Full Length Research Paper

ABSTRACT
The study was carried out between September 2011 and August 2013, using 328 Clarias gariepinus in Oba reservoir, Oyo State, Nigeria. Standard methods were used to determine gonado-somatic index (GSI), fecundity values, and enteroparasitic infestation of the fish. Sex ratio was 1.3:1, male to female, and females on an average invested 20.54% of its body weight in egg production while male GSI was 6.05 ± 0.19%. Spawning was prominent in rainy season; the fish was highly fecund (31,169 eggs) with relative fecundity of 222 ± 10 eggs per gram weight of fish. Fecundity correlated linearly (positive) with gonad weight, body weight, total length and GSI. Over fishing, led the fish to invest its body weight and growth in egg production, to increase and enhance its chances of survival. Parasites recovered were Procamallanus laevionchus, Paracamallanus cyathopharynx, Anomotaenia species, Monobothrium species, Polyonchobothrium clariae, and Neoechinorhynchus rutili. Prevalence and intensity were higher in female than in male and more in dry than rainy season. Parasitic infestation reduces weight growth in the fish; this influenced the values of reproductive indices in which body weight was a numerator or denominator.
Keywords: Clarias gariepinus, enteroparasites, fecundity, gonado-somatic.
INTRODUCTION
Studies on fish fecundity are important to evaluate the reproductive potentials of the species (Duarte and Araujo, 2002). It gives prerequisite information needed to plan a breeding programme so as to determine how many eggs would be spawned or stripped, the amount of rearing facilities required and the extent to which various culture equipment would be put to use (Eyo and Mgbenka, 1992). Clarias gariepinus (Burchell, 1822) belongs to family Clariidae, and can survive low water levels and associated reduced oxygen levels due to their possession of accessory air breathing organ (Akinsanya and Otubanjo, 2006). The species is important for the economy of Ogbomoso and surrounding settlements, it is actively caught by the local fishermen because of its high growth rate. It is resistant to handling and stress, making it easy to be preserved and well appreciated because of its palatable taste (Sotolu, 2010). Reproductive biology of members of the Clariidae family and other culture-able species has been studied by many researchers across the globe particularly in Africa and there has been specific research on fecundity. C. gariepinus is a fish that is highly fecund as reported by Fawole and Adewoye (2004) and Inyang and Ezenwaji (2004).
Parasitic diseases of fish are very common all over the world and are of particular importance in the tropics (Roberts and Janovy, 2000). The supply of proteinous food in Africa is very expensive, faced with a supply constraint; attention has been drawn to aquaculture as a means to combat protein deficiency in developing countries (Komolafe, 2004). The need to culture fishes for dietery protein in a developing country like Nigeria, have made it necessary to intensify studies on the parasite fauna of the African freshwater fishes. C. gariepinus is known to possess one of the richest parasite fauna, which can be attributed to its diet and the number of predator species feeding on them (Jansen et al., 2013). Procamallanus laevionchus, Paracamallanus cyathopharynx (Nematodes), Anomotaenia species, Monobothrium species, Polyonchobothrium clariae (Cestodes) and Neoechinorhynchus rutili (Acanthocephalan) were recovered from the stomach and intestine of C. gariepinus from Erinle Reservoir in Osun State (Ajala and Fawole, 2014a) and also from Oba Reservoir in Oyo State (Ajala and Fawole, 2014b), Southwest Nigeria.
Some parasites have been discovered to have zoonotic potential in mammalian host including man, thereby making them of public health importance. Fish borne zoonotic trematodes (FZT) are transmitted to humans as metacercariae cyst in fish. Recent global health assessments have identified FZT as among the most important (and neglected) parasitic zoonotic diseases (FAO/WHO, 2012). They are an emerging problem and there is a consensus that fish caught in the wild and fish produced in aquaculture, produces a major food safety risk; especially in Southeast Asia where aquaculture is important economically (Clausen et al., 2015). Apart from the threat to man, parasitic infestation/infection of fish, affects their appearance, growth, reproduction, performance, reduces their relative values, palatability, and production (Hecht and Endeman, 2007). This study investigated some aspects of reproductive indices and enteroparasitic infestation of C. gariepinus, to determine the effect of parasitism on reproductive indices.
MATERIALS AND METHODS
Study area
The study area was Oba Reservoir in Ogbomoso, Oyo State, Nigeria. Oba Reservoir was impounded in 1964 and the tributaries are Idekun, Eeguno, Akanbi-Kemolowo, Omoogun, and Yakun streams. The main body of Oba Reservoir lies between Latitude 08° 3? N to 08° 12? N and Longitude 04° 6? E to 04° 12? E. The vegetation of the shoreline is dense with submerged macrophytes which decompose in the rainy season. The substratum of the reservoir is mainly mud and sand. The reservoir sustains a thriving artisanal fisheries industry and serves as a source of domestic water for residents of Ogbomoso North, and South local governments. During the period of investigation, rains were observed to fall in the months of March to September regarded as rainy season while the months from October to February were regarded as dry season (Ajala, 2015).
Collection of fish and data
Samples of the fish species (C. gariepinus) were purchased monthly from catches of local fishermen using traps, gill nets, and cast nets in the reservoir. Collection of fish specimens was done from September 2011 to August 2013, every two weeks between 06:00 and 08:00 am. All fish specimens were still alive at the time of purchase. The fish were transported in containers containing water from the reservoir to the research laboratory in the Department of Pure and Applied Biology, Ladoke Akintola University of Technology, Ogbomoso, Nigeria for further investigations. A total of 328 specimens were studied during the two year period. In the first year, September 2011 to August 2012 (referred to as 2011/2012), 166 specimens (90 males; 76 females); the second year, September 2012 to August 2013 (referred to as 2012/2013), 162 specimens (96 males; 66 females) were used in the study.
Identification of C. gariepinus specimens was done using fish identification guides by (FAO, 1992; Olaosebikan and Raji, 1998). Sexes were identified morphologically (Akinsanya and Otubanjo, 2006). The sexes were confirmed after dissection with the presence of testes in males and ovaries in females.
Length and weight relationship (LWR)
The LWR was estimated by using the logarithmic linear equation (Froese, 2006):
Log10 W = Log a + b Log10 L
where W = Total weight (g), L = Total length (cm), a = intercept, and b = regression coefficient.
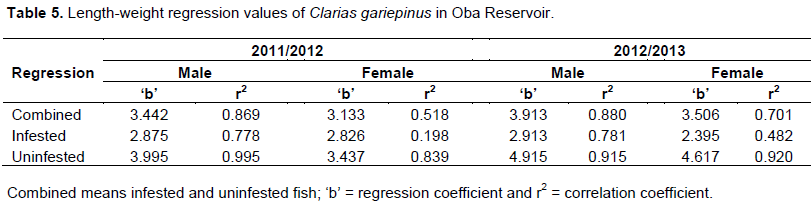
The relationship between total length and body weight was done by linear regression (Hossain et al., 2006). The length-weight regression equation was determined and the parameters ‘a’ (regression constant), ‘b’ (regression coefficient) and ‘r’ (correlation coefficient) were estimated, using the SPSS version 15 for Windows. These were done for male and female infested fish; male and female uninfested fish and male and female fish (infested and uninfested combined) harvested in 2011/2012 and 2012/2013.
Reproductive parameters
Reproductive activities in fish studies were carried out by the determination of certain reproductive parameters such as the gonado-somatic index (GSI) which is the ratio of gonad weight to body weight; it is used to estimate reproductive condition. The GSI is helpful in identifying days and seasons of spawning, during which ovaries of gravid females increase in size prior to spawning; it is indicative of reproductive success (Allison et al., 2005).
Each specimen was weighed using a top loading Metler balance (model PN1200) to the nearest 0.1 g after they were blotted dry with filter paper, while total length (TL) was measured in centimeter (cm) using a measuring board. Total length was used because no evidence of cannibalism was observed during the pre-data and data collection periods.
Numbers of males and female sexes harvested per year were counted and the ratio determined. The sex ratio was given as males: females (M: F), calculated using the formula: total number of males/total number of females (Vazzoler, 1996). Chi-square (c²) was used to verify the existence of significant differences between the sex ratio of the study species and commonly expected 1:1 sex ratio (Sokal and Jamesrohlf, 1987).
Euthanasia was carried out by physical method. Cervical dislocation with the two hands or cervical transection using a knife, or other sharp instrument inserted caudal to the skull, to sever the spinal cord and cervical vertebrae followed by pithing. The body cavity was opened ventrally with the aid of a sharp knife and scissors. The mesentery and connective tissues, connecting loops of the gut and the gonads were cut and the organs separated. The male and female gonads were placed in separate dishes and weighed. The data were used for reproductive studies.
The mean of the weight of the gonads (total weight of right and left gonad) of all specimens were calculated for the males and females.
The gonado-somatic index (GSI) of each fish was determined as:
GSI = 100 (Gw/Tw)
where Gw = weight of Gonad and Tw = Total weight of fish (King, 1995).
Monthly mean values (GSI) of male and female were computed and presented in graphical forms to ascertain monthly variations and detect the period of high reproductive activity represented by the peak(s) in the value
Ovaries collected were weighed using an electronic Metler balance model P1210 to the nearest 0.01 g. The ovaries were preserved separately in modified Gilson’s fluid as described by Komolafe (2005). The preserved ovaries were periodically shaken to ensure the separation of eggs from ovarian tissues.
Fecundity is defined as the number of ripened eggs in the ovary, just before spawning. Sub-samples of ovaries were obtained and weighed. The number of eggs in each sub-sample was enumerated manually. The absolute fecundity in the fish was estimated from the relationship established by Coward and Bromage (2000) and Bradford et al. (2011) as:
F = ½ (N1/W1+N2/W2) W.
where N1 is the number of eggs in the first sub-sample with weight W1 (left ovary). N2 is the number of eggs in the second sub-sample with weight W2 (right ovary). W is the total weight of the pair of ovary from which the sub-samples were obtained.
A mean fecundity from all samples was calculated using direct summation procedure. The relationship between fecundity, total length, body weight, and gonad weight of fish was determined by exponential and linear regression using the formula (Bagenal, 1967):
F = aLb
where F = Fecundity; L = standard length in cm; a = intercept of the regression line with the y-axis (regression coefficient); b = slope of the regression line (regression constant).
The mean relative fecundity was calculated as (Bagenal and Tesch, 1978):
mFec / bwt and expressed in eggs/g body weigh
where mFec is mean fecundity of fish; bwt is body weight of fish.
Examination for parasites
Examination of fish for parasites, handling and processing were followed using standard procedure (Moravec, 2010). After euthanasia by cervical dislocation and pithing, the body of each fish was examined for abnormalities and placed on a dissecting board. The buccal cavity was washed with 1 ml of distilled water and a fine brush into a labeled test tube after which the body cavity was opened ventrally with a sharp knife and scissors. The mesentery and connective tissues, connecting loops of the gut and the liver were cut and the alimentary system was separated. The alimentary system was then placed in a large Petri dish and cut into two regions, that is, the stomach and the intestine. The intestine was further separated into small and large intestines. Each part was then placed in separate labeled dishes containing the stomach, small intestine and large intestine. The separated parts were opened longitudinally to expose the inner surface which was washed with 0.5 ml of distilled water into labeled test tubes. The residue from the mouth, stomach, small intestine and large intestine in the labeled test tubes were then examined. A drop of the residue was placed on the slide, and observed under various magnifications of the light microscope for the parasites. This was repeated until the entire residue has been examined. Parasites found were counted, labeled with the serial number of the fish and placed in physiological saline water overnight to allow them to stretch and relax (Ajala, 2015); they were then fixed and stained for identification to species level.
The identification of the parasites was enhanced by their wriggling movement on emergence. Fish specimens found with parasite were given separate serial numbers to differentiate them from those without parasites. Identification of specimens to species level was done as described by Oniye et al. (2004) and Akinsanya and Otubanjo (2006).
Cestodes and nematode parasites recovered were stained using the procedure of Khalil (1991). Fixative used was formalin acetic acid (FAA). Cestodes were stained using acetocarmine; nematodes were stained with Horen’s trichome stain; while acanthocephalans were preserved in weak Erlich’s haematoxylin solution overnight and dehydrated, cleared in methyl-salicylate and mounted on a slide in Canada balsam.
Statistical analyses
For the reproductive parameters, a regression graph was plotted between gonad weight and body weight of male and female specimens, the correlation coefficients r2 of good fit to the line of regression were determined using MS-Excel package. Significant differences between the means of GSI in the two years for both male and female fish were carried out using t test (2-tailed) at 95% level of confidence with SPSS version 15.0 for Windows.
Infection and infestation of host by parasites were not normally distributed; as such, significant differences of parasitic infestation were tested using Kolmogorov-Smirnov K-S test at 95% level of confidence using SPSS version 15.0 for Windows. Significant difference between the means of body weight and total length of infested and uninfested fish specimens were carried out using t-test (2-tailed) at 95% level of confidence with SPSS version 15.0 for Windows.
A regression analysis was carried out on the length-weights of infested and uninfested fish specimens as described by Hossain et al. (2006). The correlation coefficients r2 of good fit for infested and uninfested fishes were determined using MS-Excel package.
RESULTS
Seasonal distribution of C. gariepinus in Oba Reservoir
Seasonally, more fish were harvested in the dry season with a range of 0 to 13 fish specimens per month giving a total of 194, while in the rainy season an average of 0 to 8 fish specimens per month were caught giving a total of 134 during the period of study.
Total length values in male C. gariepinus were an average of 28.53±2.04 cm (male) and 29.00±1.89 cm (female) for the two years; the body weights were 132.07±10.08 g in males and 141.00±9.40 g in females. The monthly mean values of length and weight are shown in Table 1. The length-weight relationship of the fish in Oba Reservoir showed a positive linear relationship indicating that an increase in length leads to an increase in weight.
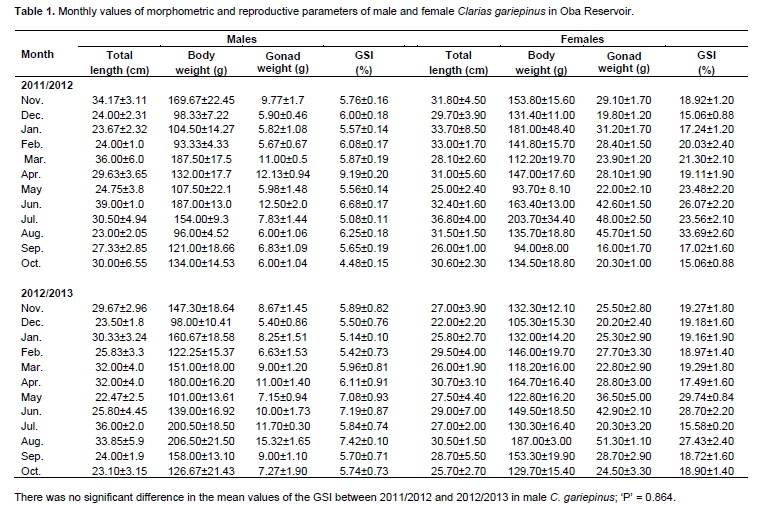
In the first year (2011/2012), sex ratio was 1.18:1 (male: female). In the second year (2012/2013), a ratio of 1.46:1 (male: female) was determined. In the two years, a ratio of 1.3:1 (male: female) was derived.
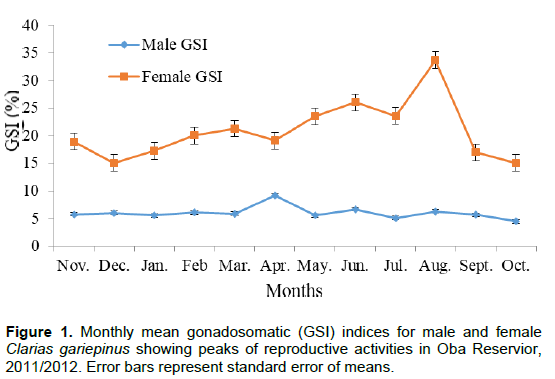
The monthly mean GSI values for males in 2011/2012 ranged from 4.48±0.15 in October to 9.19±0.20 in April (Table 1) with a yearly mean of 6.01±0.33. Reproductive activities in males of C. gariepinus in the year were from April to August, with peak periods in April, June and August (Figure 1). A regression graph plotted between gonad weight and body weight gave an equation of y = 0.049x + 1.323 with an r2 value of 0.505.
In 2012/2013, the mean values for male ranged from 5.14±0.10 in January to 7.42±0.10 in August (Table 1) with a yearly mean of 6.08±0.21. Reproductive activities in male C. gariepinus in 2012/2013 were from April to August, with peak periods in June and August (Figure 2). A regression graph plotted between gonad weight and body weight gave an equation of y = 0.059 x -0.594 with an r2 value of 0.902. The mean of the GSI for males in the two years was 6.05±0.19.
The monthly GSI values for females in 2011/2012 ranged from 15.06±0.88 in December and October to 33.69±2.60 in August (Table 1) with a yearly mean of 20.05±1.50. Reproductive activities in female in 2011/2012 were from May to August, with peak periods in June and August (Figure 1). A regression graph plotted between gonad weight and body weight of all female specimens studied gave an equation of y = 0.147x + 8.785 with an r2 value of 0.449.
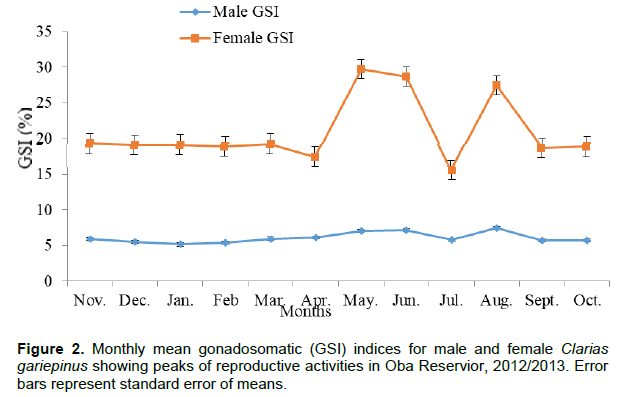
In 2012/2013, the mean values ranged from 15.58±0.20 in July to 29.74±0.84 in May (Table 1) with a yearly mean of 21.02±0.60. Reproductive activities in female in the year were from May to August, with peak periods in May, June and August (Figure 2). The mean of the GSI for females in the two years was 20.54±1.00.
A regression graph plotted between gonad weight and body weight of all female specimens studied gave an equation of y = 0.158x + 3.930 with an r2 value of 0.781. There was no significant difference (p > 0.05) in the mean values of the GSI between 2011/2012 and 2012/2013 in female C. gariepinus; the ‘P’ value was 0.095.
Fecundity
Absolute fecundity for the two years of study was 31,169 eggs. The mean relative fecundity in the two years of study was 222+10 eggs/gram weight of fish.
Regressions between morphometric and reproductive parameters
Enteroparasites recovered
Helminth parasites recovered comprised two nematodes (P. laevionchus, retrieved from the intestines, P. cyathopharynx retrieved from the stomach), three cestodes (Anomotaenia spp., Monobothrium spp., P. clariae) retrieved from the intestines and one acanthocephalan (N. rutili,) also retrieved from the Intestines. Table 4 shows the seasonal distribution and intensity of retrieved parasite species in the male and female fish hosts. The intensity of parasite species was higher in females than males in the two years of investigation.
Parasitic prevalence
The prevalence of the parasitic infestation among sexes was 30.12% in females and 24.10% in males in 2011/2012; in 2012/2013 it was 30.86 and 16.05%, respectively. Generally, it was higher in 2011/2012 (54.22%) than in 2012/2013 (46.91%) and in the two years, prevalence was 50.57%. Seasonally, prevalence was 65.96% in the dry season and 38.89% in the rainy season of 2011/2012; in 2012/2013, it was 52.08 and 39.39%, respectively.
Test of significance (t-test) at p ≤ 0.05, between the means of parasite load in fish specimens of 2011/2012 (104.00±3.24) and 2012/2013 (100.36±2.85) was not significant (p = 0.071). Also, between the means of parasite load in male (46.06±3.61) and female (67.81±8.93) in 2011/2012 was not significant (p = 0.061).
Test between the means of parasite load in male (22.75±2.53) and female (87.77±8.19) in 2012/2013 was significant (p = 0.041).
The t-test for significance at p ≤ 0.05 between the means of the body weights of infested and uninfested in the two sexes were significant in the two years of study: in females, p = 0.024, in males, p = 0.046.
Regressions between parasite intensity and body weight of infested fish in the two years showed the relationships were negatively correlated; In 2011/2012, the regression equation was y = -0.383x + 90.88; r² = 0.147; in 2012/2013, the equation was y = -0.321x +81.69; r² = 0.184.
Regression analysis carried out between the length and weight of infested specimens and between the length and weight of uninfested specimens, all showed positive linear relationships (Table 5).
DISCUSSION
Seasonally, more fish were captured in the dry seasons than the rainy seasons of the two years of study. The low percentage of fish captured in the rainy season could be as a result of high volume of water in the reservoir and C. gariepinus being a benthic species, will be difficult for fishermen using cast nets, gillnets and traps to capture because of accessibility to the bottom of the reservoir. The high water level will also be a hindrance as most fishermen will not go fishing when the water level is high.
The average total length and body weight values recorded were low compared to other values from other fresh water systems such as reported by Agboola and Anetekhai (2008). Yalcin et al. (2001) reported size at maturity for female and male C. gariepinus to be 25.05 cm TL, 108.7 g and 24.7 cm TL and 113.2 g, respectively. The low total length and weight recorded in this study may be as a result of over fishing. It was reported by Sotolu (2010), that the fish has resistance to handling and stress (making its preservation alive easy), and well appreciated because of its palatable taste. All these might have resulted in high demand for the species by the residents of the surrounding communities, putting pressure on the fishermen for increased catch. Also, inadequate enforcement of fishing regulations by the reservoir management, especially on the use of the right net mesh sizes might be responsible for the over fishing in the reservoir. This result agrees with Wootton (1992) who reported that the fish reach sexual maturity at an unusually small size in stunted population. Factors like pollution, uncontrolled hunting and other physical and chemical factors could also be responsible for the stunted growth (Yalcin et al., 2001).
Sex ratio showed that both sexes were available in the reservoir while males were more in number than the females (ratio of 1.3:1, male to female). Gonodal sex differentiation in gonochoristic fish is generally under the control of two interacting processes: genetic sex determination (GSD) and environmental sex determination (ESD) (Baroiller et al., 2009). Environmental conditions have variable effects on sex differentiation, depending on the genetic background and developmental stability of different strains. Temperature fluctuations could alter sex-determination pathways and influence the probability that development would be male or female (Guerrero-Estévez and Moreno-Mendoza, 2010). While low temperature is capable of biasing sex differentiation towards female in many fish species, elevated temperature has been shown to skew sex ratios towards males in some others (Guerrero-Estévez and Moreno-Mendoza, 2010). Santi et al. (2016) demonstrated that C. gariepinus displayed thermo sensitivity in the sex differentiation process, with a masculizing effect at high temperature, with a sex ratio skewed towards the male phenotype. Santi et al. (2017) also reported that C. gariepinus has a genetic sex determination system in which high temperature induces masculinization. The temperature values during the period of study were 27.37 ± 0.03°C in the first year and 27.36 ± 0.03°C in the second year, with no significant difference in the means of the two years (P = 0.083) (Ajala and Fawole, 2016a). The reservoir is located in the tropics (warm environment) and WHO (2011) opined that the temperature of inland waters depend on the ambient temperature. This might have influenced the high number of males recorded over females in the reservoir.
The mean of the GSI values obtained for females (20.54 ± 1.00) were relatively higher than those obtained for males (6.05 ± 0.19) in the two years of study (Table 1). This could be as a result of heavier gonads with the presence of eggs, possessed by female fish species. The GSI values recorded for males showed that reproductive activities were from April to August (Figures 1 and 2). In females, reproductive activities started from May to August and peaked in June and August (Figures 1 and 2), an indication that spawning is likely to be in the rainy season. The long rainy months (eight months) experienced during the investigation may be an added advantage for the fish species to spawn. The result obtained is in agreement with the findings of Ezenwaji (2002) and Midhat et al. (2013).
The mean of all GSI values for female specimen harvested was calculated as 20.54 ± 1.00 which means that the fish species on an average invested 20.54% of its body weight for egg production. This result was higher than 4.6% reported ten years earlier in the reservoir by Fawole and Adewoye (2004). The GSI value being an investment in egg production resulted in the low mean fecundity value (10,593 eggs) obtained by Fawole and Adewoye (2004) while 31,169 was obtained in this study. A positively linear relationship was observed from the regression of gonad weight (Gwt) and body weight (Bwt) in males and also in females. In males of 2011/2012, r2 = 0.505; while in females, r 2 = 0.449; when r 2 value is closer to infinity (that is 1.0), the two regressed data are said to be fitted to the line of regression. This result showed that the relationship was better fitted in males. This could be an indication that a factor was militating against the fish and the factor was more pronounced in females than males, the reason was probably because males had less parasitemia than female. The factor could have affected any of the indices used in calculating the GSI and it could be any of physico-chemical characteristics of the water in the reservoir, food availability/growth or parasitism. However, the positive linear relationship was an indication that generally, the gonad weight increases with the body weight.
Fecundity values obtained from matured females ranged from 13,719 to 48,514 eggs; with a mean value of 31,169.15 ± 1427.307 in the two years (Table 2). This result showed that the fish species was highly fecund. Hecht (1982) described the fish as a highly fecund species that spawn easily under captive conditions; disparity in fecundity values could be due to difference in the size, location and food availability (Inyang and Ezenwaji, 2004). Highly fecund and commercially important fish species can be easily subjected to over fishing and could be threatened with extinction as a result of over exploitation (Sadovy, 2001).
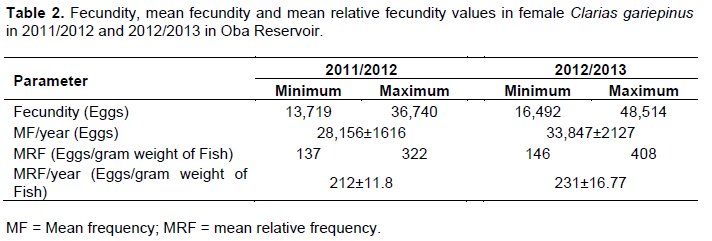
This, inadvertently probably necessitated investment of body weight in egg production, earlier sexual maturity and hatching as a survival strategy. The observation of highly fecund, low weight C. gariepinus observed in Oba reservoir could be as a result of reproductive strategies aimed at survival (Murua and Saborido-rey, 2003; Sadovy, 2001). Investigation conducted thirteen years earlier in the reservoir by Fawole and Adewoye (2004) reported a body weight range of 84.30 to 3,181.00 g; total length range of 24.20 to 56.70 cm; mean GSI of 4.6 and a mean fecundity of 10,593 eggs for C. gariepinus. In this study, a body weight range of 93.33 to 206.5 g; TL range of 20.90 to 39.00 cm; mean GSI of 20.54 and a mean fecundity of 31,169 eggs were obtained. The two results showed that in the last thirteen years, C. gariepinus in Oba Reservoir had not been given sufficient time to grow in size. This is possibly as a result of over fishing from local fishermen, due to high demand for the fish species in the surrounding communities. It may also be due to lack of enforcement of fishing regulatory laws and monitoring in the reservoir. Sharpe and Hendry (2009) were of the opinion that overfishing reduces fish populations and promote smaller sizes in fish, which also make the fish reach sexual maturity earlier than normal. Also the findings of Kuparinen et al. (2016) showed that fishing pressure is playing a major role in the life history changes observed in commercial fish stocks and is consistent with the hypothesis that fishing-induced phenotypic changes can sometimes have a genetic basis. It can thus be suggested that as a survival strategy, the fish probably sacrificed or invested its body weight in egg production, to increase and enhance the chances of its survival in the reservoir.
The mean of relative fecundity for 2011/2012 was calculated to be 212 ± 11 eggs per gram weight of fish and in 2012/2013 a value of 231 ± 17 eggs per gram weight was obtained. In the two years, 222 ± 10 eggs per gram weight of fish were obtained. This also showed that the fish species invested heavily in egg production as a means of survival. Fawole and Adewoye (2004) reported 58.3 eggs per gram body weight; Yalcin et al. (2001) reported 155 eggs per gram wet weight from River Asi, in Turkey. Fecundity in matured females were also observed to be correlated linearly (positively) with gonad weight, body weight, total length and GSI (Table 3).

The correlation showed that the gonad weight increased as the fecundity increases in the two years (r2 was 0.912 in the first year and 0.856 in the second year). This means the relationship was better fitted in the first year where 91.2% of the data collated could be explained by the relationship compared to 85.6% in the second year. However, the values of correlation with body weight in the two years were low (r2 was 0.483 in the first year and 0.405 in the second year) compared to values recorded for gonad weight (Table 3). This indicated that the gonad weight increased at a higher proportion with fecundity than the body weight during the period of study. This might have been as a result of the effect of parasite infestation on the gravid fish weight. The result also showed positive linear correlation of total length with fecundity in the two years. However, the total length did not increase with fecundity at the same rate as the body weight. The r2 values of correlation between fecundity and total length were low in the two years. Immature stages of C. gariepinus has been reported to feed on relatively higher proportion of insects and detritus (Dadebo, 2009) which serves as intermediate hosts to most of the parasites retrieved from the fish (Ajala and Fawole, 2016b). These probably increase the possibility of the fish getting infested with parasites early in life. Also, Omeji et al. (2011) reported that gravid fish have reduced resistance to infection. These would affect growth in weight and length in the infested fish. The correlation with GSI in the two years was very low and the reason would be because the body weight is a denominator in arriving at the GSI values. These results were in agreement with the report of Wootton (1992), that fecundity of fish increases as body size increases and that the volume of eggs a fish can produce depend upon the space available in the body cavity to accommodate eggs before spawning. Yalcin et al. (2001) also reported an r2 value of 0.9188 between fecundity and body weight, 0.7784 (fecundity and total length), 0.9645 (fecundity and gonad weight) in C. gariepinus in River Asi in Turkey. Anibeze and Inyang (2000) were of the opinion that fishes with a closer relationship between fecundity and body weight than what obtains between fecundity and length showed faster growth rate at the same time with fecundity and that the parameters to a large extent depend on environmental conditions. The reasons for the results in this study can be adduced to anthropological activities (over fishing) and parasitic infestation of the fish hosts in the reservoir. Also, the high fecundity recorded in this study is also an indication that the fish species will lack any form of parental care for the young.
Parasitic prevalence was found to be higher in females than males in the two years of study and it was higher in 2011/2012 (54.22%) than in 2012/2013 (46.91%). However, intensity/ parasitemia was higher in infested fishes in 2012/2013 than in 2011/2012 (Table 4). Parasitic intensity was higher in infested females than in infested males; intensity was also higher in the dry seasons than in the rainy seasons of the two years of study (Table 4). This showed that female C. gariepinus were more susceptible to parasitic infestation than males. This was also supported by the intensity of parasitemia, which was higher in females in virtually all the parasite species than in males in the two years of study (Table 4). This may be as a result of differential in feeding, either by quantity or quality of food eaten, or as a result of different degree of resistance to infection (Emere, 2000). Omeji et al. (2011) were of the opinion that due to the physiological state of the female, most gravid females could have reduced resistance to infection and infestation by parasites. The result was in consonance with the work of Ajala and Fawole (2014) who reported significant difference (p ≤ 0.05) in the infestation of male (48.78%) and female (51.22%) C. gariepinus from Oba Reservoir; Ajala and Fawole (2014a) also reported significant difference (p ≤ 0.05) between male (43.80%) and female (56.20%) of C. gariepinus from Erinle Reservoir in Osun State, Nigeria.

The t-tests between the body weights of infested and uninfested females in the two years were significant (p = 0.024) at p ≤ 0.05, while those of males (p = 0.046) though significant, but not as high as the value for females. Though the reproductive burden on the female is high, the parasitic infestation of C. gariepinus in Oba Reservoir had a greater effect on the weight of the female fish than it had on the weight of male fish. The length-weight relationship of the fish in Oba Reservoir showed a positive linear relationship indicating that an increase in length leads to an increase in weight (Table 5). The regression coefficient ‘b’ recorded for male and female specimens (when infested and uninfested were not separated) was higher than 3 (Table 5); this indicated that males and females exhibited positive allometric growth. In fishes which maintain dimensional equality, the isometric value (b) will be 3 (Bradford et al., 2011). Regression coefficient ‘b’ larger than 3 exhibited positive allometric growth while smaller than 3 demonstrated negative allometric growth (Shoesmith, 1990). A positive allometric growth means the fish body weight increase faster than the length increases. However, the ‘b’ values for infested fishes of both sexes in the two years were less than 3 (Table 5), indicating negative allometric growth. This showed the fish grow faster in length than the weight, an indication that the infestation had probably affected the growth in weight. A regression analysis between parasite intensity and body weight in infested fish for the two years were negatively correlated. This showed that as the intensity of parasite increases, the body weight reduces and vice versa. This also indicated that the gonado-somatic index values calculated with the fish body weight as a denominator, will decline as a result of the infestation, but its important role of signifying the spawning period of the fish will still be achieved.
CONCLUSION
Parasitic infestation in Oba Reservoir was found to reduce the growth in weight of the fish; thereby reducing the regression coefficient (b) to a value less than three. Gonado-somatic index will decline as a result of the infestation but its important role of signifying the spawning period of the fish will still be achieved. However, the effect may be overridden if the parasitic prevalence and intensity were low. The fish is highly fecund, and is capable of diverting its resources for growth, into more egg production, when it encounters problems such as pressure of over fishing in the aquatic habitat. Parasitic infestation of C. gariepinus by P. laevionchus, P. cyathopharynx, Anomotaenia spp., Monobothrium spp., P. clariae, and N. rutili reduces its body weight. Prevalence and intensity is higher in females than males, hence the females are mostly affected. With climate change and global warming, investigations should be carried out periodically to revalidate previously acquired data in an aquatic ecosystem and to detect developments or changes in the water body. It is suggested that fishing guidelines on use of mesh sizes for fishermen harvesting fish in Oba Reservoir, be enforced and monitored to prevent over fishing and protect the resident fish species.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Agboola JI, Anetekhai MI (2008). Length-weight relationships of some fresh and brackish water fishes in Badagry creek, Nigeria. Journal of Applied Ichthyology 24(5):623-625. Crossref |
||||
| Ajala OO (2015). Studies on aspects of Limnology and Biology of selected fish Species in Oba Reservoir, Ogbomoso, Nigeria. Unpublished PhD Thesis (Provide name of institutuion). | ||||
|
Ajala OO, Fawole OO (2014a). Multiple Infections of Helminths in the Stomach and Intestine of Clarias gariepinus (Burchell, 1822) In Oba Reservoir, Oyo State, Nigeria. IOSR Journal of Pharmacy and Biological Sciences 9(3V):05-12. Crossref |
||||
|
Ajala OO, Fawole OO (2014b). Multiple infections of helminths in the alimentary system of Clarias gariepinus (Burchell, 1822) in a tropical reservoir. International Journal of Fisheries and Aquaculture 6(6):62-70. Crossref |
||||
| Ajala OO, Fawole OO (2016a). Effects of water limnology and Enteroparasitic infestation on morphometrics of Oreochromis niloticus (Linne, 1757) (Cichlidae) in a tropical reservoir. European Journal of Advanced Research in Biological and Life Sciences 4(2):1-14. | ||||
| Ajala OO, Fawole OO (2016b). A study of diets and parasitic infestation in Clarias gariepinus (Burchell, 1822) in Oba Reservoir. Journal of Research in Applied, Natural and Social Sciences 2(2):27-38. | ||||
|
Akinsanya B, Otubanjo OA (2006). Helminth Parasites of Clarias gariepinus (Clariidae) in Lekki Lagoon, Lagos, Nigeria. Revista de Biologia Tropical 54(1):93-99. Crossref |
||||
| Allison ME, Sikoko FD, Vincent-Abu IF (2005). Fecundity, sex ratio, Maturity stages, size at first maturity, breeding and spawning, of Parailia pellucid (Boulenger, 1901) in the lower Nun river, Niger Delta, Nigeria. Caderno de pesquisa, sirrie Biology 20(2):33. | ||||
|
Anibeze CIP, Inyang NM (2000). Oocyte Structure, Fecundity and Sex Ratio of Heterobranchus longifilis (Valenciennes 1840) in Idodo River Basin (Nigeria) with comments on the Breeding Biology. Journal of Aquatic Sciences 15(1):59-62. Crossref |
||||
| Bagenal TB (1967). A Short Review of Fish Fecundity. In: Gerking, SD (Ed.), The Biological Basis of Freshwater Fish Production. Blackwell Scientific, Oxford. pp. 89-111. | ||||
| Bagenal TB, Tesch FW (1978). Age and growth of fish in (Ed. Ricker, WE). Methods for Assessment of fish production in Freshwaters. Blackwell Scientific Publication. Oxford, London P 364. | ||||
|
Baroiller JF, D'Cotta H, Saillant E (2009). Environmental effects on Fish sex determination and differentiation. Sexual Development 3(2-3):118-135. Crossref |
||||
|
Bradford M, Kroon FJ, Russell DJ (2011). The biology and management of Tilapia mariae (Pisces:Cichlidae) as a native and invasive species: a review. Marine and Freshwater Research 62(8):902-917. Crossref |
||||
|
Clausen HJ, Madsen H, Van TP, Dalsgaard A, Murrell KD (2015). Integrated parasite management: path to sustainable control of fishborn trematodes in aquaculture. Trends in Parasitology 31(1):8-15. Crossref |
||||
|
Coward K, Bromage NR (2000). Reproductive physiology of female tilapia broodstock. Reviews in Fish Biology and Fisheries 10(1):1-25. Crossref |
||||
| Dadebo E (2009). Filter feeding habit of the African catfish Clarias gariepinus Burchell, 1822 (Pisces: Clariidae) in Lake Chamo, Ethiopia. Ethiopian Journal of Biological Sciences 8(1):15-30. | ||||
| Duarte F, Araujo FG (2002). Fecundity of the Hoplostomus affinis (Siluriformes Loricariidae) in the Lajes Reservoir, Rio de Janeiro, Brazil. Revised Biology 50(1):197-200. | ||||
|
Emere MC (2000). Parasitic infection of the Nile perch Lates niloticus (L) in River Kaduna. Journal of Aquatic Sciences 15(1):51-54. Crossref |
||||
| Eyo JE, Mgbenka BO (1992). Aspect of the biology of Clarias gariepinus in Anambra river basin I: Oocyte diameter fecundity and sex ratio. Journal of Agriculture Science and Technology 2(I):47-51. | ||||
|
Ezenwaji HMG (2002). The biology of Clarias ebriensis (Pellegrin, 1920) (Osteichthyes: Clariidae) in an Africa rain forest river basin. Fisheries Research 54(2):235-252. Crossref |
||||
| Fawole OO, Adewoye SO (2004). Aspect of the biology of Clarias gariepinus (BURCHELL, 1822) in Oba reservoir, Ogbomoso, Nigeria. Science Focus 8:96-100. | ||||
| Food and Agriculture Organization (1992). Field guide to the freshwater fishes of Tanzania. Food and Agricultural Organization, Rome P 145. | ||||
| Food and Agriculture Organization/World Health Organization (2012). Multi criteria based ranking for risk management of foodborne parasites 2014. (Report of a joint FAO/WHO expert meeting, Rome, 3-7 September, 2012). | ||||
|
Froese R (2006). Cube law, condition factor and weight- length relationships: history, meta-analysis and recommendations. Journal of Applied Ichthyology 22(4):241-253. Crossref |
||||
|
Guerrero-Estévez S, Moreno-Mendoza N (2010). Sexual determination and differentiation in teleost fish. Reviews in Fish Biology and Fisheries 209(1):101-121. Crossref |
||||
|
Hecht T, Endeman F (2007). The impact of parasites, infections and diseases on the development of aquaculture in sub-Saharan Africa. Journal of Applied Ichthyology 14(3-4):213-221. Crossref |
||||
| Hecht T, Uys W, Britz PJ (1982). The culture of sharptooth catfish Clarias gariepinus in Southern Africa. South African National Scientific Programmes Report 153, 133 p. | ||||
|
Hossain MY, Ahmed ZF, Leunda PM, Islam AK, Jasmine MR, Oscoz J, Miranda R, Ohtomi J (2006). Length-weight and length-length relationships of some small indigenous fish species from the Mathabhanga River, Southwestern Bangladesh. Journal of Applied Ichthyology 22:301-303. Crossref |
||||
|
Inyang NM, Ezenwaji HMG (2004). Size, Length-weight Relationship, Reproduction and Trophic Biology of Chrysichthys nigrodigitatus and Chrysichthys auratus (Siluriformes: Bagridae) in a Natural West African Lake. Bio-Research 2(1):47-58. Crossref |
||||
|
Jansen van RC, Jo GV, Asand P, King H (2013). New records of digenean parasites of Clarias gariepinus (Pisces: Clariidae) from the Okavango Delta, Botswana, with description of Thaparotrema botswanensis sp. n. (Plathelminthes: Trematoda). African Invertebrates 54(2):431-446. Crossref |
||||
| Khalil LF (1991). Techniques for identification and investigative helminthology. Helminthology manual. International Institute of Parasitology, St. Albans, UK. P 156. | ||||
| King M (1995). Fisheries biology, assessment and management (Blackwell Science, London. P 341. | ||||
|
Komolafe OO (2004). Reproductive aspects of cichlid fish Tilapia zilli (Gervais) (Pisces: Cichlidae) in Opa reservoir, Ile Ife, Nigeria. Ife Journal of Science 6(2):119-122. Crossref |
||||
|
Komolafe OO (2005). The breeding biology of Oreochromis niloticus linnaeus (pisces: cichlidae) in Opa reservior Ile-Ife, Nigeria. Journal of Science and Technology 25(2):77-83. Crossref |
||||
|
Kuparinen A, Boit A, Valdovinos FS, Lassaux H, Martinez ND (2016). Fishinh - induced life history cxhaanges degrade and destabilize harvested ecosystems. Scientific Reports 16(22245). Crossref |
||||
|
Midhat AE, Kariman AS, Mohammad MNA (2013). Studies of some reproduction characters of Tilapia species in Damietta Branch of the River Nile, Egypt. Journal of Fisheries and Aquatic Science 8:323-339. Crossref |
||||
|
Moravec F (2010). Some aspects of the taxonomy, biology, possible evolution and biogeography of nematodes of the spirurine genus Rhabdochona Railliet, 1916 (Rhabdochonidae, Thelazioidea). Acta Parasitologica 55:144-160. Crossref |
||||
|
Murua H, Saborido-rey F (2003). Female reproductive strategies of marine fish and their classification in the North Atlantic. Journal of Northwest Atlantic Fishery Science 33:23-31. |
||||
| Olaosebikan BD, Raji A (1998). Field Guide to the Nigerian Fresh water Fishes. Federal College of Freshwater Fisheries Technology New Bussa, Nigeria. P 106. | ||||
|
Omeji S, Solomon SG, Idoga ES (2011). A Comparative Study of the Common Protozoan Parasites of Clarias gariepinus from the Wild and Cultured Environments in Benue State, Nigeria. Journal of Parasitology Research 8:41-47. Article ID 916489: Crossref |
||||
|
Oniye SJ, Adebote DA, Ayanda OI (2004). Helminthes parasites of Clarias gariepinus (Tuegels) in Zaria. Nigeria Journal of Aquatic Science 19(2):71-75. Crossref |
||||
| Roberts LS, Janovy J (2000). Gerald D Schmidt and Larry S Robert's foundations of Parasitology. 6th Ed. McGraw-Hill Internatioal editions, Boston. P 382. | ||||
|
Sadovy Y (2001). The threat of fishing to highly fecund fishes. Journal of Fish Biology 59(A):90-108. Crossref |
||||
|
Santi S, Gennotte, V, Toguyeni A, Melard C, Nadine A, Rougeot C (2016). Thermosensitivity of the sex differentiation process in the African catfish, Clarias gariepinus: Determination of the thermosensitive period. Aquaculture 455:73-80. Crossref |
||||
|
Santi S, Rougeot C, Toguyeni A, Gennotte V, Kebe I, Melard C (2017). Temperature preference and sex differentiation in African catfish, Clarias gariepinus. Journal of Experimental Zoology 327A:28-37. Crossref |
||||
|
Sharpe DMT, Hendry AP (2009). Life history change in commercially exploted fish stocks: An analysis of trends across studies. Evolutionary Applications 2:260-275. Crossref |
||||
|
Shoesmith E (1990). A comparison of methods for estimating mean fecundity. Journal of Fish Biology 36:29-37. Crossref |
||||
| Sokal RR, Jamesrohlf F (1987). Introduction to biostatistics. 2nd ed. Freeman and Co. New York. 363 p. | ||||
|
Sotolu AO (2010). Effects of varying dietary protein levels on the breeding performances of Clarias gariepinus brood stocks and fry growth rate. Livestock Research for Rural Development 22(67). |
||||
| Vazzoler AEAM (1996). Biologia da Reprodução de PeixesTeleósteos: Teoria e Prática. Maringá: EDUEM 169 p. | ||||
| Wootton RJ (1992). Ecology of Teleost Fishes. Chapman and Hall, P 404. | ||||
| World Health Organization (2011). Background document for the development of WHO guidline for drinking water quality. 4th Ed. 2011. 564p. | ||||
| Yalcin S, Solak K, Akyurt U (2001). Certain Reproductive Characteristics of the Catfish (Clarias gariepinus Burchell, 1822) Living in the River Asi, Turkey. Turkey Journal of Zoology 25(4):453-460. | ||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


