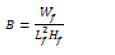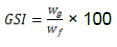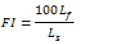ABSTRACT
Although artisanal commercial and net fishers landings has received attention, the morphometrics of bait fisher landings is poorly documented. This is in spite of the importance of such data to infer fished stock growth and developmental status in order to reduce excessive removal of immature and highly fecund fish. This study compares fish species, size and weight, and derived fish condition, Gonadosomatic and Fishing Indices, between hook and line gear baited with polychaetes and hermit crabs, and traps baited with gastropods. Results indicate fishers land over 20 fish types with similar condition and GSI below unity and fishing index of below 20%. Variation in growth indicators was largely attributed to species specific differences, with high value apex predators, such as carangidae, having lower morphometrics than smaller bodied permanent creek and mangrove residents. Participatory effort controls through closure, alternative livelihoods, and voluntary release of immature, coupled with limiting the use of small mesh traps and hooks, may reduce harvesting of immature and lead to improvement of future landings and sustainability of the fishery.
Key words: Bait, condition factor, gear, Gonadosomatic index, fishing index, immature.
Tropical artisanal fishers contribute to significant amount of global fisher landings (Cheunpagdee et al, 2006). Along the Kenyan coastline, artisanal fishers land 90% of marine fisheries landings (8 Mt), with hook and line and basket trap fishers forming 40 to 60% of fishers (Fondo, 2004; Frame Survey, 2012). Tropical artisanal fishers use a variety of gears to target multispecies assemblage of fish using unmotorised crafts (Van der Elst et al., 2005). Among gears used, a variety of nets (e.g. seines, cast nets etc), hooks (e.g. handline, long-line) and traps (e.g. basket traps, weirs, fence traps), have been mentioned in literature. While the morphometrics of fish landed using nets by artisanal fishers along the Kenyan coast (Kaunda-Arara et al., 2003; Agembe et al., 2010; Mbaru et al., 2010; Gajdzik et al., 2014) and elsewhere, have been described, those landed by bait fishers (hook and trap fishers), have received less attention and hence the current study.
In temperate regions, angler fisher landings using baited hooks (Arlinghaus et al., 2008) and traps (e.g. Garrison et al., 2004; Fisher and Fisher, 2006; Alos et al., 2009), are available, similar information is scanty for tropical bait fishers. For instance, description of tropical artisanal bait fishery is limited to the mention of unidentified baits, such as worms, squid, octopus and small fish, for hook fishers, and seaweeds, urchin, mollusks and octopus for trap fishers with limited data on the fish they target (Samoilys et al., 2011; Elders of Atafu Atoll, 2012). Tropical baited trap fish landings in general is described for instance in Wambiji et al. (2008), Harnish et al. (2004) and also Garrison et al. (2004), however from these studies, bait use is inadequately described.
Tropical bait fishers use a variety of bait, attached to hooks or inserted into traps to target nearshore fish, prevalent in coral or mangrove fringed shores and lagoons (Glaesel, 1997; Samoilys et al., 2011). Currently, net mesh size restrictions are enforced along the East African coastline, primarily to reduce off-take of juvenile immature fish (Samoilys et al., 2011), but the catch and selectivity of bait and baited gears are less known and consequently unregulated.
Fish size and weight, are commonly used morphometrics used to derive information on fisher induced mortality, and hence the status of a fishery. Condition factor (k) is a derived morphometric indicator that infers the health of a fishery, with high k associated with a fishery specializing in large, healthy, and plump fish stocks (Froese, 2006; Richter et al., 2000). Gonadosomatic index (GSI) on the other hand, is an indicator that reflects the fertility, spawning and fecundity of the fish (Nandikeswan and Anandan, 2013). Fish condition and GSI have been shown to vary with season, diet and fish shape (Nash et al., 2006). Condition factor of 0.2 to 1.2 have been recorded with values close to or above unity, representing fish with superior condition (Agembe et al., 2010; Al-Zibdah and Kan’an, 2009; Mbaru et al., 2010). Depending on fish species and sex, GSI cutoff of between 1 to 10% have been suggested to indicate mature adults (Al-Zibdad and Kan’an, 2009; Kreiner et al., 2001; Zeyl et al., 2013). Seasonal variation in nutrition and breeding patterns affect both condition factor and gonadal maturity (Calderone et al., 2012; Daliri et al., 2012). Mangrove fringed creeks are closely associated with breeding fish and hence the need for their protection (Krumme and Saint-Paul, 2010).
An ideal fishery should therefore have a high condition factor and relatively low GSI. In fisheries with poor morphometric indicators, it is essential to demonstrate the potential harm to the resource and ecosystem to both managers and users prior to implementing changes. For that matter, size at first maturity of targeted fish is also an important parameter that informs resource managers whether the fishery target sub-adults or mature fish. A suitable index (here referred to as Fishing Index) can be derived by comparing morphometrics of landings with standard adult dimension documented elsewhere (Fishbase, 2011) and measuring departures.
This study evaluates morphometrics of bait fisher landings by comparing size, species and derived indices such as condition factor, gonadosomatic and fishing index of fish landed using basket traps baited with tondo (Terebralia palustris- mangrove whelk), and hooks baited with choo (Marphysa sp) and dophe (Clibenarius sp- hermit crab). The underlying question addressed is whether the fishers are employing the different baits, land fish with similar characteristics. Information derived from this study may thus be useful to resource managers to identify and design targeted intervention for unsustainable bait fishery. The study site adopted here, Mida creek, occurs within a Marine Biosphere reserve, where commercial fishery exploitation is restricted (Tuda and Omar, 2012) and hence is suitable for evaluating artisanal fisheries patterns and impacts.
Study area
Mida creek (03° 21’S; 39° 59’E) is a Marine National Park and World Biosphere Reserve situated 88 km North of Mombasa town, that has been in existence since 1968 (Figure 1). The creek covers an area of 31.6 km2, consisting of mangrove forests, seagrass beds, sandflats, rocky outcrops and subtidal habitats.
The communities living around the creek (a population of 10,000), are actively involved in fishing, subsistence farming, small scale businesses and tourism (Mwaipopo et al., 2011). A 2012 fisheries survey estimates between 250 and 500 registered fishers, dependent on the Mida creek ecosystem (Frame Survey, 2012). The Mijikenda and Bajuni tribal groups dominate the fisher community (Hoorweg et al., 2006). Additionally, migrant seasonal fishers from Pemba (Tanzania) also participate in fisheries at Mida during the Northeast monsoon period (October to January).
Trap fishing at Mida
The hexagonal basket traps (malema) used (2×3 m at widest part × 0.5 m height), are woven using local fibers, and have funnel mouth opening of 6 to 8 cm and mesh openings of 3 to 5 cm on the sides. The traps are baited using crushed tondo (T. palustris- Gastropoda; mangrove whelk), collected from the adjacent mangrove forest floor. Although other species are used elsewhere by trap fishers, at Mida, tondo are preferred. Fishers gather 200 to 300 tondo from within the mangrove forest. Trap fishers insert 50 to 100 gastropods within the traps. Each trap fisher owns and operates between three and five traps. The traps are weighted with coral rock and retrieved after overnight soaking, within creeks in the mangrove forest.
Hook fishing at Mida
Bait used by hook fishers is either choo (Marphysa sp- polychaeta) or dophe (Clibanarius sp-crustacea; hermit crab) gathered and prepared prior to the fishing. Although other bait types may be used
at Mida, and elsewhere, hook fishers at Mida prefer choo and dophe. The local names of the bait, that is, choo- Marphysa and dophe- Clibanarius, will be used interchangeably in subsequent discussions. The choo are extracted at the seaward edge of the mangrove forest by digging and extraction from the sandy-muddy substrate and placed in suitable containers for subsequent fishing. On the other hand, the dophe are gathered by searching within the mangrove forest floor and collected for processing. The hermit crab shells are crashed and the crab within removed. The abdomen of the crab is cut off and subsequently used as the bait for fishing.
Hook fishers attach appropriate bait to one or more hooks of size 8 to 18’, and to a line, sinker and Styrofoam float. Hook fishing occurs from a dugout canoe and the line is reeled in by hand, and catch placed at the bottom of the canoe.
Fish sample collection
Fish specimen were collected from fishermen during two sampling occasions in November 2013 and February 2014. The North Eastern Monsoon period that runs from November to February represents the main marine fishing season in Kenya (Tuda et al., 2008). Participating fishers were identified at fishing villages at Mida: Kirepwe, Mayonda and Dabaso, and monitored daily over a period of 10 days during each sampling occasion. Approximately 140 fishers were monitored (~10 per day × 7 days). The types of bait used, as well as the fish landed, were evaluated. In situation where fisher landing was less than 1 kg the whole haul was taken, however, when landing was larger subsampling was done by sorting the fish into species and size categories and 10% of the fish samples taken. The fish specimen obtained were appropriately labeled and immediately preserved in ice for transportation and subsequent refrigeration. In the laboratory, preserved specimen were thawed, identified according to Fishbase (2011) and Richmond (2011) and used in morphometric analysis. Fish species occurring more than three (n=3) times were included in subsequent morphometric analysis.
Determination of morphometric parameters
Condition factor
The standard length of fish was determined using a measuring board, as distance from snout to the tip of caudal fin, while height was taken as distance at widest part of the fish specimen. Excess moisture was removed prior to weight determination, with a sensitive balance. Precision for the size and weight determined were 0.1 cm and 0.01 g respectively. Data obtained was used to calculate condition factor (B) using the modified Foulton’s condition factor, as used by Richter et al. (2000):
Where B- condition factor, Wf- weight of fish, and Lf- standard length of fish and Hf- height of fish. It was assumed that fish had isometric growth due to correction by inclusion of height in the original Foulton condition factor equation as described by Richter et al. (2000). Condition factor data obtained were compared between gear and bait.
Gonadosomatic index
The gonad of each fish was dissected out and the weight determined. Data obtained was used to calculate the Gonado Somatic Index (GSI) using the following equation:
Where GSI- Gonadosomatic Index, Wg- Weight of gonad, Wf –Weight of the fish.
Data obtained was compared between different gear and bait used by the fishers.
Fishing index
Fishing impact was assessed using a fishing index (FI):
Where FI- fishing index, Lf- size of landed fish, Ls- size of adult.
Adult sizes of landed fish were obtained from Fishbase (2011). Sizes at first maturity were adopted, whenever possible, and where absent, common size was used. Where multiple records of size at first maturity for a species were indicated in Fishbase (2011), the lower value was adopted. Data on fishing index obtained were compared between gear and bait.
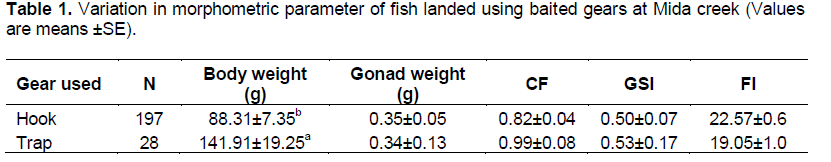
Fishers at Mida landed fish with an average body weight of 115.11±10.30 g and an average gonad weight of 0.35±0.07 g corresponding to a Gonadosomatic Index (GSI) of 0.51. The mean condition factor (CF) for fish landed at Mida was 0.87±0.04, while the average fishing index (FI) for landed the fish was 21.4±0.5.
Variation in fish landings with gear employed
Although trap fishers landed higher weight of fish (ANOVA F=6.77, df=1, P<0.05) than hook fishers, condition factor (χ2=184, df-179, P=0.4), GSI (χ2=80.4, df=85, P= 0.6), and fishing index (χ2=91.9, df-84, P=0.3), were similar (Table 1). Trap fishers landed fish with a condition and GSI slightly higher than hook fishers, but lower Fishing index than hook fishers (Table 1).
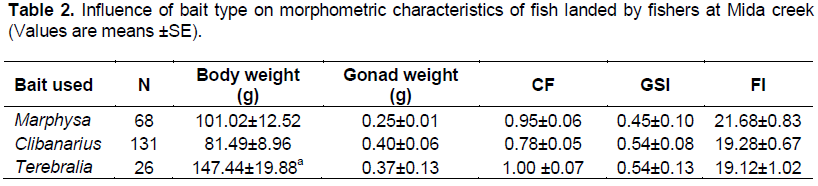
Variation in fish landings with bait types
Three types of bait; Marphysa, Clibanarius and Terebralia used by Mida fishers were tested. Significant differences in weight of fish landed were detected among the bait used with Terebralia landing the heaviest fish (147.44±19.88 g), while Clibanarius the lightest fish (81.49±8.96 g) (Table 2). However, condition factor (χ2=363, df-358, P=0.4), GSI (χ2=165.4, df-170, P=0.6) and fishing index (χ2=177.7, df-168, P=0.3), were not significantly different among the bait types (Table 2). The lowest condition factor was from Clibanarius fishers (0.78), while the highest for Terebralia fishers (1.00). On the other hand, Clibanarius (0.54) and Terebralia (0.54) fishers had higher GSI. In contrast, Marphysa fishers had the highest FI of 21.7.
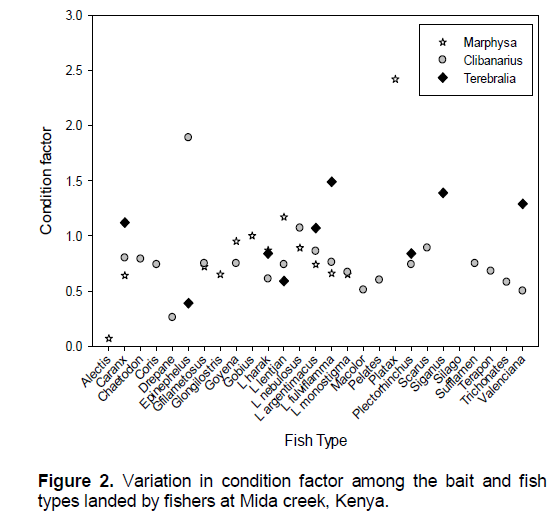
Variation in condition factor among the fish species and bait types
Comparison of condition factor among the species revealed no statistical difference (χ2=4995, df-4945, P=0.3). The highest condition factor was reported for Platax (2.4), Silago (1.39), Epinephelus (1.14) and Gobius (1.0), while Drepane (0.26) had the lowest (Figure 2).
Apart from Epinephelus, Lethrinus harak, Lethrinus lentjan and Plectorhinchus, most species landed using Terebralia had condition factors of above 1.0, with the highest being Siganus sutor. Marphysa fishers had only 3 fish types; Platax (2.42), Gobius (1.0) and L. lentjan (1.17) with condition factors above unity, however, other species condition factor rarely fell below 0.6. Among Clibanarius fishers, only two species Epinephelus (1.89) and Lethrinus nebulosus (1.07) had condition factor above unity, while the lowest was Drepane (0.26).
Comparison of GSI among the species and bait types
Comparison of GSI among the species indicate no significant differences (χ2=2792, df-2795, p<0.0001). Less than 20% of the fish species landed had a GSI above unity (Figure 3). The highest GSI was recorded Drepane punctata (2.08), L. nebulosus (1.66), L. lentjan (1.39), Sufflamen (1.37), Pelates (1.35) which recorded GSI above unity. On the other hand, S. sutor, Silago sihama, Terapon jarbua, Coris, Gerres oyena, Lethrinus argentimacus, Platax and Alectis had the lowest GSI of below 0.1. Other species such as Caranx ignobilis, Epinephelus, Gerres filamentosus, Gerres longilostris, L. harak, Lutjanus fulviflamma, Lutjanus monostigma, Trichonates and Valenciana had between 0.1 to 0.5. Chaetodon, Gobius, Lutjanus argentimacus, Macolor niger, Plectorhinchus, and Scarus sodidus had between 0.5 and 1.
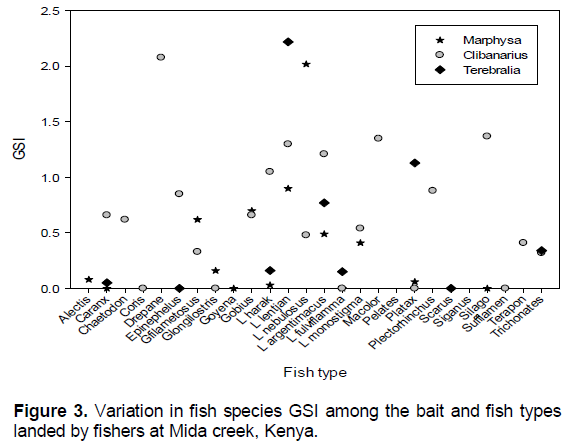
Among Terebralia fishers, the highest GSI was L. lentjan, and the lowest Epinephelus and Siganus. Among Marphysa fishers Drepane was the highest while the lowest was for G. oyena and L. fulviflamma. Among Clibanarius fishers, L. nebulosus was the highest while Terapon, Plectorhinchus and Drepane, the lowest (Figure 3).
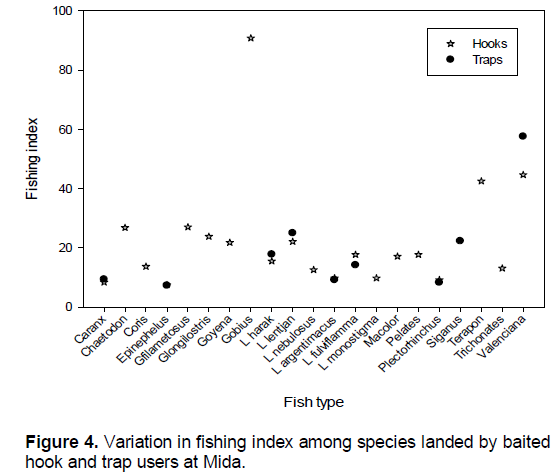
Variation in fishing index among the species and gear employed
Comparison of fishing index among the species landed were significantly different (χ2=3323, df-2184, P<0.0001). Only two fish species Valenciana (50.9) and Gobius (90.9) had a fishing index of above 50% (Figure 4). Chaetodon, Epinephelus, L. argetimacus, L. monostigma, Platax, and Plectorhinchus were harvested before achieving 10% of adult size, while other species had 10 to 50% of adult body dimensions (Figure 4).
Among hook users, three fish taxa had above 40% fishing index; Gobius keiensis (91%), T. jarbua (43%) and Valenciana (45%), while five species (Caranx, Epinephelus, L. fulviflama, Macolor) were landed prior to achieving 10% of adult’s body dimension (Figure 4). Among trap gear users, the highest FI was recorded for Valenciana (58%), while four species had less than 10% the lowest (C. ignobilis, Epinephelus, L. argentimacus, Plectorhinchus).
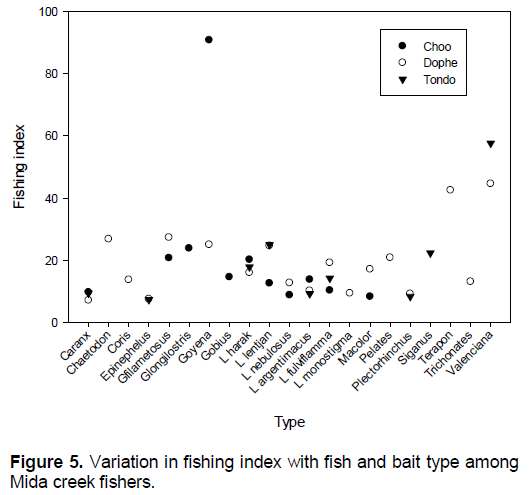
Variation in fishing index among fish species and bait types
Comparison of fishing index among fish types landed by the different bait revealed that among Marphysa fishers, the highest fishing index was 91% for Gobius, while two species Caranx (9.6), and L. fulviflama (8.8) and Plectorhinchus (8.3) had below 10% (Figure 5). Among Clibanarius fishers Valenciana, Terapon, had the highest fishing index while 4 species Caranx, Epinephelus, Macolor, Siganus had less than 10%. Among Terebralia fishers only Valenciana had above 40% of normal size.
Fish landed by bait fishers at Mida were of moderate to low condition and GSI of less than unity. The fishes landed were harvested after achieving approximately 22% of their potential adult size. Hence the fishery at Mida, targets immature sub-adult growth stages of relatively large marine fish from over 20 species. These results suggests that the fishery lands immature fish. The immediate impact of excessive harvesting of immature may be collapse of the fishery, due to failure to recruit new cohorts. Increasing fishing effort among jobless local communities will compound the current situation as reported by Verleysen and Hoorweg (2008), since bait
fishing in nearshore areas is the cheapest and easiest fishing livelihood strategy (Cinner et al., 2008). In addition, future deep water fish stocks may be depleted due to limited recruitment of immature back to the open sea, and hence affect both the open sea ecosystem and commercial fishery potentials therein.
Bait fishers along the Kenyan coastline land approximately 4Mt of fish valued at 8 million dollars annually with a calculated earning per fisher of between 1 to 2 million $ annually. Assuming all bait fishers land sub-adults (20% of adult weight), it can be estimated that continued harvesting of immature (20% of adult weight) would represent a loss to ecosystem of over 32 million dollars annually, as prospective future earnings from mature fish. Furthermore, this would be augment earnings for the estimated 4,000 to7,000 bait fishers to approximately 5 to 7 million dollars per fisher annually and potentially enhance fisher livelihoods. This could potentially lift the artisanal fisher from the poverty trap that characterizes their livelihoods.
Ninety percent of the fish types landed by all the gears and bait types were immature. The most developed fish type landed was G. keiensis which achieved up to 90% of their adult dimension. High commercial value top predator species such as carangidae, Serranidae, Siganidae and Lutjanidae, were landed at less than 10% of their potential adult dimensions. The harvesting of immature high value fish by bait fishers may limit the earning ability of the fishers since such small fishes are of limited market value. While fishers sell fresh landed fish to local fish traders and brokers at about 2$.kg-1, selling fresh high value fish directly to local tourist hotels may earn fishers double that amount. Tourism is an important foreign exchange earner in Kenya, and the Kenyan coast hosts a large number of tourist hotels (GOK, 2009; Mclean et al., 2014). Direct marketing of freshly landed high value fish to coastal tourist hotels may be an important market for locally landed fish, however, penetration into this market may require the fishers to offer more suitable specimen. Ideally reduction in fishing pressure through effort controls may allow larger mature fish to dominate landings and hence leverage penetration into the lucrative tourism industry and further improve livelihoods.
Landing immature fish reported here emanate from a combination of fishing within fish nursery grounds and ovefishing. Mangrove forest fringed zones are important nursery grounds for marine fish and crustaceans and mangrove forest extent is closely correlated to marine landings (Gadjzik, et al, 2014). Similarly, landing of small immature fish may be an indicator of overfishing pressure.
While trap fishers land significantly heavier fish, no significant differences in condition factor, GSI and fishing index were detected among the gears employed. This implies that, while trap gear land larger fish, the condition and developmental stages of the fish were similar to hook gears. While trap fishers land 9 fish taxa, hook user land over 15 taxa. Difference in fish sizes and taxa landed may be attributed to gear selectivity and habitat utilization of the fish. The mesh of the basket traps allow smaller fish to escape but in addition, the traps are soaked overnight and fish retrieved live, which allows more time for smaller fish to escape. Similarity in fish morphometric index may be attributed to interconnectivity between mangrove forest, seagrass beds and coral reefs habitats utilized by the fish(Garrison et al, 2004; Gadjzik et al, 2014), that cancels out these apparent differences. Mangrove forests, apart from housing permanent residents, are also renowned for harboring transiting predatory and breeding adult fish (Krumme and Saint-Paul, 2010; Gajdzik et al., 2014).
Wambiji et al. (2008) and also Agembe et al. (2010) report higher condition factor of 0.9 to 1.2 for Siganids captured using basket trap and other gears, in the south coast of Kenya. On the other hand, Mbaru et al. (2010) report condition factors of 0.01 to 0.4 for L. lentjan and L. fulviflamma caught using nets, which are lower than values reported here. Nandikeswan and Ananda (2013) report GSI of over 4.0 in Terapon puta from Asia, which is higher than levels reported here. Zeyl et al. (2013) report GSI cutoff points for a gobid (Neogobius melanostostonus) of 1.0 and 8.0 for adult males and females respectively, which are higher than those reported here for G. keiensis. Apart from differences in derivation of respective morphometric parameters used in this and comparable studies, variations in distribution patterns and also fishing pressure on the fish populations, are implied. It is plausible that deployment of artisanal gears in different habitats (apart from within mangrove, and creek) may explain the observed results. In addition, anecdotal evidence from Uyombo, a landing beach used by migrant Pemba fishers at Mida creek, indicates that these fishers use larger traps, to land relatively larger fish from the open sea, than local fishers. It is possible that facilitating local baits fishers to venture into alternative fishing grounds, may improve landings, while reducing pressure from the immature creek fish population; however additional data is needed before implementation of such interventions.
Although heavier fish are landed using tondo bait, similarity in the condition, gonadal development and fishing index among the bait types tested are reported here. This indicates that the different baits used by artisanal fishers attract and catch fish with similar morphometric characteristics. This differs with results presented by Jacobsen and Joensen (2004) in a longline bait fishery that showed difference in catches among the baits tested at Faroe Islands. However, in the same study, Jacobsen and Joensen (2004) report higher catch using whelk than squid bait in longline fishery, but no differences in shoreline handline fishery, similar to results reported here. Chemical and visual cues emitted by baits, are crucial to ensuring catching fish (Kasumyan and Doving, 2004), hence, exposure to unfamiliar bait cues may arouse fish curiosity, leading to higher catch in novel habitats. It is possible, that deployment of nearshore benthic invertebrate bait familiar to fish may result in lower but similar catch. Pemba fishers at Uyombo seem to employ similar bait (tondo) to target larger fish in the open sea. This is an issue that requires further experimentation to compare landings and thereafter, identify alternative fishing grounds suitable for local fishers.
It was expected that fish caught by trap fishers operating within mangrove forest would be less abundant, since they primarily respond to chemical cues. This is because, in shaded and also highly turbid aquatic habitats, visual predation is limited and hence tactile, electrical and chemical cues are more commonly used. On the other hand, fish caught from the open creek fishing ground, used by hook fishers, where both visual and chemical cues operate, would be more abundant. Results presented here indicate that although the variety of fish caught in the open creek grounds was higher, their morphometric parameters were similar to those landed from mangrove habitats. This may imply that sustained fishing pressure has depleted large mature adults from both fishing grounds. Anecdotal evidence from the fishers indicates that, fishes at the creek were previously larger and more abundant, than the current situation. Verleysen and Hoorweg (2008) suggest the massive increase in fishers at Mida, was precipitated by conversion of the largely agricultural Mijikenda into fishers, due to lack of suitable alternative livelihoods. In addition, a significant proportion of artisanal fishers is partime fishers, that also takes up farming, business or employment in the tourism industry (Mr. Anyembe- Watamu –Turtle Watch, Personal Communication). Thus improvement in alternative livelihood options may provide respite for a depleted fishery.
Fish types landed by bait fishers at Mida have similar condition factor that ranged from 0.2 to 2.4. This indicates that most of the fish landed by bait fishers at Mida are in a sub-optimal condition. Additionally, over 80% of the fish landed had limited gonadal enlargement, especially carangidae (e.g. D. punctata) and Lethrinidae (e.g. L. nebulosus, L lentjan). The lowest gonadal development was reported for Siganid (S. sutor), Gerrid, Labrid (Coris), Lethrinid (e.g. L. argentimacus). Of special interest are the low GSI reported for mangrove dwelling fishes, such as gobids and mangrove snapper, which presumably spend most of their lives in the vicinity of mangrove. It is important to note that GSI values change significantly with seasonal breeding patterns and hence results on GSI differences presented here need to be taken as an indicator of potential impacts. More elaborate longer term studies of variation in GSI are needed to concretize the findings.
Only Gobid and Valenciana are harvested at an advanced stage of development having achieved over 50% of their adult size. The observed species difference in condition factor, GSI, and fishing index may indicate differences in habitat utilization among the species. It was expected that permanent residents within the creek would record higher levels of the indicators monitored. Knowledge of exploited species breeding and habitat utilization strategies may shed more light on this aspect.
Minimum size restriction and catch-release of immature and juveniles, for high value large top predatory species, such as Serranidae, Lethrinidae, Lutjanidae, and Carangidae with fishing indices below 10% of their potential adult dimension may improve future landings. Such intervention is commonly employed in temperate angler recreational fishery (Alos et al., 2009) and may require investigation in tropical regions. Similarly, the use of small mesh basket trap and also small hooks need to be evaluated and regulated. While initial resistance from resource poor fishers may be anticipated, future improved earnings within participatory management frameworks, may be essential to sustaining livelihoods in the semi-enclosed Mida creek basin. Inadequate administrative authority policing of remote shorelines, may require concerted local community participation through strengthened Beach Management Units.
The study revealed that bait fishers at Mida use basket traps baited with tondo and hooks baited with choo and dophe to catch similar immature fish, with moderate to low condition factor, GSI and fishing index. This is the first record describing the characteristics of fish landed using specific types of bait in a tropical multi-bait fishery. The differences in landed fish characteristics observed are largely attributed to overfishing, species specific growth characteristics and habitat utilization patterns. Only small bodied fishes, such as Gobidae are landed at significantly higher developmental status, while potentially larger top carnivores such as carangid, are landed as immature of low condition and gonadal development. Reduction in fishing pressure, through voluntary catch-release of juveniles and immature, minimum size restriction, coupled with identification of alternative fishing grounds and livelihoods may allow improvement in morphometrics of landings and offer a respite and sustainability to the Mida creek fishery.
The authors have not declared any conflict of interests.
This study was financed by a National Council of Science and Technology, Kenya grant. Laboratory and facilities were provided by Nairobi and Egerton Universities and Kenya Marine and Fisheries Institute. Members of the Mida Creek Conservation Community, especially J. Mwamure, facilitated interactions with fishers. This paper formed part of a BSc thesis by Hendrick Y. (Egerton University). A. Nthiga and V. Njuguna (MSc Students, University of Nairobi), participated in field and laboratory analysis, while, J. Ngatia, S. Muhamed and M. Abutrika (BSc students, University of Nairobi), participated in fieldwork. B. Otieno (KMFRI, Mombasa), assisted in field identification of fish. The contributions and suggestions made by anonymous reviewers are also appreciated.
REFERENCES
|
Agembe S, Mlewa CM, Kaunda-Arara B (2010). Catch composition, abundance and length-weight relationship of groupers (Pisces- Serranidae) from inshore waters of Kenya. Western Indian Ocean J. Mar. Sci. 9(1):91-102. |
|
|
|
Al-Zibdah M, Kan'an N (2009). Aspect of growth, reproduction and feeding of three pomacentrid fish from Gulf of Aqaba, Jordan. Jordan J. Biol. Sci. 2(3):119-128. |
|
|
Alos J, Arlinghaus R, Palmer M, March D, Alvarez I (2009). The influence of types of natural bait on fish catch and hooking location in a multispecies marine recreational fishery with implication for management. Fish. Res.
Crossref |
|
|
Arlinghaus R, Klefoth T, Kobler A, Cooke SJ (2008). Size selectivity, injury, handling time and determinants of initial hooking mortality in recreational angling for Northern Pike: The influence of type and size of bait. North Am. J. Fish. Manage. 28:123-134.
Crossref |
|
|
|
Calderone EM, Maclean SA, Sherack B (2012). Evaluation of bioelectric index analysis and Foulton condition factor as a nonlethal method for estimating short-term response of post molting alternative in Salmon (Salmo sala) to food availability. Fish. Biol. 110(2):257-271. |
|
|
|
Cheunpagdee R, Liguori L, Palomares MLD, Pauly D (2006). Bottom-up, global estimates of small scale marine fisheries catch. Fish. Centre Res. Report 14(8). |
|
|
Cinner JE, Daw T, McClanahan TR (2008). Socioeconomic factors that affect artisanal fisher's readiness to exit a declining fishery. Conserv. Biol. 23(1):124-130.
Crossref |
|
|
|
Daliri M, Paighabari SY, Shabani MJ, Pouladi M, Davoodi R (2012). Length-Weight and length growth relation, relative weight and relative condition factor of 4 commercial fish species of Northern Persian Gulf. Ann. Rev. Res. Biol. 2(1):15-26. |
|
|
|
Elders at Atafu Atoll (2012). Echoes at Fisheries Rock: Traditional Tokelau fishing. Ed A Hooper & I Tinielu, Knowledge of nature, 4, UNESCO, Paris. p. 120. |
|
|
|
Fishbase (2011). Fishbase 2011. In Froese, J.E. & Pauly, D. (Eds) World Wide Web electronic publications. |
|
|
|
Fisher RA, Fisher DL (2006). The use of bait bags to reduce the need for horseshoe crab as bait in the Virginia whelk fishery. VIMS Marine resource Report # 2006-10, Virginia USA, p. 11. |
|
|
|
Fondo EN (2004). Assessment of the Kenya marine fisheries from selected fishing areas. United Nation University Fisheries Training program, Final Training Report, Reykjavik, Iceland, p. 55. |
|
|
|
Frame Survey (2012). Marine fisheries frame survey. Ministry of fisheries Development, Fisheries Department, Nairobi, Kenya, p. 85. |
|
|
Froese R (2006). Cube law, condition factor and weight-length relationship: History, metaanalysis and recommendation. J. Appl. Ichthyol. 22:241-253.
Crossref |
|
|
Garrison VH, S Roger, J Beets, AM Friedlander (2004). The habitats exploited and the species trapped in a Caribbean island trap fishery. Environ. Biol. Fish. 71:247-260.
Crossref |
|
|
|
Gajdzik L, Vanreusel A, Koedam N, Reubens J, Muthumbi AWN (2014). The mangrove forest as a nursery habitat for the ichthyofauna of Mida creek (Kenya, East Africa). J. Mar. Assoc. United Kingdom pp. 1-13. |
|
|
|
Glaesel H (1997). They are not just 'samaki': Towards an understanding of fisher voices on the Kenyan coast. AAP 51:65-179. |
|
|
|
Government of Kenya (GOK) (2009). State of the coast: Towards integrated management of coastal and marine resources in Kenya. NEMA, Nairobi, Kenya. p. 88. |
|
|
Harnish L, Rogers JH, Beets J, Friedlander AM (2004). The habitats exploited and the species trapped in a Caribbean Island trap fishery. Environ. Biol. Fish. 71:247-260.
Crossref |
|
|
|
Hoorweg J, Versleyen N, Wangila B, Degen A (2006). Income diversification and fishing practice among artisanal fishers on the Malindi-Kilifi coast. Presented at the Coastal Ecology Conference, May 2006, Mombasa. |
|
|
|
Jacobsen JH, Joensen J (2004). Comparison of bait in longline fishery. Bsc Thesis University of Faroe Islands. |
|
|
|
Kaunda-Arara B, Rose GA, Muchiri MS, Kaka R (2003). Long term trends in coral reef fish yields and exploitation rates of commercial species from coastal Kenya. Western Indian Ocean J. Mar. Sci. 2(2):105-116. |
|
|
Kasumyan AO, Doving KB (2004). Taste preferences in fish. Fish. Fish. 4:289-343.
Crossref |
|
|
|
Kreiner A, Van Der Linden CD, Freon P (2001). A comparison of condition factors and GSI of sardine (Sardinops sagax) stocks in the Northern and Southern Benguela upwelling ecosystems, 1984-1999. In A decade of Namibian Fish Science, Payne A.I. Pillar S.C., Crawford R.J.M., (Eds). South Afr. J. Mar. Sci. 23:123-134. |
|
|
|
Krumme U, Saint-Paul U (2010). Dynamics in mangrove fish assemblages on a macrotidal coast. Saint-Paul U., Schneider H., (Eds), In Mangrove dynamics and management in North Brazil. Ecol. Stud. 211:189-209. |
|
|
Mbaru EK, Mlewa CN, Kimani EN (2010). Length weight relationship of 39 selected reef fishes in the Kenya coastal artisanal fisheries. Fisher. Res. 106:567-569.
Crossref |
|
|
|
Mclean B, Kingwill K, Govender J, Ogega VN (2014). Reef and Marine recreational management (RMRM) thematic area Kenya. Marine Tourism management Operation Strategy. Version 1, Eco-Africa Environ. Consultant, p. 57. |
|
|
|
Mwaipopo R, Fischer E, Wanyonyi I, Kimani P, Tunje J, Msuya F, Bashemerewa V (2011). The relationship between community based organizations and the effective management of coastal and marine resources in the WIO region. A report for the WIOMSA 8:85. |
|
|
|
Nandikeswan R, Anandan V (2013). Analysis of the gonadosomatic index and fecundity of Terapon puta from Nallavadu coast Pondicherry. Int. J. Sci. Res. Pub. ISSN 3(2): 2250-3153. |
|
|
|
Nash RDM, Valencia AH, Geffen AF (2006). The origin of Fulton's condition factor- setting the record straight. Fisheries 31(5):236-239. |
|
|
|
Richmond MD (2011). A field guide to the seashores of Eastern Africa and the Western Indian Ocean Islands. 3rd Ed, Sida/WIOMSA; p. 464. Richter H, Lückstädt C, Focken UL, Becker K (2000). An improved procedure for the assessment of fish condition on the basis of length weight relationship. Arch. Fish. Mar. Res. 48(5):226-235. |
|
|
|
Samoilys M, Maina GW, Osuka K (2011). Artisanal fishing gears of the Kenya coast. CORDIO, Mombasa, Kenya. |
|
|
|
Tuda A Omar M (2012). Protection of marine areas in Kenya. The George Wright Forum 29(1):43-50. |
|
|
|
Tuda P, Nyaga W, Maina GW, Wanyonyi I, Obura D (2008). Total fishing effort over tidal to annual periods in the Diani-Chale-Gazi reef fishery in Kenya. In Obura DO; J Tamelander, O Linden, Eds, Ten years after the bleaching; facing the consequences of climate change in the Indian Ocean. CORDIO status report SIDA-SAREC, Mombasa, Kenya. pp. 321-335. |
|
|
Van der Elst R, Everett B, Jiddawi N, Mwatha G, Afonso PS, Buolle D (2005). Fish, fishes and fishers of the Western Indian Ocean; their diversity and status, A preliminary assessment. Philos. Trans. Royal Soc. A, 363:263-284.
Crossref |
|
|
|
Verleysen N, Hoorweg J (2008). From farming to fishing: Marine resource conservation a new generation of fishermen. Western Indian Ocean J. Mar. Sci. 7(1):1-14. |
|
|
|
Wambiji N, Ohtomi J, Fulanda GW, Kimani E, Kulundu N, Hossain MY (2008). Morphometric relationship and condition factor of Siganus stellatus, S. caniculatus, S. sutor (Pisces; Siganidae) from Western Indian Ocean waters. South Pac. Stud. 29(1):1-16. |
|
|
|
Zeyl JN, Love OP, Higgs DM (2013). Evaluating gonadosomatic index as an indicator of reproductive development in the invasive round gobid Neogobius melanostostonus. J. Great lakes Res. Article in press 8 pp. |