Full Length Research Paper

ABSTRACT
The Citrus sp. from Parang Island Karimunjawa is a wild type of Citrus plant that grow on salt area and exhibits a high level of vitamin C. Due to its ability, morphological, molecular and nutritional characterization needs to be carried out to improve its potential. The study was carried out at the Biotechnology Laboratory and the Integrated Laboratory of Diponegoro University from 2016 to 2017. The characterization was based on the morphplogical appearence of the tree, its fuits, and leaves. Its DNA barcoding consists of 18S ribosomal RNA and the ITS region on the plants is dispersed along the coastal ends and the centre of the island. Furthermore, the nutritional characterization consists of an edible fruit part, a high vitamin C level and protein content. The research showed that the Citrus sp. on the coastal ends and at the center of Parang Island Karimunjawa exhibits a close relation with the C. hystrix and members of Papeda clade. However, the morphology of the Citrus is quite different from the common C. hystrix in Indonesia. The nutritional content also shows that the Vitamin C content is higher than that of the C. hystrix by almost four times. The features of the Citrus sp. in Parang Island Karimunjawa have potential and showed possibility to improve its superiority in industrial applications and breeding programs.
Key words: Karimunjawa, Parang, Citrus, 18S rRNA, ITS, nutrition.
INTRODUCTION
MATERIALS AND METHODS
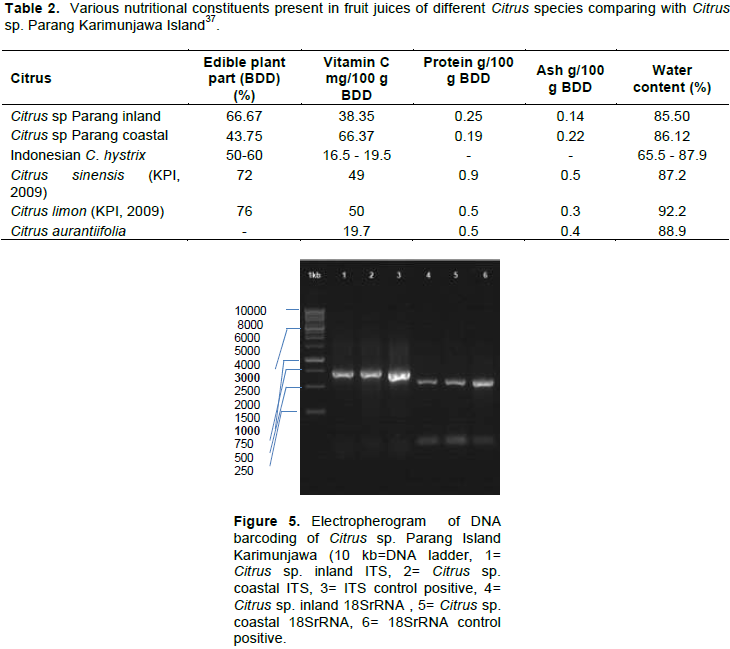
RESULTS



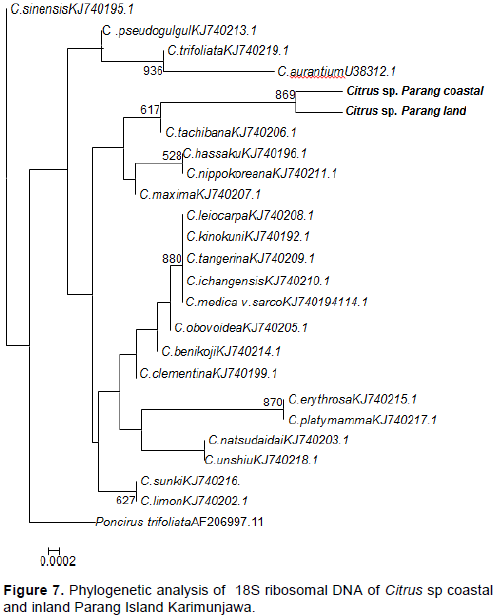
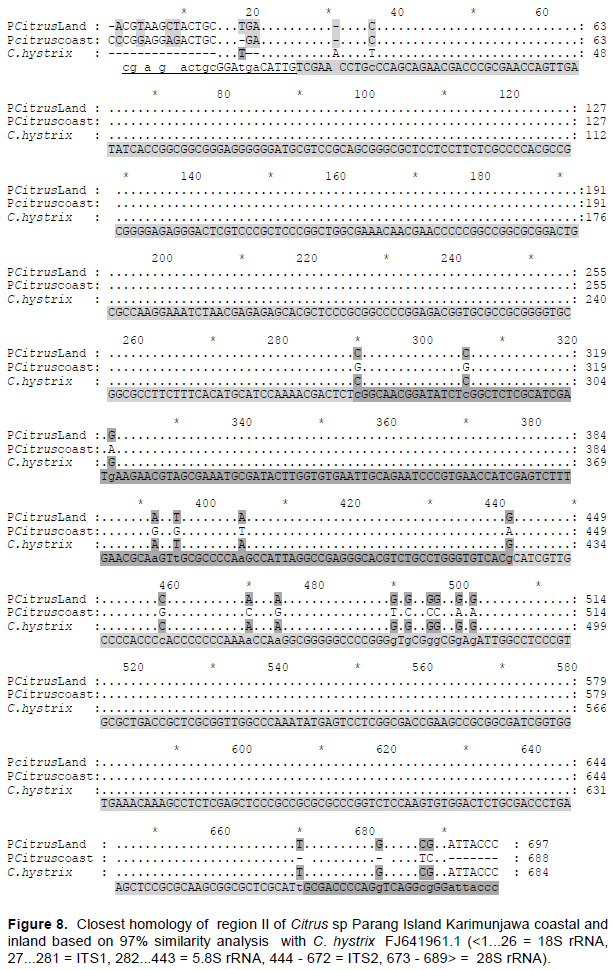
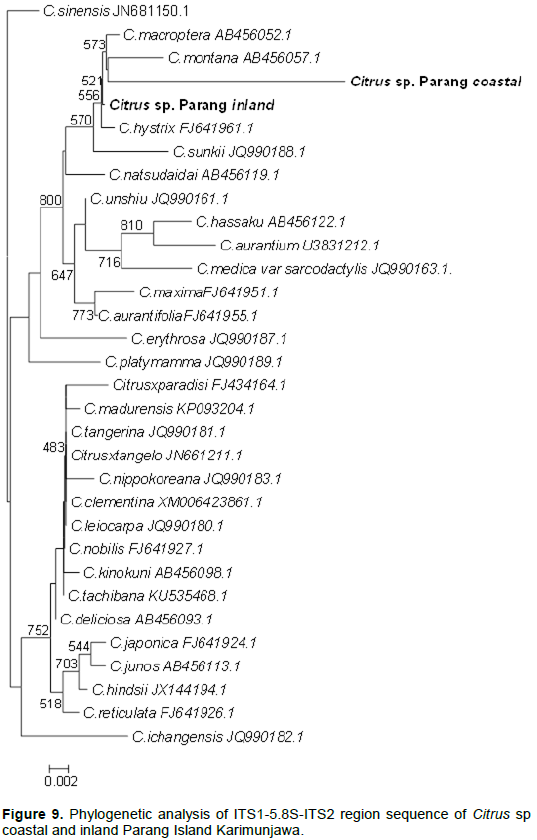
DISCUSSION
CONCLUSION
ACKNOWLEDGEMENT
CONFLICT OF INTERESTS
The authors declare that they have no conflict of interest.
REFERENCES
|
Abirami A, Nagarani G, Siddhuraju P (2014). In vitro antioxidant, anti-diabetic, cholinesterase and tyrosinase inhibitory potential of fresh juice from Citrus hystrix and C. maxima fruits. Food Science and Human Wellness 3:6-25. |
|
|
Acosta-Motos J, Ortu-o MF, Bernal-Vicente A, Diaz-Vivancos P, Sanchez-Blanco M, Hernandez JA (2017). Plant responses to salt stress: adaptive mechanisms. Agronomy Journal 7(18):1-38. |
|
|
Adelina SO, Adelina E (2017). Identifikasi morfologi dan anatomi jeruk lokal (Citrus sp) di desa Doda dan desa Lempe Kecamatan Lore. Jurnal Agrotekbis (In Indonesian) 5(1):58-65. |
|
|
Agisimanto D, Martasari C, Supriyanto A (2007). Perbedaan primer RAPD dan ISSR dalam identifikasi hubungan kekerabatan genetik jeruk siam (Citrus suhuniensis L. Tan) Indonesia. Jurnal Hortikultura (In Indonesian) 17(1):101-110. |
|
|
Álvarez I, Wendel JF (2003). Ribosomal ITS sequences and plant phylogenetic inference. Molecular Phylogenetics and Evolution 29:417-434. |
|
|
AOAC (Association of Official Analitycal Chemists) (2005). Official method of analysis (18th Edition). Washington.DC: Association of Officiating Analytical Chemists. |
|
|
Bayer RJ, Mabberley DJ, Morton C, Miller CH, Sharma IK, Pfeil BE, Rich S, Hitchcock R, Sykes S (2009). A molecular phylogeny of the orange subfamily (Rutaceae: Aurantioideae) using nine cpDNA sequences. American Journal of Botany 96(3):668-685. |
|
|
Bulley SM, Rassam M, Hoser D, Otto W, Schünemann N, Wright M, MacRae E, Gleave A, Laing W. (2009). Gene expression studies in kiwifruit and gene over-expression in Arabidopsis indicates that GDP-L-galactose guanyltransferase is a major control point of vitamin C biosynthesis. Journal of Experimental Botany 60(3):765-778. |
|
|
Chiba H, Uehara M, Wu J, Wang X, Masuyama R, Suzuki K, Kanazawa K, Ishimi, Y (2003). Hesperidin, a citrus flavonoid, inhibits bone loss and decreases serum and hepatic lipids in ovariectomized mice. Journal of Nutrition 133(6):1892-1897. |
|
|
Ciancaglini P, Santos HL, Daghastanli KRP, Thedei G (2001). Using a classical method of vitamin C quantification as a tool for discussion of its role in the body. Biochemistry and Molecular Biology Education 29(3):110-114. |
|
|
Davey MW, Kenis K, Keulemans J (2006) Genetic control of fruit vitamin C contents. Plant Physiology 142(1):343-351. |
|
|
Direktorat Jenderal Holtikultura (2015). Sub Sektor Holtikultura, (In Indonesian). |
|
|
Doyle JJ, Doyle JL (1987). A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochemical Bulletin 19:11-15. |
|
|
Fang D, Krueger RR, Roose ML (1998). Phylogenetic relationships among selected Citrus germplasm accessions revealed by inter-simple sequence repeat (ISSR) markers. Journal of the American Society for Horticultural Science 123(4):612-617. |
|
|
Felsenstein J (2004). PHYLIP (Phylogeny Inference Package) Version 3.6. (Distributed by the author) Department of Genome Sciences, University of Washington, Seattle. |
|
|
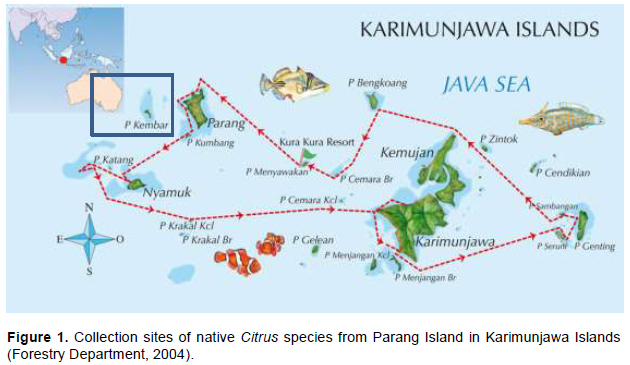
Forestry Department (2004). Zonatıon of Karımunjawa natıonal park. Jepara Regency. Central Java Provınce. |
|
|
Gosslau A, Chen KY, Ho CT, Li S (2014). Anti-inflammatory effects of characterized orange peel extracts enriched with bioactive polymethoxyflavones. Food Science and Human Wellness 3:26-35. |
|
|
Hanin M, Ebel C, Ngom M, Laplaze L, Masmoudi K (2016). New insights on plant salt tolerance mechanisms and their potential use for breeding. Frontiers in Plant Science pp. 1-17. |
|
|
Hernandez JA, Jimenez A, Mullineaux P, Sevilla F (2000). Tolerance of Pea (Pisum sativum L.) to long-term salt stree is associated with antioxidant defences. Plant, Cell and Environment 23(8):853-862. |
|
|
Hirai M, Mitsue S, Kita K, Kajiura I (1990). A survey and isozyme analysis of wild mandarin, Tachibana (Citrus tachibana (Mak.) Tanaka) growing in Japan. Japanese Society for Horticultural Science 59:1-7. |
|
|
Hussain S, Luro F, Costantino G, Ollitrault P, Morillon R (2012). Physiological analysis of salt stress behaviour of Citrus species and genera: Low chloride accumulation as an indicator of salt tolerance. South African Journal of Botany 81:103-112. |
|
|
Hynniewta M, Malik SK, Rao SR (2014). Genetic diversity and phylogenetic analysis of Citrus (L) from north-east India as revealed by meiosis, and molecular analysis of internal transcribed spacer region of rDNA. Meta Gene 2:237-251. |
|
|
Johnson LA, Soltis DE, Soltis PS (1999). Phylogenetic relationships of Polemoniaceae inferred from 18S ribosomal DNA sequences. Plant Systematics and Evolution 214:65-89. |
|
|
Jukes TH, Cantor CR (1969). Evolution of protein molecules. In Munro HN, editor, Mammalian Protein Metabolism, Academic Press, New York 3(21):132. |
|
|
Karsinah, Sudarsono, Setyobudi L, Aswidinnoor H (2002). Keragaman genetik plasma nutfah jeruk berdasarkan analisis penanda RAPD. Jurnal Bioteknologi Pertanian (In Indonesian) 7(1):8-16. |
|
|
Koehler-Santos P, Dornelles ALC, De Freitas LB (2003). Characterization of mandarin citrus germplasm from Southern Brazil by morphological and molecular analyses. Pesquisa Agropecuária Brasileira 38(7):797-806. |
|
|
Kumar R, Vijay S, Khan N (2013). Comparative nutritional analysis and antioxidant activity of fruit juices of some Citrus spp. Octa Journal of Biosciences 1(1):44-53. |
|
|
Kusumanıngrum HP (2008). Karakterisasi alga hijau Dunaliella sp. dan isolat sianobakteria serta deteksi gen DXS penyandi enzim kunci biosintesis karotenoid [Characterization of green microalgae Dunaliella sp. and cyanobacteria isolate and detection of DXS gene encoding key enzyme of carotenoid biosynthetic] [PhD Dissertation]. Yogyakarta: Gadjah Mada Universitas (In Indonesian). |
|
|
Kyndt T, Dung TN, Goetghebeur P, Toan HT, Gheysen G (2010). Analysis of ITS of the rDNA to infer phylogenetic relationships among Vietnamese Citrus accessions. Genetic Resources and Crop Evolution 57(2):183-192. |
|
|
Li X, Xie R, Lu Z, Zhou Z (2010). The origin of cultivated Citrus as Inferred from internal transcribed spacer and chloroplast DNA sequence and amplified fragment length polymorphism fingerprints. Journal of The American Society for Horticultural Science 135:341-350. |
|
|
Liu F, Wang L, Gu L, Zhao W, Su H, and Cheng X. (2015). Higher transcription levels in ascorbic acid biosynthetic and recycling genes were associated with higher ascorbic acid accumulation in blueberry. Food Chemistry 188:399-405. |
|
|
Mahmud MK, Hermana N, Zulfianto A, Rozanna R, Apriyantono NI, Hartati B, Bernardus T (2009). Table of Indonesian food composition. Jakarta. Indonesian Nutritionist Association (In Indonesian). |
|
|
Mellidou I, Kanellis AK (2017). Genetic control of ascorbic acid biosynthesis and recycling in horticultural crops. Frontiers in Chemistry Mini Review 5(40):1-8. |
|
|
Morton CM (2009). Phylogenetic relationships of the Aurantioideae (Rutaceae) based on the nuclear ribosomal DNA ITS region and three noncoding chloroplast DNA regions, atpB-rbcL spacer, rps16, and trnL-trnF. Organisms Diversity and Evolution 9(1):52-68. |
|
|
Moya JL, Primo-Millo E, Talon M (1999). Morphological factors determining salt tolerance in Citrus seedlings: The shoot to root ratio modulates passive root uptake of chloride ions and their accumulation in leaves. Plant Cell and Environment 22:1425-1433. |
|
|
Murkute A, Sharma S, Singh SK (2005). Citrus in terms of soil and water salinity: A review. Journal of Scientific and Industrial Research 64(06):393-402. |
|
|
Nei M, Kumar S (2000) Molecular evolution and Phylogenetics. Oxford University Press pp. 33-46 |
|
|
Novelli VM, Cristofani M, Souza A, Machado M (2006). Development and characterization of polymorphic microsatellite markers for the sweet orange (Citrus sinensis L. Osbeck). Genetics and Molecular Biology 29(1):90-96. |
|
|
Nuryandani, E (2012). Persebaran dan karakterisasi induk jeruk keprok Tawangmangu asli (Citrus reticulata Blanco ssp Tawangmangu) (In Indonesian) 13(1):33-42. |
|
|
Penjor T, Mimura T, Matsumoto R, Yamamoto M, Nagano Y (2014). Characterization of limes (Citrus aurantifolia) grown in Bhutan and Indonesia using high-throughput sequencing. Scientific Reports 4:1-9. |
|
|
Rafiq S, Kaul R, Sofi SA, Bashir N, Nazir F, Nayik GA (2016). Citrus peel as a source of functional ingredient: A review. Journal of The Saudi Society of Agricultural Sciences 7:1-8. |
|
|
Rogers SO, Bendich AJ (1987). Ribosomal RNA genes in plants: variability in copy number and in the intergenic spacer. Plant Molecular Biology 9(5):509-520. |
|
|
Samraj S, Rajamurgugan S (2017). Qualitative and quantitative estimation of bioactive compounds and antioxidant activity in Citrus hystrix. International Journal of Engineering Science and Computing 7(6):13154-13163. |
|
|
Shimizu T, Kitajima A, Nonaka K, Yoshioka T, Ohta S, Goto S, Toyoda A, Fujiyama A, Mochizuki T, Nagasaki H, Kaminuma E, Nakamura Y (2016). Hybrid origins of citrus varieties inferred from DNA marker analysis of nuclear and organelle genomes. PLoS ONE 11(11): 1-58. |
|
|
Singh A, Saini ML, Behl RK (2003). Screening of citrus rootstocks for salt tolerance in semi-arid climates-A review. Tropics 13:53-66. |
|
|
Spínola V, Mendes B, Câmara JS, Castilho PC (2013). Effect of time and temperature on vitamin C stability in horticultural extracts. UHPLC-PDA vs iodometric titration as analytical methods. LWT - Food Science and Technology 50:489-495. |
|
|
Sun YL, Kang HM, Han SH, Park YC, Hong SK (2015). Taxonomy and phylogeny of the genus Citrus based on the nuclear ribosomal and its region sequence. Pakistan Journal of Botany 47(1):95-101. |
|
|
Syvertsen JP, Melgar JC, García-Sánchez F (2010). Salinity tolerance and leaf water use efficiency in Citrus. Journal of the American Society for Horticultural Science 135(1):33-39. |
|
|
Tamura K, Glen S, Daniel P, Alan F, Sudhir K (2013). MEGA6: molecular evolutionary genetics analysis version 6.0. Molecular Biology and Evolution 30(12):2725-2729. |
|
|
Tomascir T, Mah AJ, Nontji A, Moosa MK (1970). The ecology of the Indonesian Seas, Part Two. Hong Kong: Eric Oey, Periplus Editions Ltd pp. 685-686. |
|
|
Uchoi A, Malik SK, Choudhary R, Kumar S, Pal D, Rohini MR, Chaudhury R (2017). Molecular markers in assessing genetic variation of Indian citron (Citrus medica L.) cultivars collected from different parts of India. Indian Journal of Biotechnology 16:346-356. |
|
|
Uzun A, Yesiloglu T, Aka-Kacar Y, Tuzcu O, Gulsen O (2009). Genetic diversity and relationships within Citrus and related genera based on sequence related amplified polymorphism markers (SRAPs). Scientia Horticulturae 121(3):306-312. |
|
|
Vijayan K (2009). Approaches for enhancing salt tolerance in mulberry (Morus L) -A review. Review Article. Plant Omics Journal 2(1):41-59. |
|
|
Wang X, Xu Y, Zhang S, Cao L, Huang Y, Cheng J, Wu G, Tian S, Chen C, Liu Y, Yu H, Yang X, Lan H, Wang N, Wang L, Xu J, Jiang X, Xie Z, Tan M, Larkin RM, Chen L-L, Ma B-G, Ruan Y, Deng X, Xu X (2017). Genomic analyses of primitive, wild and cultivated citrus provide insights into asexual reproduction. Nature 49(5):765-775. |
|
|
White TJ, Burns T, Lee S, Taylor J (1990). Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. M.A. Innis. D.H. Gelfand, J.J. Sninsky, T.J. White (Eds.), PCR Protocols: A Guide to Methods and Amplifications, Academic Press, San Diego & London pp. 315-322. |
|
|
World Conservation Monitoring Centre (1998). Limnocitrus littoralis. The IUCN Red List of Threatened Species 1998: e.T37412A10051534. |
|
|
Wu GA, Terol J, Ibanez V, Lopez-Garcia A, Perez-Roman E Borredá C, Domingo CR, Tadeo FR, Carbonell-Caballero J, Alonso R, Curk F, Du D, Ollitrault P, Roose ML, Dopazo J, Gmitter Jr FG, Rokhsar DS, Talon M (2018). Genomics of the origin and evolution of Citrus. Nature 554:311-316. |
|
|
Xu Q, Chen L, Ruan X, Chen D, Zhu A, Chen C, Bertrand D, Jiao W, Hao B, Lyon MP, Chen J, Gao S, Xing F, Lan H, Chang J, Ge X, Lei Y, Hu Q, Miao Y, Wang L, Xiao S, Biswas MK, Zeng W, Guo F, Cao H, Yang X, Xu X, Cheng Y, Xu J, Liu J, Luo OJ, Tang Z, Guo W, Kuang H, Zhang H, Roose ML, Nagarajan N, Deng X, Ruan Y (2013). The draft genome of sweet orange (Citrus sinensis). Nature Genetics 45(1):59-66. |
|
|
Yahata M, Kunitake H, Komatsu H (2017). Morphological characterization and evaluation of reproductive function in a haploid pummelo [Citrus maxima (Burm.) Merr.]. Japan Agricultural Research Quarterly 51(4):293-298. |
|
|
Yamaji H, Kondo K, Kuniga T, Nesumi H, Yoshida T, Hashimoto K, Takheda O (2013). Origin of cultivated Citrus (Rutaceae) documented by the contents of internal transcribed spacer sequences (ITS) in nuclear ribosomal DNA. Journal of Japanese Botany 88:222-238. |
|
|
Ywassaki LA, Canniatti-Brazaca SG (2011). Ascorbic acid and pectin in different sizes and parts of citric fruits. Ciência e Tecnologia de Alimentos 31(2):319-326. |
|
|
Zhang C, Liu J, Zhang Y, Cai X, Gong P, Zhang J, Wang T, Li H, Ye Z. (2011). Overexpression of SlGMEs leads to ascorbate accumulation with enhanced oxidative stress, cold and salt tolerance in tomato. Plant Cell Report 30(3):389-398. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


