S, Garcia C (2017). Parasitism perturbs the mucosal microbiome of Atlantic salmon. Scientific Report 7:43465. |
|
|
Lowrey L, Woodhams DC, Tacchi L (2015). Topographical mapping of the rainbow trout (Oncorhynchus mykiss) microbiome reveals a diverse bacterial community with antifungal properties in the skin. Applied Environmental Microbiology 81:6915-6925. |
|
|
Madigan MT (2012). Brock biology of microorganisms (13th edn.). San Francisco: Benjamin Cummings. ISBN 9780321649638. |
|
|
Mendes R, Raaijmakers JM (2015). Cross-kingdom similarities in microbiome functions. The ISME Journal 9:1905-1907. |
|
|
Merrifield DL, Rodiles A (2015). The fish microbiome and its interactions with mucosal tissues. In: Peatman BHB (ed.) Mucosal Health in Aquaculture. San Diego: Academic Press pp. 273-295. |
|
|
Montalban-Arques A, De Schryver P, Bossier P (2015). Selective manipulation of the gut microbiota improves immune status in vertebrates. Frontiers in Immunology 6:512. |
|
|
Moya A, Ferrer M (2016). Functional redundancy-induced stability of gut microbiota subjected to disturbance. Trends in Microbiology 24:402-413. |
|
|
Okaeme AN (2006). Fish diseases prevention and control paper presented at the VCN Professional country education seminar, Akure pp. 1- 17. |
|
|
Onions AHS, Allospp D, Eggins HOW (1981): Smiths Introduction to Industrial Mycology 7th ed. Edward Arnold (Publisher) Ltd, London WCIB 3DQ. |
|
|
Raaijmakers JM, Mazzola M (2016). Ecology. Soil immune responses. Science 352:1392-1393. |
|
|
Salvucci E (2016). Microbiome, holobiont and the net of life. Critical reviews in microbiology 42(3):485-494. |
|
|
Sanusi AI (2015). Impact of Burning E-waste on Soil Physicochemical Properties and Soil Microorganisms. British Microbiology Research Journal 8(2):434-442. |
|
|
Shewan JM (2000). The Microbiology of sea water fish. Academic Press, New York 1:487-560. |
|
|
Stecher B, Maier L, HardtW-D (2013). 'Blooming' in the gut: howdysbiosis might contribute to pathogen evolution. Nature Reviews Microbiology 11(4):277-284. |
|
|
Sugita H, Matsuo N, Hirose Y, Iwato M, Deguchi Y (1997). Vibrio species. Strain NM 10 with an inhibitory effect against pasteurellapiscicida form the intensive of Japanese coastal fish. Applied Environmental Microbiology 63:4986-4989. |
|
|
Sylvain F-E, Cheaib B, Llewellyn M (2016). pH drop impacts differentially skin and gut microbiota of the Amazonian fish tambaqui (Colossoma macropomum). Scientific Reports 6:32032. |
|
|
Willyard C (2011). Microbiome: Gut reaction. Nature 479 (7374):S5-S7. |
|
|
Xu Z, Takizawa F, Parra D (2016). Mucosal immunoglobulins at respiratory surfaces mark an ancient association that predates the emergence of tetrapods. Nature Communication 7:10728. |
|
|
Yusuf AJ, Galadima A, Garba ZN, Nasir I (2015). Determination of some Heavy Metals in Soil Sample from Illela Garage in Sokoto State, Nigeria. Research Journal of Chemical Sciences 5(2):8-10. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


