Full Length Research Paper

ABSTRACT
The benthic zone of Lake Malombe was sampled for invertebrates, fungi and bacteria using an Ekman Grab measuring 15.2 cm by 15.2 cm. Thirty-six stations were surveyed for macro-invertebrates which were identified to the lowest taxa and enumerated to estimate abundance for the lake. Snails were the most dominant macro fauna, belonging to four genera Melanoides, Bellamya, Bulinus and Lanistes with the mean densities of 177.5, 34.7, and 4.3 and 0.1 m-2 individuals, respectively. Blood worms and Tubifex were also present. Although there are few such studies in Malawi, it was generally postulated that dominance of snails is a recent phenomenon following previous studies which showed that the invasion of a form of Melanoides of Asian origin; its success might be responsible for its proliferation. The benthic substrate was mainly composed of mud, clay granules, sand and bedrock. The biomass of macro-fauna is being reported here for the first time and coincides with a decline in fish catches on Lake Malombe. The prevalence of Melanoides species and other high pollution tolerance species suggests that there is high ecosystem modification due to anthropogenic activities including sediment and nutrient loading from agricultural practices in the surrounding area. Compared to Lake Malawi and Upper Shire, Lake Malombe is by far the most productive. There were significantly higher (P<0.5) densities of aerobic, anaerobic bacteria and fungi, demonstrating the importance of the detrital food chain. Therefore, future programs aimed at enhancing fish restoration in Lake Malombe would be advised to include a suite of bottom feeding fish species. The state of benthos found in Lake Malombe is an indication of confounding impacts of over-fishing, climate change and catchment-wide activities. Thus, use of QIIME software could unravel microbiome characteristics, including climate change signatures. Similarly, further studies on food webs could contribute to a better understanding of the Lake Malombe trophic functions.
Key words: Benthic invertebrates, aquatic snails, fungi, bacteria, ecosystems, food chain, Lake Malombe.
INTRODUCTION
Benthic studies have not been frequently conducted in conjunction with fishery related sciences, because there is little appreciation of the role benthic of organisms play in the sustenance of fisheries stocks. Scientists and ecologists who constructed ecosystem models long realised the important role the benthic zone plays in recycling of nutrients and in the provision of food for the bottom dwelling flora and fauna (Christensen and Pauly, 1993; Strayer, 2009). The heterotrophic food chain contributes to primary and secondary production, although energy flowing through the aquatic system is normally modest. However, the same may not have been said about microbial contribution to primary and secondary production. Generally, as aquatic ecosystems develop and mature, the flow and exchange of matter between the aquatic and terrestrial systems becomes more complex (Pfannkuche et al., 1985; Weyl et al., 2010).
Depending on size of organisms passing through sieves, benthic invertebrates can be classified as macro-invertebrates once they pass through 0.5 mm mesh size and are retained by fine sieve of 0.05 mm mesh as meiobenthos if they pass through fine sieves (Strayer, 2009). Furthermore, the zoobenthos biomass usually includes macro-invertebrates and excludes large bivalves. The benthic animals depend on nutrients from phytoplankton, aquatic plants, animals, bacteria and detritus and facilitate biogeochemical cycles (Alexander et al., 2009). It has been reported that where suspension feeders like mussels, sponges and cladocerans are abundant, they reduce phytoplankton biomass and influence its composition (Reeder et al., 1993).
In Malawi, few studies have been devoted to the aquatic benthic habitats. Therefore, such ecosystem changes are not well understood. On the other hand, most of the scientific studies have been focused on the littoral and pelagic zones of Lake Malawi, because of the diversity of fish species and interest from evolutionary scientists. The Lake Malombe fishery has undergone complete cycles of utilization passing through four phases of development from undeveloped, developing, maturation, decline and senescent, hence to understand Lake Malombe’s dynamics, all ecological niches needed to be investigated (FAO, 1993; Matiya and Wakabayashi, 2005; Dulanya et al., 2013; Singini et al., 2013;Hara and Njaya, 2016). This study therefore focused on determining the composition, quantity and contribution of benthic macro-invertebrates and their contribution to the functioning of Lake Malombe ecosystem. Unfortunately, the co-management approach has not produced desired results (Hara et al., 2002), hence fisheries management remain in dire need of fresh efforts.
METHODOLOGY
Study area
The study was carried out in Lake Malombe which is a natural impoundment of the Upper Shire River and is located approximately at 16 km downstream of the outlet of Lake Malawi. It lies between latitudes 14°30′S and 14°45′S and longitudes between 35°12′E and 34°20′E with the surrounding area of the south western part of lake being mostly for agricultural activities while the land-water interface is occupied by emergent aquatic vegetation (Dulanya et al., 2013). The lake is directly fed by waters from Upper Shire River and was known to have a total surface area of approximately 390 to 450 km2. The lake runs 30 km long from and 15 km wide with an average depth of 5 to 7 m, showing steady declining in depth (Jamu et al., 2011; Maguza-Tembo, 2002; FAOa, 2018). The lake is polymictic, where the shallow depths permit complete mixing by wind and frequent recycling of nutrients from the sediments (Donda, 2011; FISH, 2015). Upper Shire River and south east arm of Lake Malawi were also surveyed to compare with the findings from Lake Malombe.
Sample collection and preservation
Field sampling of invertebrates was done in May and October, 2018 using an Ekman Birge grab to have a profile of species that are abundant both during high lake levels and low levels (Strayer, 2009). Part of the soil sample collected at each sampling station was put in sterilized bottle and sealed for microbial analysis and the rest was washed through sieves to separate macro-fauna from sand, silt and other sediments. Larger fauna including, gastropods and beetles were placed in a container while the rest of live aquatic organisms were placed in a separate container. All samples were stored in 70% ethanol to arrest post-mortem degeneration of tissues induced by autolysis and prevent microbial attack. The Garmin Etrex 10 GPS device was used to locate and record the sampling points. Measurements of depth and temperature were taken by a Fish Finder 140 Garmin equipment. Organisms were preserved using alcohol and sorted carefully based on physical characteristics into their lowest taxa possible using a dissection microscope and magnifying lens. All large organisms were placed in a tray and physically observed using a magnifying lens. The smaller organisms were spread onto petri dishes and examined under a dissection microscope.
Media preparation for microbiological analyses
Benthic soil samples were analyzed for total viable counts (TVC), aerobic and anaerobic bacteria, and fungi populations. The soil samples were cultured in an incubator for 18 h. Preparing the media involved adding 28 g of nutrient agar (NA), 15.5 g of malt extra agar and 15.5 g of Nutrient broth which were placed into conical flasks into which 1 L of ionized water was added. The media was then autoclaved for 15 min at 121°C and later cooled. The plates were prepared in duplicates on sterile petri dishes and dilutions of 0.1 mL were obtained by a micropipette and transferred aseptically into the pre-prepared agar plates by raising the upper lid sufficient enough to enter the tips of the pipette. Samples were spread homogenously and aseptically arranged by sterile flamed L-shaped glass rod throughout the surface of the media until the sample dried out.
The nutrient broth media was placed in a Bio-Bag system consisting of an air-tight plastic bag in which petri plates were sealed. A disposable hydrogen (H2) gas container was made to react with oxygen (O2) to form water in sealed bag; eventually, all oxygen in the bag was consumed. This created an anaerobic environment. This was followed with autoclaving the medium to drive off dissolved oxygen. Thereafter, the shake culture technique was used and a tube of Fluid Thioglycollate (FTG) medium held at 48°C was inoculated before the medium solidified. FTG medium contained a small amount of agar to increase viscosity and reduce diffusion of oxygen into medium. Finally, an indicator which turned colourless in the absence of oxygen was included to confirm that anaerobic conditions were achieved.
Soil sample inoculation into the media
For each sample, a subsample of 1 g of soil was transferred to a sample vial containing 9 ml of 0.1% sterile peptone water. The vial was closed, shaken thoroughly for 10 min and allowed to stand for 20 min. Thereafter, a 6-fold serial dilution was carried out in triplicates. The inoculated plates were used for Total Viable Counts (aerobic and anaerobic bacterial counts) after incubation at 28°C. The colony-forming units (CFU/g) were counted under a Quebec dark field colony counter (Leica, Buffalo. NY. USA) equipped with a guide plate ruled in square centimetres. Results were recorded as Colony Forming Units per gram (CFU/g).
Statistical analysis
Data were processed by Microsoft Excel. Mean distribution and abundance of micro-invertebrates were compared by One Way Analysis of Variance (ANOVA) at 5% level of significance using SPSS software version 16.0. Significantly different treatment means were separated using the Least Significant Difference. The Shannon index was used to calculate the diversity using the equation:
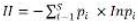
where H = the Shannon diversity index, Pi = fraction of the entire population made up of species I, S = numbers of species encountered and ∑ = sum from species 1 to species S.
RESULTS
Benthic organisms in Lake Malombe
Data analysis from 35 sampling station in Lake Malombe revealed that snails dominate the zoobenthic invertebrates in the Lake and is shown in Table 1. This was followed by Diptera larvae, annelids and black fly larvae.
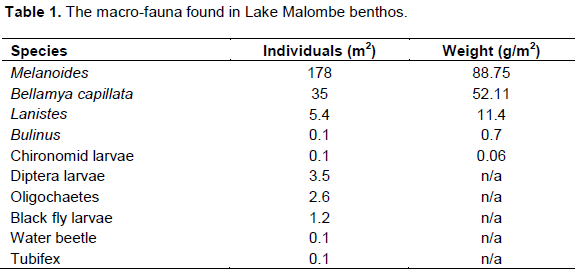
Overall, Melanoides registered average density of 177 m-2 followed by Bellamya capillata 35 m-2. Although B. capillata (Figure 1) are larger in size (almost 4 times) as compared to Melanoides, their relatively smaller numbers were outnumbered by the smaller forms of Melanoides.

Other invertebrates including tubifex, water beetles and chironomids larvae, were especially present in water depths ranging from 2.5 to 3.5 m. These macro-invertebrates were mostly found in muddy bottom substrates.
Spatial distribution of snails in Lake Malombe
Geo-referencing results of the sampled stations showed that the snails Melanoides which dominated the benthic zones of Lake Malombe were more abundant in the north-western part of the lake (Figure 2).
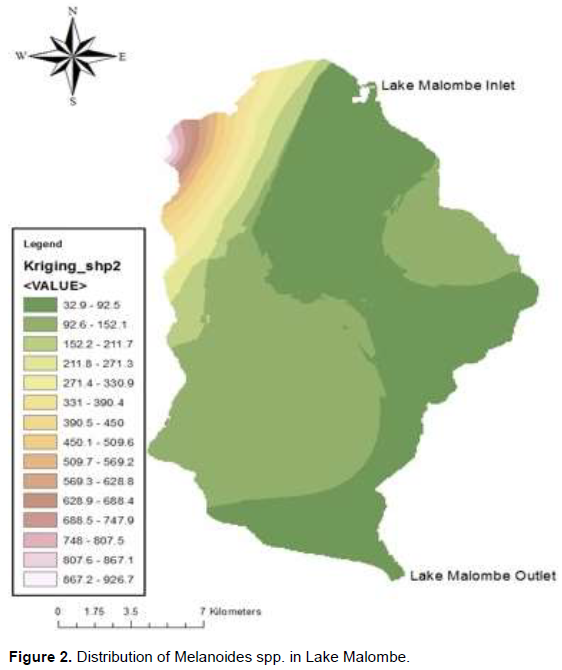
In contrast, the distribution of B. capillata in Lake Malombe showed that the snail was highly concentrated in the south western part of the lake (Figure 3).
Substrate types and composition in Lake Malombe
The hard clay substrates were common in deeper waters (>3 m). These areas contained less live benthic fauna and debris but contained considerable amounts of decaying snail shells. The composite substrate had a unique light greyish colour with very tiny particles making it very smooth to tough but not mouldable. The substrate was composed of weathered and decayed shells of snails and fine particles of clay soils (Figure 4).

Snails in Lake Malawi, Malombe and Upper Shire River
A comparative analysis of Lake Malombe, Upper-shire River and Lake Malawi benthic macro-invertebrates at depths ranging from 1.5 to 3.5 m showed remarkable differences in snail distribution and concentration. In Lake Malombe, the snail of the genus Melanoides are dominant making up 85% of the benthic macro-fauna (Figure 5).
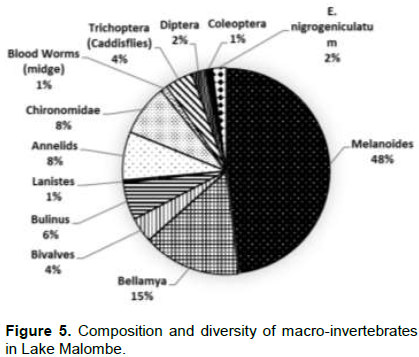
The B. capillata was found to contribute 12% while chironomids larvae contributed only 1%. Thus, Lake Malombe showed great disparities of benthic fauna distribution and abundance.
The abundance of Melanoides in the Upper-Shire River was estimated at 48%, showing a 50% difference with Lake Malombe. B. capillata abundance is slightly higher than 15% compared to Lake Malombe at 12%. The Upper Shire River had a more diverse number of benthic fauna which included nymph stages of species of the order Coleoptera, Diptera and Trichoptera (Figure 6).
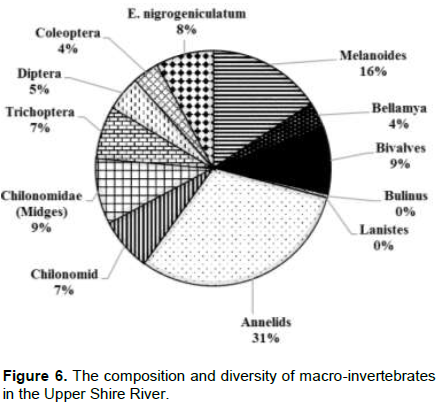
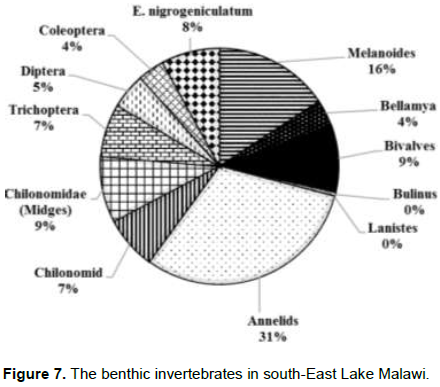
There was also an increase in the abundance of snails of the genus Bulinus and Annelids. The abundance of both Bulinus and Annelids in Upper-Shire River was 8% as compared to 1% in Lake Malombe. Results from Lake Malawi showed that the measures of abundance of Annelids, Melanoides, Bellamya, Chironomids larvae and bivalves were 31, 16, 4, 9 and 4%, respectively. In Lake Malawi, dominance is displayed Annelids and the community is more diverse (Figure 7).
Species richness in south east arm of Lake Malawi, Lake Malombe and Upper Shire River
Although the Upper Shire and southeast Lake Malawi were not as intensively sampled as Lake Malombe, comparisons can be drawn to aid future studies.
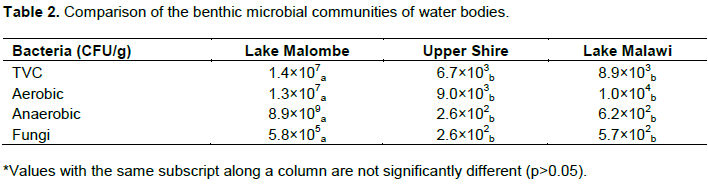
The Shannon index of species richness (Table 2) showed that the order of importance was Lake Malawi (1.64), Upper-Shire River (1.46) and Lake Malombe (0.64), respectively. The species richness of Lake Malombe were statistically different to that of Lake Malawi and Upper-Shire River (P<0.05).
Microbial population of benthic soils for the three lake sytems
Table 2 shows a summary of microbiological analyses results for the three ecosystems. Lake Malombe had significantly higher numbers of total plate count of 1.4×107 CFU/g compared to Upper Shire and part of Lake Malawi which had the same order of magnitude; 6.7×103 and 8.9×103 CFU/g, respectively; these results were significantly different (P=0.001). Significantly higher populations (P=0.01) of fungi, aerobic and anaerobic bacteria were identified in Lake Malombe as compared to Upper Shire and Lake Malawi. The difference in microbiological populations of the benthic soil samples reveals productivity and comparative importance of the heterophic food chain of the three ecosystems.
DISCUSSION
Abundance and distribution of benthic macro-invertebrates in Lake Malombe
The macrobenthos fauna of Lake Malombe showed little diversity as compared to the Upper-Shire River or Lake Malawi. The most common gastropods in Lake Malombe belonged to the genus Melanoides, which are believed to be very tolerant to diverse environments, according to Berry and Kadri (1973) and Vogler et al. (2012). This genus has been found to occupy a variety of habitats, ranging from weakly saline close to sea-level to freshwater of higher altitudes and can thrive in a wide range of pH and hardness regimes and in distinct kinds of substrates, however, it is most common in the mud. This is in-line with current results where Melanoides were most abundant in muddy substrates. The diets of this snail comprise of several types of algae, bacteria, deposits of organic materials and decaying flora (Pointier et al., 1991; Ben-Ami and Heller, 2005; Raw et al., 2016); hence, the lake provided a perfect environment.
B. capillata was the second most abundant snail in Lake Malombe and is also a widely distributed gastropod in the Africa region. This species is also found in coastal Tanzania, north-eastern Natal, Democratic Republic of Congo and Lake Victoria Basin (Van Damme and Lange, 2016). The distribution of B. capillata is influenced by physico-chemical parameters notably water temperature and salinity, whose extreme values shows an inverse effect regarding distribution and abundance (Brown, 1994). Freshwater gastropods are regarded as bio-indicators of water quality and play a vital role in purifying water since they are saprophytic, obtaining nourishment from dead and decaying organic matter. The other gastropods like Bulinus are of great medical importance for being intermediate hosts of infectious trematodes and other parasites of animals and human beings (Brown, 1994). One of the most important parameters being calcium salts are found in both food and influence the growth of gastropods (Dillon, 2000). The benthic ecosystem generally favours the growth and reproduction of gastropods, therefore, the success of gastropods to colonize and dominate Lake Malombe benthos can also be attributed to overfishing. Snail eating fish species such as Trematocranus placodon and Synodontis njassae have been over-fished in the lake, thereby removing the natural biological control (FAO, 2018c).
The presence of Tubifex worms and blood worms in Lake Malombe benthic ecosystem confirms the assumption made that deeper parts of the lake (>3 m) are eutrophic and are generally of low oxygen content. According to Pelegri and Blackburn (1995), Tubifex worms along with chironomids larvae are the most abundant macro fauna species in eutrophic lake sediments that occur in Lake Malombe. Tubifex worms feed in the ranges of 2 to 8 cm of the benthic sediments, primarily consuming microfauna composed of bacterial and fungi.
The benthic fauna of southeast arm of Lake Malawi and Upper Shire River
The benthic fauna of these two ecosystems were not significantly different; however, both were lower than in Lake Malombe. In Upper Shire River, Melanoides were dominant at 48% and Bellamya came second at 15%. In Lake Malawi, the benthic fauna was observed to be dominated by Annelids (31%) seconded by Melanoides (16%). The density of snails and other aquatic organisms in Lake Malawi and Upper Shire River was high in the littoral zone as compared to the middle of the lake or deeper waters. However, Melanoides were present in all the sampling stations, this confirms what Genner et al. (2004) had previously observed that Melanoides species showed rapid reproductive potential and spread to littoral habitats.
The reduction of the benthic fauna towards the middle of the two ecosystems (Lake Malawi and Upper Shire River) may be explained by the increase in silt content, indicating a reduction of suitable space. Martin et al. (1998) showed that the oxygenated layer in the sediment is reduced to the few first millimetres in shallow water and <1 mm near the anoxic layer. A limnology study conducted in the same Lake showed that dissolved oxygen levels did not exceed 7 mg/L throughout the Lake (FAO, 2018b). It has been observed elsewhere that the longer periods of very low oxygen content or anoxic conditions may prevent survival of aerobic species that are tolerant to low oxygen (Ott and Schiemer, 1973). Another probable reason for the density decrease of benthic invertebrates may be reduction of food availability. In deep water, benthic standing stock usually reflects food availability (Pfannkuche, 1993). In addition, there was relatively high water velocity at the middle of the lake as well as in the river (Giovanelli et al., 2005; Buss et al., 2004; Kloos et al., 2001).
It was observed that the benthic substrate of Lake Malawi had several submerged aquatic plants, which were scarce in Upper Shire River and especially at the area close to Lake Malombe. Most fishers use beach seine nets which drag the bottom, especially the “Nkacha” nets which are illegalised in Lake Malawi. This is likely to have affected growth of submerged aquatic plants in Upper Shire River contributing to reduction of benthic fauna that depend on aquatic vegetation for food, shelter and breeding (FISH, 2015). This association needs further investigation.
The role of microbial population in the ecosystems
The total viable counts for bacteria and fungi in Lake Malombe were shown to be higher than Upper Shire River and Lake Malombe. This suggests that benthic bacteria are essential food for invertebrates forming the base of the heterotrophic food chain in these systems. Regarding their trophic role, bacteria would be expected to play a vital role in mineralization of detritus, dead shells of snails and the recycling of essential nutrients. The microbial populations in the aquatic systems serve as a general indicator of nutrient recycling and health of the benthic system (Castelli et al., 2004; Weyl et al. 2010). Aerobic and anaerobic microbial populations in Lake Malombe were shown to be more abundant as compared to Lake Malawi and Upper Shire benthic systems. The slow movement of water in Lake Malombe, essentially a floodplain lake, meant that bacteria have sufï¬cient time to establish themselves and to allow interactions that support multifunctional activities (Gamfeldt et al., 2008). The aerobic bacteria and fungi consume dissolved oxygen and leave anaerobic zones where the anaerobic bacteria conduct metabolic processes. Steele et al. (2001) reported that these bacteria constitute the foundation of all earth’s ecosystems, being responsible for the degradation and recycling of essential elements such as carbon, nitrogen and phosphorus. Alongi (1994) reported that the energetic power of microbes is greater than all other living organisms combined and is the powerhouse driving aquatic food webs. Fungi are saprobes and obtain substrates from the remnants of dead plants and animals in soil or other aquatic habitats (Sergeeva and Kopytina, 2014). Therefore, higher counts in the Lake Malombe benthos indicate a lot of decomposition of detritus and dead shells; this is evidence of high nutrient recycling as well as effects of anthropogenic inputs from catchment area of Lake Malombe. This was also evident in high concentrations of carbonates and bicarbonates (FAO, 2018b), which could be signature nutrients for the lake. Therefore, the biological processes in the benthic soils are exerting substantial effect on the aquatic environment of Lake Malombe through the heterotrophic food chain.
Evidence of role of substrate in the ecosystems
The substrate composition has been reported to be very closely linked to ecosystem functions, including microbial communities as they harbour microbial populations (Ylla et al., 2013). Lake Malombe is a rich source of established benthic communities which have been developed on the appropriate substrate (Gamfeldt et al., 2008). The study found that clay and composite substrates were highly prevalent in Lake Malombe, while sand, silt and fine sand were present in Upper Shire River. Only a small portion of Lake Malawi was sampled and had silt, mud and clay.
CONCLUSION AND RECOMMENDATIONS
Having demonstrated that the benthic community of Lake Malombe is potentially a rich source of a suite of food including bacteria, fungi and snails; finding fish species which will feed on these organisms should be considered for stock enhancement programs that will contribute to effective fisheries management. To further advance an understanding of Lake Malombe, studies using QIIME software are required to sequence the microbes and couple with climate change parameters including signature carbon species.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Alexander E, Stock A, Breiner HW, Behnke A, Bunge J, Yakimov MM (2009). Microbial eukaryotes in the hypersaline anoxic L'Atalante deep-sea basin. Journal of Environmental Microbiology 11:360-381. |
|
|
Alongi D (1994). The role of bacteria in nutrient recycling in tropical mangrove and other coastal benthic ecosystems. Hydrobiologia 285:19-32. |
|
|
Ben-Ami F, Heller J (2005). Spatial and temporal patterns of parthenogenesis and parasitism in the freshwater snail Melanoides tuberculata. Journal of Evolutionary Biology 18:138-146. |
|
|
Berry AJ, Kadri ABH (1973). Reproduction in Malayan Freshwater Cerithiacean Gastropod Melanoides tuberculata. Journal of Zoology 172(3):369-381. |
|
|
Brown SD (1994). Freshwater snails of Africa and Their Medical Importance. Taylor and Francis, United Kingdom. |
|
|
Buss DF, Baptista DF, Nesimian JL, Egler M (2004). Substrate specificity, environmental degradation and disturbance structuring macroinvertebrate assemblages in Neotropical streams. Hydrobiologia 518:179-188. |
|
|
Castelli A, Lardicci C, Tagliapietra D (2004). Soft-bottom macrobenthos. In: Gambi MC, Dappiano M (Eds). Biologia Marina Mediterranea, Mediterranean Marine Benthos: A Manual of methods for its sampling and study. S.I.M.B. Onlus, Genova, pp. 99-131. |
|
|
Christensen V, Pauly D (1993). Trophic Models of Aquatic Ecosystems. ICLARM Conference Proceedings 26:390-396. |
|
|
Damme D, Lange C (2016). Bellamya capillata. The IUCN Red List of Threatened Species. |
|
|
Dillon RT (2000). The Ecology of Freshwater Molluscs. Cambridge University Press. London. |
|
|
Donda SJ (2011). Fishing is our gold mine: lessons learned from participatory fisheries management in Malawi. World Small-scale Fisheries: Contemporary Visions pp.235-247. |
|
|
Dulanya Z, Reed JM, Trauth MH (2013). Mapping Changing Shorelines in the Malombe and Chiuta Lakes of Malawi-Environmental Effects of Recent Climatic Variations. Catena 104:111-119. |
|
|
Food and Agriculture Organization (FAO) (2018a). Report on the Bathymetric Survey of Lake Malombe prepared by Gift Chigona and Orton V. Msiska. FAO, Lilongwe, Malawi. 15 p. |
|
|
Food and Agriculture Organization (FAO) (2018b). A Study of Limnology of Lake Malombe, Malawi prepared by Yamikani Balaka, Hermis Chagoma, Titus Phiri and Orton Msiska. 35 p. FAO. Lilongwe, Malawi. |
|
|
Food and Agriculture Organization (FAO) (2018c). Fisheries Assessment Studies of Lake Malombe, 2017. Compiled by Orton V. Msiska, Lilongwe, Malawi, 67 p. |
|
|
Food and Agriculture Organization (FAO) (1993). Fisheries management in the south-east arm of Lake Malawi, the Upper Shire River and Lake Malombe, with particular reference to the fisheries on Chambo (Oreochromis spp.). CIFA Technical Paper. No. 21. Rome 113 p. |
|
|
FISH (2015). Environmental Threats and Opportunities Assessment (ETOA) of Four Major Lakes in Malawi. USAID/FISH Project, Pact Publication, Lilongwe, Malawi: 250 p. |
|
|
Gamfeldt L, Hillebrand H, Jonsson PR (2008). Multiple functions increase the importance of biodiversity for overall ecosystem functioning. Ecology 89:1223-1231. |
|
|
Genner MJ, Michel E, Erpenbeck D, De Voogd N, Witte F, Pointier JP (2004). Camouflaged invasion of Lake Malawi by an Oriental gastropod. Molecular Ecology 13:2135-2141. |
|
|
Giovanelli A, Coelho da Silva CLPA, Leal GBE, Baptista DF (2005). Habitat preference of freshwater snails in relation to environmental factors and the presence of the competitor snail Melanoides tuberculatus (Müller, 1774). Memórias do Instituto Oswaldo Cruz 100(2):169-176. |
|
|
Hara MM, Donda SJ, Njaya F (2002). An evaluation of the Lake Malombe Co-management Programme. Research Report. Department of Fisheries, Lilongwe, Malawi. |
|
|
Hara M, Njaya F (2016). Between a rock and a hard place: The need for and challenges to implementation of Rights Based Fisheries Management in small-scale fisheries of southern Lake Malawi. Fisheries Research 174:10-18. |
|
|
Jamu D, Andrew NL, Bootsma HA, Hecky RE (2011). Editorial: Reconciling livelihoods and aquatic ecosystem resilience in the lakes of Malawi. Journal of Great Lakes Research 37:1-2. |
|
|
Kloos H, Souza C, Gazzinelli A, Filho BSS, Temba PC, Bethony J, Page K, Grzywacz C, Lewis F, Minchella D, LoVerde P, Oliveira RC (2001). The distribution of Biomphalaria spp. in different habitats in relation to physical, biological, water contact and cognitive factors in a rural area in Minas Gerais, Brazil. Memórias do Instituto Oswaldo Cruz 96:57-66. |
|
|
Maguza-Tembo F (2002). Bio-economics of common resource over exploitation. Case of Lake Malombe Chambo (Oreochromis sp. Cichlidae) fishery in Malawi. MSc., Economics Department, Norwegian College of Fishery Science, University of Tromso, Tromso, Norway 51 p. |
|
|
Matiya G, Wakabayashi Y (2005). Small Scale Fisheries of Malawi. An outline of Lake Malombe fisheries. Mem. Faculty of Agriculture, Ehime University 50:3-9. |
|
|
Ott JA, Schiemer FS (1973). Respiration and anaerobiosis of free-living nematodes from marine and limnic sediments. Netherlands Journal of Sea Resources 7:233-243. |
|
|
Pelegri SP, Blackburn TH (1995). Effects of Tubifex (Oligochaete: Tubifex) on N-Mineralization in Freshwater Sediments, Measured 15N Isotopes. Journal of Aquatic Microbe Ecology 9:289-294. |
|
|
Pfannkuche O (1985). The Deep-Sea Meiofauna of the Porcupine Seabight and Abyssal Plain (North East Atlantic): Population Structure, Distribution, Standing Stocks. Acta Oceanologica Sinica 8(3):343-353. |
|
|
Pfannkuche O (1993). Benthic standing stock and metabolic activity in the bathyal Red Sea. Marine Ecology 14(1):67-79. |
|
|
Pointier JP, Toffart JL, Lefevre M (1991). Life tables of freshwater snails of the genus, Biomphalaria-alexandrina, Biomphalaría-straminea and one of its competitors Melanoides tuberculata under laboratory conditions. Malacologia 33(1-2):43-54. |
|
|
Raw JL, Perissinotto R, Miranda NAF, Peer N (2016). Feeding dynamics of Melanoides tuberculate (Müller, 1774). Journal of Molluscan Studies 82(2):328-335. |
|
|
Reeder HA, Bijde VR, Noordhuis R (1993). Potential of the zebra mussel (Dreissena polymorpha) for water quality management. In: Nalepa TE, Schloesser DW (Eds.). Zebra Mussels: Biology, Impacts and Control. pp. 439-451. |
|
|
Sergeeva NG, Kopytina NI (2014). The first marine filamentous fungi discovered in the bottom sediments of the oxic/anoxic interface and in the bathyal zone of the Black Sea. Turkish Journal of Fisheries and Aquatic Science 14:497-505. |
|
|
Singini W, Kaunda E, Kasulo V, Jere W (2013). Wealth Based Fisheries Management of Chambo (Oreochromis spp.) Fish Stock of Lake Malombe in Malawi. International Journal of Fisheries and Aquaculture 5(11):270-277. |
|
|
Steele JH, Thorpe SA, Turekian KK (2001). Encyclopaedia of Ocean Sciences. Academic Press 6:748-749. |
|
|
Strayer DL (2009). Benthic invertebrate fauna, lake and reservoirs. In: Gene EL (Ed.). Lake Ecosystem Ecology. Encyclopedia of Inland Waters pp. 27-40. |
|
|
Vogler RE, Nú-ez V, Gutiérrez Gregoric DE, Beltramino AA, Peso JG (2012). Melanoides tuberculata: The history of an invader. In: Hämäläinen EM, Järvinen S (Eds.). Chapter 3 Snails: Biology, Ecology and Conservation pp. 65-85. |
|
|
Weyl OLF, Ribbink AJ, Tweddle D (2010). Lake Malawi: fishes, fisheries, biodiversity, health and habitat. Aquatic Ecosystem Health and Management 13(3):241-254. |
|
|
Ylla I, Peter H, Romaní AM, Tranvik LJ (2013). Different diversity–functioning relationship in lake and stream bacterial communities. FEMS Microbiology Ecology 85:95-103. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


