Full Length Research Paper

ABSTRACT
The goals of this study were to evaluate dourado female egg viability submitted to stress condition caused by two different reproductive induction methods, extruded and semi-natural. Sixteen females randomly chosen were induced with pituitary extract and allocated in two groups; the first, in which oocytes were manually extruded and the second group, females were allowed to naturally spaw. Blood samples were collected for glucose, cortisol and hematocrit determinations as well as for red blood cells and white blood cells smears evaluations. A control group had its blood collected before hormonal induction. Fertilization rate was 40.6 and 91.7% for extruded and semi-natural methods, respectively, and the survival rate was higher in semi-natural groups. Both experimental groups showed higher glucose and cortisol levels compared to control group. The results for hematocrit, hemoglobin concentration and erythrocytes numbers did not differ among groups. Extruded and semi-natural procedures elevated monocytes percentage. In conclusion, the semi-natural procedures are more efficient than extruded method and should be taken into account for Salminus brasiliensis handling of breeders.
Key words: Fish breeders, reproductive methodology, hematological parameters.
INTRODUCTION
Dourado, Salminus brasiliensis (Cuvier, 1816), from Characidae family, is broadly distributed at Pantanal, Paraná, Uruguay, São Francisco watersheds as well as at Lagoa dos Patos associated-basin (Morais and Schubart, 1955; Gomes et al., 2003). It is a carnivorous fish, found in lotic environmental and performing ascending reproductive migrations (Streit et al., 2007). Zaniboni and Schulz (2003) assigned the dwindling of this specie in nature to riparian deforestation, fishing and capture of young specimens, drainage of adjacent lagoons and alterations of hydrological regime caused by dams’ construction, water contamination and nonnative species introduction. Hence, S. brasiliensis is candidate for artificial reproduction programs for aquaculture preservation purposes.
More recently, some studies have demonstrated environmental factors-induced cortisol releasing to play crucial role in physiological response during the onset of sexual development and increasing steroids levels (Solomon-Lane et al., 2013; Nozu and Nakamura, 2015); however, the exact mechanism involved in cortisol signaling is still unclear (Goikoetxea et al., 2017). During induced reproductive process, the fertilization rate of S. brasiliensis is considered limited, which can be associated with inadequate procedures, commonly applied to exotic species reproduction (Zaniboni-Filho and Weingartner, 2007). Eggs quality and viability are directly related with reproductive stress (Zanoni et al., 2016).
Handling of fish breeders during hormonal induction leads to morphological, biochemical and physiological alterations that can be characterized as a stress condition (Eslamloo et al., 2014). In this context, fish stress monitoring through attendance of physiological conditions is an important and valuable tool for aquaculture. The studies of Mazeaud et al. (1977), Barton and Iwama (1991)and Schreck (2010)proposed that fish physiological stress response initiates with stressor agent perception, which leads to hypothalamic-pituitary-interrenal axis catecholamines releasing. The elevated levels of these hormones in turn result in a secondary physiological response that influence other organs and systems causing osmotic status, immunological status and bioenergetic use to change, leading to important alterations of reproductive capacity .
Hematological parameters are important tools to evaluate fish physiological status, may serve as an indicator of stress and reflects the associated pathological alterations (Fazio et al., 2012; Zanoni et al., 2016). To recognize these alterations and to compare them with reference values for a determined species allow to quantify the duration and the severity of stress (Fazio et al., 2015). In addition, hematological profile has been used as biomarker for aquaculture (Tavares-Dias et al., 2007)and able to validate stress conditions in fish. Blood sample analysis predict pathological alterations in an organism since hematopoietic process is under influence of several biological and environmental factors (Tavares-dias et al., 1999). Hematocrit alterations such as red blood cells numbers and hemoglobin levels (Graham et al., 1985), are normally investigated in this context. The RBC elevation results from the releasing of young erythrocytes caused by spleen contraction (Caldwell and Hinshaw, 1994). Yet, there is a considerable variation in the results found by different authors.
Among blood figurative elements, leukocytes play crucial role in nonspecific immunity, and changes in its value can be attributed to the fishes’ welfare (Misra et al., 2006). Particularly, leukocyte profile is useful as a physiological indicator of stress. Plasma glucocorticoids disturbance can increase neutrophil numbers (neutrophilia) and reduce limphocyte concentrations (lymphocytopenia), and as result, the proportional neutrophil to limphocyte ratio is positively associated to the intensity of stressor agent as well as to the levels of circulating glucocorticoids. Some evidences also point to infections and diseases caused by cortisol excess to influence neutrophil to lymphocyte ratio (Davis et al., 2008).
In order to put forward conservational aquaculture, there is an increasing need to achieve higher breeder survival indexes and improve quality of their eggs. During induced reproduction, maintenance of physiological homeostasis and minimization of stress procedures could lead to an increased number of healthy fish fry. The aim of this study was to evaluate the fertilization rate, survival, cortisol and glucose concentrations; and, hematological parameters from dourado breeders underwent two different induced reproduction procedures; semi-natural and extruded methods.
MATERIALS AND METHODS
The experiment was conducted with 32 males and 16 females of dourado randomly chosen from the broodstock of Hydrology and Aquaculture Station of Duke Energy International, located at Salto Grande, Sao Paulo State, Brazil (49° 130 W and 23° 100 S).
From December 2009 to January 2010, specimens were captured by trawling, selected based on their external indicators of gonad maturation (hyperaemic urogenital papilla, bulged and soft womb) and transported to laboratory with 50 L plastic bags, containing 1/5 water . Then, fish were anesthetized with 1 g/100 L of water benzocaine solution, and weighed. The body weight was used to calculate the pituitary extract dosage. After these procedures, breeders were transferred to maintenance tanks and kept under 28°C, with constant water flux to anesthesia recovery. To reproductive induction processes, it was utilized carp pituitary gland extract macerated in porcelain crucible and diluted in saline (0.9%) at room temperature. Females received two intraperitoneal applications , one considered preparatory of 0.5 mg/kg, injected near the pectoral fin peduncle using a 5 ml syringe and hypodermic needles and the second one, 5 mg/kg, 12 h after the former. Male dourado were injected a single dose of 2.5 mg/kg pituitary extract at the time of the second dose of females.
For semi-natural procedures, 8 females and 16 males were randomly transferred to an external, circular tank, with 5.1 m radius and 2.0 m deep, receiving constant flow of 131 L/s of water, at 28°C. This architecture creates an unidirectional water flux, in such a way that the eggs could be collected by a drainage system at the bottom of the tank, communicating directly with a 200 L Woynarovich’ incubator, with continuous flow of 7 L/s. Retained eggs were collected every hour for a period of 6 h consecutively, then they were taken to the laboratory where they remained in Israelis-like incubators until hatching (Zanoni et al., 2016). The number of eggs was calculated by multiplying the average of three different samples of one mL by the total volume of eggs.
For extrusion procedure, at 8.5 h after the last dose of pituitary extract, all females were anesthetized with benzocaine and the oocytes extruded manually and collected in plastic containers. Two males were randomly chosen, anesthetized and semen collected by extrusion was added to oocytes and mixed dryly . Hydration of oocytes and activation of male gametes were completed by adding water at 28ºC, followed by the transfer to Israelis-like incubators. The number of eggs was calculated as previously described.
For both procedures, the following reproductive efficiency indexes were calculated:

To measure the concentration of cortisol, 1 mL of blood was collected by venipuncture in caudal region using 3 ml syringes embedded in EDTA 10% and 28 x 12 needles . Female blood samples of the two experimental groups were collected soon after spawning and control group blood sample was collected soon after fish were captured. Cortisol concentration was determined by ELISA using commercial kit (EIA, 55050, Human-Kit; Cayman Chemical, Ann Arbor, MI, USA) and glucose levels were determined with digital glucometer ACCU CHECK active®. 1 mL of blood was also collected and added to tubes with EDTA 10% for hematological analysis that were carried out at the Clinic Pathology Laboratory of Universidade Estadual do Norte do Paraná – UENP, Paraná, Brazil. Blood smears, in duplicate, were stained with May–Grunwald–Giemsa dye (Rosenfeld, 1947). The indirect total and differential leucocyte count were performed as proposed by Hrubec and Smith (2000). Our results were evaluated by one-way ANOVA and complemented with Tukey, p≤0.05, while data are expressed as mean±SD.
RESULTS AND DISCUSSION
In the semi-natural procedure, females produced 8 L of hydrated eggs, with 46 eggs in each mL, resulting in 368,000 eggs and in extruded procedure, the result was 636.000 eggs, in a final volume of 12 L with 53 eggs/L as shown in Table 1. In both procedures, 100% of dourado females responded to the treatment with pituitary extract. According to Bromage et al. (1994), eggs releasing at the time of spawning is crucial for egg quality; in such way, the release of eggs before or after this event can lead to low fertilization rates and low quality fish larvae (Springate and Bromage, 1985).
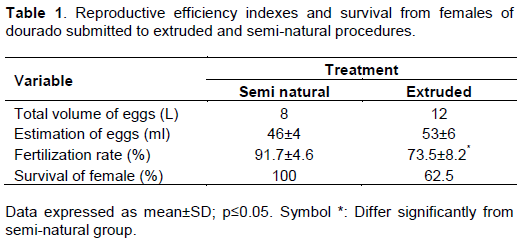
Fertilization rate can be an indicative for eggs quality, and immature eggs display small perivitelinic space caused by incomplete cortical activation which in turn can compromise fertilization process and embryogenic development (Kjørsvik et al., 1990; Zanoni et al., 2016). Our results show fertilization rate to be higher in semi-natural method compared to extruded method as described in Table 1. This result corroborates David et al. (2002) , who obtained fertilizations rate of 25.8% for extruded and 94.5% for semi-natural spawning of Leporinus macrocephalus, and also by Zanoni et al. (2016)with Brycon orbignyanus, who described fertilization rates of 87.9 and 8.17%, for semi-natural and extruded procedures, respectively.
The percentage of females that survived the reproduction procedures was 100 and 62.5% for semi-natural and extruded, respectively. Notably, 24 h after the extrusion processes, all the females of the last group died. The mortality rate after extruded procedure for S. brasiliensis is normally high. Sato et al. (1997) related 83.3% death of extruded breeders, which showed several lesions, body hiperemia, bacteria and fungi secundary infections. All these symptoms can be caused by loss of scales during fish manipulation (Elaine et al., 2002), and as in our case could, in part, explaining our findings pointing the first caution with extruded procedure.
Barton (2002)proposed ideal levels for corticosteroids to be under 5.0 ng/mL of blood, although in fishes, this range between 30-40 ng/mL. In Figure 1, it can be seen that cortisol concentration in control, semi-natural and extrude groups were 11.27±4.26, 22.74±12.93 and 34.9±11.26 ng/mL respectively. These results are in accordance with previous studies from our group and with other authors who have not found differences in this hormonal parameter in fish submitted to stressor conditions (Gomes et al., 2003; Hoshiba, 2009; Zanoni et al., 2016). However, in semi-natural group, the lower cortisol level and higher matrix survival can be attributed to an improved recovering period since the animal were displaced in an external circular tank with bigger space and increased water flow which is similar to what is found in natural environments (Boesoaard et al., 1993; Milligan et al., 2000).
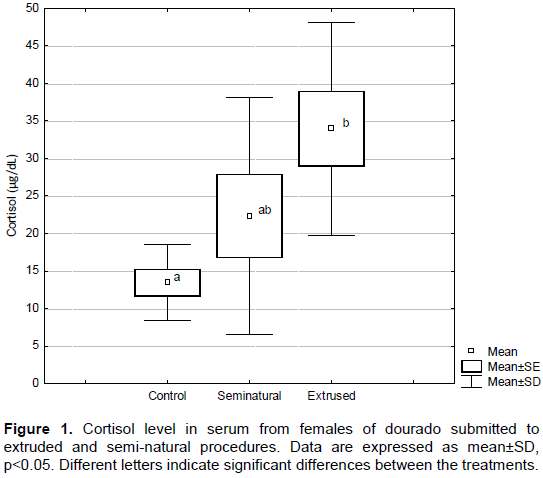
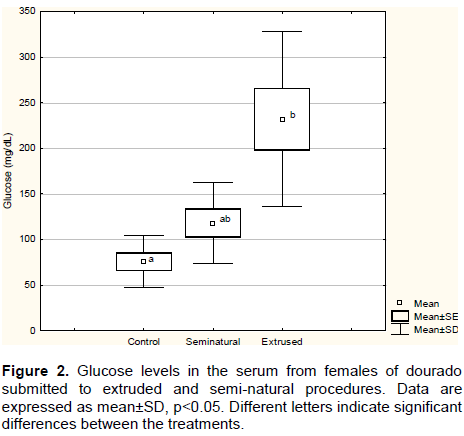
Our outcomes for fertilization and cortisol levels demonstrate the stress effects for eggs quality, which can be explained by cortisol influence on fish reproductive glands. Nepomnaschy et al. (2006) and Whirledge and Cidlowski (2010)showed the necessity of glucocorticoids levels for a normal gonadal function. Elevated levels of cortisol have a negative impact over fertility rates (Carragher and Sumpter, 1990); furthermore, stress itself can suppress reproductive hormones (Pickering et al., 1987; Pankhurst, 1994; Haddy and Pankhurst, 1999). Extruded procedure increased glucose circulating levels compared to control and semi-natural groups, as it can be seen in Figure 2. Similar response was observed by Gomes et al. (2003)and (Brandão and Levy, 2006), for Arapaima gigas, and Gomes et al. (2003)for Colossoma macropomum. Glucose levels elevation is considered as an adaptive response during the stress response and is related to cortisol and others catecholamine’s levels. Taken together, cortisol and glucose levels can be considered good predictors of stress in S. brasiliensis.
According to Lucelle et al. (2004), hematological evaluation of fishes during fish farming can reflect its physiological status. Stress conditions increase cortisol concentration modifying physiologic and metabolic status, which can be demonstrated by erithrocytes elevation and mean corpuscular volume (VosylienÄ—, 1999). Alterations found in erythrogram helps the identification of anemic conditions, and some alteration seen in leucograms is useful for infectious process and homeostasis imbalance diagnostics. Data presented in Table 2 demonstrate that there is no statistically difference for hemoglobin concentration, hematocrit and number of red blood cells comparing experimental groups. Similarly, Abreu and Urbinati (2006), for Brycon amazonicus,Martins et al. (2004), for Oreochromis niloticus and Pimpão (2006), for Ancistrus multispinis, found no difference in hematocrit in fish under stress conditions. Number of red blood cells and hemoglobin concentration, otherwise, differ from Das and Mukherjee (2003)and Adhikari et al. (2004), who demonstrated reduction in these parameters for Labeo rohita exposed to stressor agents. It can be assumed that, for dourado, semi-natural and extruded methodologies do not alter eritropoietic process, besides that, for the same species, different results for hemoglobin concentration and hematocrit can be caused by blood samples collection techniques, including anticoagulants and anesthetic use, time elapsed between collection and laboratorial analysis, among others (Tavares-Dias and Sandrim, 1998). More studies are needed to the understanding of these differences.

Teleosts blood composition depends on physiological and ecological factors such as sex, gonadal development stage, stress, infection and environmental conditions. Leukocytes number vary among families, genus and species, and it is related with the surrounding environment (Tavares-dias and Moraes, 2004) There was no difference in total leukocytes when comparing both methods as described in Table 3. According to Abreu and Urbinati (2006), feeding Brycon cephalus breeders with different concentrations of vitamin C submitted to air exposure, and Pimpão (2006), studying Ancistrus multispinis exposed to deltametrin, found similar results. For the most of fishes’ species, leukocytosis can be found as soon as a stressor agent is present in attempt to recover from homeostasis disruption, while leukopenia is generally attributed to diseases that affect immunological system (VosylienÄ—, 1999). In this study, leukocytes total number was unaltered despite cortisol levels alterations. Nevertheless, monocytes number was higher in both experimental groups compared to control as shown in Table 3. This cell type acts in inflammatory responses involving phagocytosis process (Dalmo and Bøgwald, 2008; Salvador et al., 2013). The result demonstrated here point the reproductive induction process in Dourado to be considered an inflammatory stimulus, recruiting monocytes for local tissue, since they are considered primary cells for antigens presenting in teleosts.
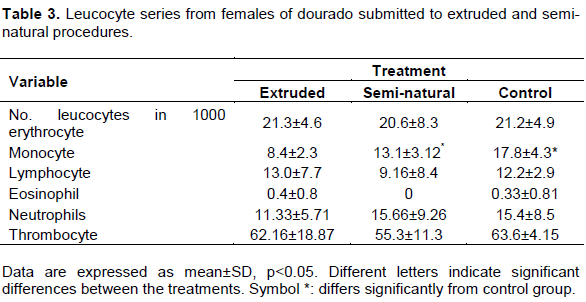
Neutrophils, lymphocytes and thrombocytes numbers did not differ among groups as seen in Table 3. Stress leads to a lymphocyte redistribution, mainly in lymphoid organs, lowering their circulating levels or, the high levels of cortisol in response to a stressor agent can induce lymphocytes death (Benfey and Biron, 2000). Wojtaszek et al. (2002)confirmed this last alternative by demonstrating accentuated lymphopenia in Cyprinus carpio 24 h after cortisol inoculation. Other authors however have related higher number of neutrophils after stress conditions in Salmo trutta e Salmo gairdneri (Johansson-Sjobeck and Larsson 1979; Pickering et al., 1987). Thrombocytes number found in our experiment are in accordance with Benfey and Biron (2000). These researches studying Oncorhynchus mykiss and Salvelinus fontinalis under feedlot and manipulation stress, have verified a slight, but non-statistically significant increase in the number of thrombocytes in fish. This cell type is produced at kidneys and spleen in teleost fishes and play diverse functions, being in a constant flux between circulation and organs (Tavares-dias and Moraes, 2004). Lower number of thrombocytes in circulation could be related to the moment of blood sample collection.
CONCLUSION
Our result demonstrates S. brasiliensis manipulation by utilizing semi-natural procedure or by extrusion causes similar stress characteristics. The improved egg quality obtained by semi-natural procedure is relevant and needed to point out. Taken into account, semi-natural proceeding should be preferred in order to achieve a larger number of healthy larvae
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Abreu JS de, Urbinati EC (2006). Physiological responses of matrinxã (Brycon amazonicus) fed different levels of vitamin C and submitted to air exposure. Acta Amaz 36:519-524. |
|
|
Adhikari S, Sarkar B, Chatterjee A (2004). Effects of cypermethrin and carbofuran on certain hematological parameters and prediction of their recovery in a freshwater teleost, Labeo rohita (Hamilton). Ecotoxicology and Environmental Safety 58:220-226. |
|
|
Barton BA (2002). Stress in Fishes: A Diversity of Responses with Particular Reference to Changes in Circulating Corticosteroids. Integrated Computer and Biology 42:517-525. |
|
|
Barton BA, Iwama GK (1991). Physiological changes in fish from stress in aquaculture with emphasis on the response and effects of corticosteroids. Annual Review of Fish Diseases 1:3-26. |
|
|
Benfey TJ, Biron M (2000). Acute stress response in triploid rainbow trout (Oncorhynchus mykiss) and brook trout (Salvelinus fontinalis). Aquaculture 184:167-176. |
|
|
Boesoaard L, Nielsen ME, Rosenkilde BR (1993) Moderate exercise decreases alaska cortisol levels in atlantic salmon (Salmo Salar). Computer and Biochemistry Physiology M:641-643. |
|
|
Brandão FR, Levy de C, Gomes ECC (2006). Respostas de estresse em pirarucu (Arapaima gigas) durante práticas de rotina em piscicultura practices (importancia na aquicultura to lendo) 36:349-356. |
|
|
Bromage N, Bruce M, Basavaraja N (1994). Egg Quality Determinants in Finfish The Role of Overripening with Special Reference to the Timing of Stripping in the Atlantic Halibut Hippoglossus hippoglossus. Journal of World Aquactic Society 25:13-21. |
|
|
Caldwell CA, Hinshaw J (1994). Physiological and haematological responses in rainbow trout subjected to supplemental dissolved oxygen in fish culture. Aquaculture 126:183-193. |
|
|
Carragher JF, Sumpter JP (1990). The effect of cortisol on the secretion of sex steroids from cultured ovarian follicles of rainbow trout. General Computer Endocrinology 77:403-407. |
|
|
Dalmo RA, Bøgwald J (2008) ß-Glucans As Conductors of Immune Symphonies. Fish and Shellfish Immunology 25:384-396. |
|
|
Das BK, Mukherjee SC (2003). Toxicity of cypermethrin in Labeo rohita fingerlings: Biochemical, enzymatic and haematological consequences. Computer and Biochemistry Physiology - C Toxicology Pharmacology 134:109-121. |
|
|
David AR-T, Betina ME, Juan RE (2002). Reproducción inducida del piauçu, Leporinus macrocephalus garavello y britski, 1988 (characiformes, anostomidae). Bol do Inst da Pesca 28:11-18. |
|
|
Davis AK, Maney DL, Maerz JC (2008). The use of leukocyte profiles to measure stress in vertebrates : a review for ecologists. Functional Ecology 22(5):760-772 |
|
|
Elaine FA-T, Emico TK, Massuka YNNF-V (2002) Reprodução Induzida da Piabanha, Brycon insignis (Steindachner, 1876), Mantida em Cativeiro. Brazilian Journal of Animal Science 31:803-811. |
|
|
Eslamloo K, Akhavan SR, Fallah FJ, Henry MA (2014). Variations of physiological and innate immunological responses in goldfish (Carassius auratus) subjected to recurrent acute stress. Fish Shellfish Immunology 37:147-153. |
|
|
Fazio F, Ferrantelli V, Fortino G (2015). The influence of acute handling stress on some blood parameters in cultured sea bream (Sparus aurata Linnaeus, 1758). Italian Journal of Food Safety 4:4-6. |
|
|
Fazio F, Filiciotto F, Marafioti S (2012). Automatic analysis to assess haematological parameters in farmed gilthead sea bream (Sparus aurata Linnaeus, 1758). Marine and Freshwater Behaviours and Physiology 45:63-73. |
|
|
Goikoetxea A, Todd EV, Gemmell NJ (2017). Stress and sex: Does cortisol mediate sex change in fish? Reproduction 154:R149-R160. |
|
|
Gomes LC, Araujo-Lima CARM, Roubach R (2003). Effect of fish density during transportation on stress and mortality of juvenile Tambaqui Colossoma macropomum. Journal of World Aquactic Society 34:76-84. |
|
|
Graham MS, Haedrich RL, Fletcher GL (1985). Hematology of three deep-sea fishes: a reflection of low metabolic rates. Computer and Biochemistry Physiology -- Part A Physiology 80:79-84. |
|
|
Haddy JA, Pankhurst NW (1999). Stress-induced changes in concentrations of plasma sex steroids in black bream. Journal of Fish Biology 55:1304-1316. |
|
|
Hoshiba M (2009). Respostas fisiológicas de estresse no matrinxã (Brycon amazonicus) após exercício físico intenso durante a captura. Acta Amaz 39:445-452. |
|
|
Hrubec TC, Smith S (2000). Hematology of fish. In: Feldman.B.F.; Zinkl, J.G., Jain NCS (ed) Veterinary Hematology, 5 ed. London pp. 1120-1125. |
|
|
Johansson-Sjobeck ML, Larsson A (1979). Effects of Inorganic Lead on Delta-Aminolevulinic Acid Dehydratase Activity and Hematological Variables in the Rainbow Trout, Salmo gairdnerii. Archives of Environmental Contamination Toxicology 8:419-431. |
|
|
Streit Jr S, Sirol RN, Ribeiro RP (2007). Parâmetros qualitativos do sêmen de Dourado (Salminus maxillosus) em cativeiro. Bol Inst Pesca 34:337-344. |
|
|
Kjørsvik E, Mangor-Jensen A, Holmefjord I (1990). Egg quality in fishes. Advances in Marine Biology 26:71-113. doi: 10.1016/S0065-2881(08)60199-6 |
|
|
Lucelle DA; Chagas EC, Gomes LC, Brandao FR (2004). Efeito de banhos terapêuticos com formalina sobre indicadores de estresse em tambaqui Lucelle. Pesquisa Agropecuária Brasileira 39:217-221. |
|
|
Martins ML, Pilarsky F, Onaka EM (2004). Hematologia e resposta inflamatória aguda em Oreochromis niloticus (osteichthyes : cichlidae) Submetida aos estímulos único e consecutivo de estresse de captura. InstPesca 30:71-80. |
|
|
Mazeaud MM, Mazeaud F, Donaldson EM (1977). Primary and secondary effects of stress in fish: some new data with a general review. Transactions of the American Fisheries Society 106(3):201-212. |
|
|
Milligan CL, Hooke GB, Johnson C (2000). Sustained swimming at low velocity following a bout of exhaustive exercise enhances metabolic recovery in rainbow trout. Journal of Experimental Biology 203:921-926. |
|
|
Misra CK, Das BK, Mukherjee SC, Meher PK (2006). The immunomodulatory effects of tuftsin on the non-specific immune system of Indian Major carp, Labeo rohita. Fish Shellfish Immunology 20:728-738. |
|
|
Morais FMB, Schubart O (1955). Contribuição no estudo do dourado (Salminus maxillosus Val.) do rio Mogi Guassu (Pisces, Characidae) (1aed.), Ministério da Agricultura: São Paulo-SP, Brazil 131 p. |
|
|
Nepomnaschy PA, Welch KB, McConnell DS (2006). Cortisol levels and very early pregnancy loss in humans. Proceedings of the National Academy of Sciences 103:3938-3942. |
|
|
Nozu R, Nakamura M (2015). Cortisol administration induces sex change from ovary to testis in the protogynous wrasse, Halichoeres trimaculatus. Sex Development 9:118-124. |
|
|
Pankhurst NW (1994). Effects of caputure and recovery on plasma levels of cortisol, lactate and gonadal steroids in a natural population of raiwbon trout. Journal of Fish Biology 45:1013-1025. |
|
|
Pickering AD, Pottinger TG, Carragher J, Sumpter JP (1987). The effects of acute and chronic stress on the levels of reproductive hormones in the plasma of mature male brown trout, Salmo trutta L. General and Comparative Endocrinology 68:249-259. |
|
|
Pimpão CT (2006). Avaliação aguda dos efeitos toxicológicos da deltametrina em uma espécie de peixe fluvial nativo: estudo bioquímico e imunotóxico. Dr. Thesis University Federal do Paraná, Brazil. |
|
|
Salvador R, Silva CG, Loureiro BA (2013). Desempenho e hematologia de tilápias-do-nilo alimentadas com saccharomyces cerevisiae e vacinadas contra streptococcus agalactiae. Pesquisa Agropecuária Brasileira 48:892-898. |
|
|
Sato Y, Fenerich-Verani N, Verani JR, Godinho HP, Vieira L (1997). Reprodução artificial do dourado Salmilllls brasilieflsis (Pisces : Characidae) da bacia do rio São Francisco (Pisces : Characidae) from the São Francisco river basin). Revista Brasileira de Reprodução Animal 21:113-116. |
|
|
Schreck CB (2010). Stress and fish reproduction: The roles of allostasis and hormesis. General and Comparative Endocrinology 165:549-556. |
|
|
Solomon-Lane TK, Crespi EJ, Grober MS (2013). Stress and serial adult metamorphosis: Multiple roles for the stress axis in socially regulated sex change. Frontiers in Neuroscience 7(2013):210. |
|
|
Springate JRC, Bromage NR (1985). Effects of egg size on early growth and survival in rainbow trout (Salmo gairdneri Richardson). Aquaculture 47:163-172. |
|
|
Tavares-dias M, Moraes F (2004). Hematologia de peixes teleósteos (1aed.), Ed.Eletrônica e Arte Final. Ribeirão Preto-SP, Brazil . 144 p. |
|
|
Tavares-dias M, Henrique S, Schalch C (1999). Hematologia de teleósteos brasileiros com infecção parasitária . I . Variáveis do Leporinus macrocephalus Garavelo e Britski , 1988 (Anostomidae) e Piaractus mesopotamicus Holmberg , 1887 (Characidae). Patologia (Mex) 21:337-342 |
|
|
Tavares-Dias M, Ono EA, Pilarski F, Moraes FR (2007). Can thrombocytes participate in the removal of cellular debris in the blood circulation of teleost fish? A cytochemical study and ultrastructural analysis. Journal of Applied Ichthyology 23:709-712. |
|
|
Tavares-Dias M, Sandrim EFS (1998). Influence of anticoagulants and blood storage on hematological values in tambaqui, Colossoma macropomum. Acta Scientiarum. Biological Sciences 20:151-155. |
|
|
VosylienÄ— MZ (1999). The Effect of Heavy Metals on Haematological Indices of Fish (Survey). Acta Zoologica Lituanica 9:76-82. |
|
|
Whirledge S, Cidlowski JA (2010). Glucocorticoids, stress, and fertility. Minerva Endocrinology 35:109-125. |
|
|
Wojtaszek J, Dziewulska-Szwajkowska D, Lozińska-Gabska M (2002). Hematological effects of high dose of cortisol on the carp (Cyprinus carpio L.): Cortisol effect on the carp blood. General and Comparative Endocrinology 125:176-183. |
|
|
Zaniboni-Filho E, Weingartner M (2007). Técnicas de indução da reprodução de peixes migradores. Revista Brasileira de Reprodução Animal 31:367-373. |
|
|
Zaniboni E, Schulz U (2003). Migratory fishes of the Uruguay River. In: Migratory Fishes of South America. Victoria, pp. 168-205. |
|
|
Zanoni MA, Costa FG, de Carvalho S, Seiva FR (2016). Physiological and biochemical changes of females of Piracanjuba, subjected to induced reproduction. Journal of Animal Physiology and Animal Nutrition (Berl) 100: |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


