Full Length Research Paper

ABSTRACT
Chicken is a major livestock species raised by smallholder households in rural areas in developing countries of Sub-Saharan Africa as a chief source of meat protein and income generation. However, Newcastle disease largely compromises the production of this precious bird under backyard production settings. The purpose of the study was to assess antibody responses and growth performance following Newcastle disease vaccination in kuroiler, broiler, and local Tanzanian chickens raised under the same tropical environmental condition. In the experiment, a total of 358 chickens comprised of 127 kuroilers, 121 broilers, and 110 local Tanzanian chickens were raised for seven weeks, and body weights were recorded weekly. Birds were vaccinated at four weeks, and blood samples were collected at days 10, and 21 post-vaccination for antibody titres quantification by ELISA test. Results demonstrated higher (P < 0.05) antibody titres in kuroilers (3.81 ± 0.06) as compared to local chicken (3.73 ± 0.07) and broilers (3.53±0.06) at day 10 post-vaccination. The growth performance of local Tanzanian chickens was comparable to that of kuroilers. The present findings contribute to on-going work in understanding chicken immune responses against NDV and inform breeding programs designed for developing chickens with increased resistance to NDV.
Key words: Tanzania, local chicken, kuroilers, Newcastle disease virus, growth performance, ND vaccination, Sub-Saharan Africa, antibody responses
INTRODUCTION
The world chickens population is estimated at 18 billion (Conan et al., 2012), and about 80% of chickens that are raised in Africa are indigenous local chickens (Conan et al., 2012). The majority (60%) of African households raise chickens under backyard production settings where chickens are left to scavenge for their nutritional need (Mapiye et al., 2008; Mtileni et al., 2009). In Tanzania, 96% of livestock farmers keep local chickens, which supply 94% of poultry meat and eggs in rural areas (Ministry of Livestock and Fisheries Development- MLFD, 2015).
Local chickens are well adapted to harsh tropical environmental conditions and survive persistent exposure to endemic infectious diseases (Msoffe et al., 2001; Minga et al., 2004: Muchadeyi et al., 2017). The growth of chicken is rapid and its farming requires relatively small land size. More importantly, chicken has a high nutritive value from chicken eggs and meat, and serves as a chief source of high-quality meat protein among households in resource-poor countries in SSA (Ahlers et al., 2009; Mtileni et al., 2009; Martin et al., 2015). In Africa setting, chickens particularly local chickens play a crucial role in socio-cultural functions such as traditional ceremonies and rituals (Mtileni et al., 2009; Conan et al., 2012).
However, local chickens are characterized by low productivity traits like low growth performance and eggs production (Yakubu and Ari, 2018). Efforts are made to introduce improved chicken genetic resources adapted to harsh tropical environmental conditions, for example, with an introduction of kuroiler chicken, a dual-purpose scavenger hybrid chicken raised for meat and egg production, which was imported to Africa from India (Sharma et al., 2015; Fleming et al., 2016; Yakubu and Ari, 2018). Introduction of improved breeds is in an agreement with Tanzania Livestock Modernization Initiative (TLMI) (MLFD, 2015), which among other key priority actions in poultry modernization is the identification of dual-purpose breeds suitable for local free-range scavenging conditions (MLFD, 2015).
Newcastle disease (ND) caused by the Newcastle disease virus (NDV) is ranked as the number one killer of chickens under backyard production systems (Kitalyi, 1998; Gondwe and Wollny, 2007; Alders et al., 2014; Yune and Abdela, 2017; Absalón et al., 2019). Current disease prevention and control option largely depends on an appropriate vaccination of susceptible flocks (Alexander, 2001; Sharif et al., 2014). The ability of chickens to elicit protective antibody against the virus is crucial to prevent disease occurrence (Pevzner et al., 1981; Yune and Abdela, 2017). Chickens variation in antibody responses against NDV has been documented (Pevzner et al., 1981; Luo et al., 2013). Selection of chicken genotypes with high protective antibody titres may have significant contribution in preventing ND outbreak in chicken populations (Kapczynski et al., 2013; Luo et al., 2013). The present study was aimed to assess antibody titres and growth performance following ND immunization in local Tanzanian chicken, kuroilers, and broilers.
MATERIALS AND METHODS
Chicken population and husbandry
Three chicken breeds (local Tanzanian chicken, kuroilers, and broilers), which were raised under the same environment and management condition were involved in the present study. Local Tanzanian chicken and kuroiler eggs were obtained from Urio Cross and Pure Breeding LTD, a local farm in Tanzania (Tengeru, Arusha, Tanzania). Eggs were incubated to hatch at the Nelson Mandela African Institution of Science and Technology (NM-AIST) Laboratory egg incubators. The incubation conditions were 37.9°C temperature and 55% humidity. The eggs were candled at 10 and 18 days of incubation to detect infertile eggs and dead embryos. On the other hand, because of difficulty of obtaining broiler chicken eggs, instead, day-old broiler chicks were obtained from commercial poultry company in Arusha, Tanzania (Tanzania Poultry Farm, Usa River, Arusha Tanzania). Therefore, the experiment started with chicks of the same age, and was housed at Livestock Training Agency-Tengeru campus (LITA-Tengeru). The broiler starter feed (0 – 28 days) and finisher (29 – 50 days) were provided ad libitum. Light was provided throughout the experiment and was also used for room temperature adjustment during brooding. The feeds manufactured by commercial company (Harsho Milling Co.Ltd, Moshi Kilimanjaro, Tanzania) were used throughout the experiment.
Experimental design
One-day-old chicks were kept in brooding chicken facility at LITA-Tengeru, and kept for four weeks to allow maternal antibodies to wane (Jalil et al., 2010). Birds at three weeks of age before vaccination were wing tagged and randomly distributed between the control (A) and the vaccination (B) group. Birds in control and challenge groups were kept in separate houses. Group B birds were randomly allocated in three replicates for each chicken type, and birds in each replicate were randomly kept in separate pen of 1.5 m2. A total of 358 chickens (Table 1) were raised for seven weeks.

Growth performance assessment
Body weights (BW) in grams (g) were recorded weekly using analytical balance to assess chicken growth performance. The weekly average BW was calculated for the duration of the experiment. The effect of vaccination on the chicken growth performance was evaluated by calculating the mean body weight gain (BWG). The BWG was obtained by taking the body weight difference between week 7 (BW, 21 days post-vaccination) and Week 4 (BW pre-vaccination).
Virus and vaccination
Newcastle disease virus live vaccine (Vir 116, Freeze Dried, LaSota strain, Biovac, Akiva, Israel) was dissolved in 40 μl sterile normal saline in accordance with the manufacturer instructions to reconstitute to virus titer of 106.5 EID50. The vaccine was stored at 4°C until use as recommended by manufacturer. The viability of the reconstituted virus was confirmed by inoculating 10-day-old embryonated chicken eggs with 0.1 ml of the virus suspension. At 4 weeks of age, birds in the vaccination group were inoculated via an ocular route with 100 μL of the virus suspension (106.5 EID50), and 50 μL into each eye using a micropipette. Likewise, birds in control group were given 100 μL of phosphate buffered saline (PBS) as mock infection via the same route.
Blood sampling
Initially, blood samples were collected from chickens that were selected randomly before vaccination was conducted to evaluate if there was prior expose of chickens to NDV. Subsequent blood samples collection was done on days 10 and 21 post-vaccination. The blood samples were drawn from wing vein into sterile micro-tube. The chicken blood samples were properly labeled by using the wing tag number assigned to each bird. The coagulated blood samples were centrifuged at 3000 rpm to harvest sera, which were transferred into clean sterile micro-tube for storage at -20°C until use.
Antibody titers measurement
Antibody titers were quantified using NDV ELISA antibody test kit (BioCheck (UK) Ltd., Hounslow, London). Samples were tested in duplicate in accordance with the manufacturer’s instructions. Sample optical density determined spectrophotometrically using 96-wells microliter plate reader (SYNERGY|HTX multi-mode reader, BioTek Instruments Inc, Winooski, VT, USA) at 405 nm wavelengths. The calculation and interpretation of ELISA results was in accordance with the manufacturer guidelines. Briefly, the S/P ratio was calculated by using the formula: (mean of test sample−mean of negative control)/(mean of positive control−mean of negative control), and then at 1:500 dilution, the Log10 titer =1.0×Log (S/P) +3.52. Serum sample with S/P value ≥ 0.35 or titer value ≥ 1159 was considered positive for antibodies against NDV.
Data analysis
Descriptive and inferential statistical analyses were performed using R software (Team, 2017). One-way analysis of variance (ANOVA) and Turkey’s HSD were conducted to assess difference in BWG and average antibody titers between chicken types. Also, the Student t-test was used to test difference between BWG between control and challenge birds, and to test difference between average antibody titers between two time points (days 10 and 21 post challenge). The relationship between BWG and antibody titers was evaluated by performing correlation test. Significance differences were taken at p <0.05 levels.
Ethical statement
The experiment was conducted in compliance with the Guidelines on the Humane Treatment of Laboratory Animals as stipulated in the Tanzania Animal Welfare Act, 2008.
RESULTS AND DISCUSSION
Local chickens are well adapted to harsh tropical environmental conditions and survive persistent exposure to endemic infectious diseases (Msoffe et al., 2001; Minga et al., 2004; Mpenda et al., 2019). However, local chickens are characterized by low productivity traits like low growth performance and eggs production (Yakubu and Ari, 2018). Efforts are made to introduce improved chicken genetic resources adapted to harsh tropical environmental conditions, for example, with an introduction of kuroiler chicken (Yakubu and Ari, 2018). Introduction of improved breeds is in agreement with TLMI (MLFD, 2015), which among other key priority actions in poultry modernization is the identification of dual-purpose breeds suitable for local free-range scavenging conditions. The present study was aimed to assess antibody responses following ND vaccination and growth performance of kuroilers, broilers, and local Tanzanian chickens.
Findings demonstrated higher (P < 0.05) antibody titres in kuroilers (3.81 ± 0.06) as compared to local chicken (3.73 ± 0.07) and broilers (3.53±0.06) at day 10 post-vaccination. The antibody titres remained higher in kuroilers and local Tanzanian chickens as compared to antibody titres in broilers at day 21 post-vaccination (Table 3). As expected, the highest growth performance observed in broiler chicken as compared to kuroilers and local Tanzania chickens (Table 2). However, the growth performance was not significant different (P > 0.05) between kuroilers and local Tanzanian chickens.
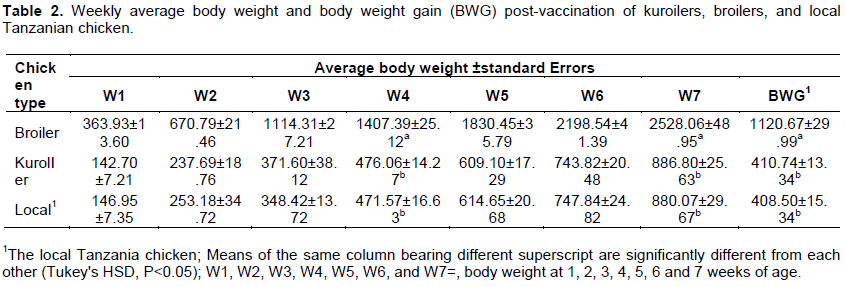
Body weight and body weight gain
The weekly means ± standard errors of body weight and BWG post-vaccination are presented in Table 2. The broilers had the highest growth performance throughout the experiment followed by kuroilers. The kuroiler chickens had higher (P>0.05) body weight than local chickens except at the first three weeks (W1, W2, and W3), whereas, the body weights of kuroilers and local Tanzanian chickens were not different (P>0.05) from week four (W4) to the rest of the experimental duration. Furthermore, the BWG was higher (P>0.05) in the control group than in the challenge group (Figure 1).
The broilers average body weights of 1407.39 ± 25.12 g and 2528.06 ± 48.95 g at 4 and 7 weeks of age, respectively reported in the present study are higher as compared to the previous reports (Munisi et al., 2015; Adeleke et al., 2011). In the previous report in Tanzania, the mean broilers body weights were 396 and 1255 g at 4 and 8 weeks of age, respectively (Munisi et al., 2015). In another study that was conducted in Nigeria, the broiler mean body weight was 2360 g at 20 weeks of age (Adeleke et al., 2011). The higher average broilers body weight observed in the present study may be explained by breed difference of the broilers involved in the studies. Also, the differences may be explained by feed content variations between the studies. For example, 4 weeks old broilers that were fed with feed supplemented with baobab seed oil cake had body weight of 1266 g, which is comparable to the broilers mean body weight of 1407.39 ± 25.12 g reported in the present study (Chisoro et al., 2018).
Kuroiler is dual-purpose chicken raised for egg and meat production (Dessie and Getachew, 2016). Like local chickens, kuroilers can thrive under harsh tropical environmental conditions, and they can scavenge for nutrition needs just like local chickens (Ahuja et al., 2008; Dessie and Getachew, 2016). The breed outperforms local chickens in terms of meat and egg production (Ahuja et al., 2008; Yakubu and Ari, 2018). However, in the present study, kuroilers (886.80 ±25.63) and local Tanzania chicken (880.07 ±29.67) body weights were comparable (P > 0.05). The discrepancy may be explained by differences in chicken breeds involved in the studies. Local Tanzania chickens are not well characterised and due to lack of genetic resources conservation programs, possibly local chicken involved in the present study have acquired high growth performance genetic materials from improved breed through interbreeding. Also, feed regime used in the present study probably had significant contribution on growth performance observed in kuroilers and local Tanzanian chickens. This is corroborated by higher body weights observed in the present study as compared to the previous reports. For example, Egyptian chicken (Mandarah) mean body weights at 4 and 7 were 299 and 747 g, respectively (Taha et al., 2012). In another study conducted in Nigeria to evaluate growth characteristic of kuroiler and Nigerian local chicken (Fulani), at 6 weeks of age the mean body weight of kuroiler and Fulani was 450 and 228 g, respectively (Yakubu and Ari, 2018). In another study conducted in Tanzania involving local chickens, the mean body weights at 4 and 7 weeks of age were 151 and 419 g, respectively (Munisi et al., 2015).
Antibody titres following ND vaccination
The means ± standard errors of antibody response against NDV at two time points (days 10 and 21 post-vaccination) are presented in Table 3. The antibody titers were highest in kuroiler chickens compared to antibody titers in local and broiler chickens. The antibody titers of kuroiler and local chickens at 10 days post-vaccination were different (P < 0.05) from that of broilers. The kuroiler chickens had higher (P > 0.05) antibody titres than local chickens at two time points. Overall, the antibody titers were different (P < 0.0001) between days 10 and 21 post challenge. Mean (3.91 ± 0.04) antibody titer at 21-days post challenge was higher than the mean (3.69 ± 0.04) antibody titer at 10-days post-vaccination (Figure 2). Furthermore, there was positive correlation of antibody titers between days 10 and 21 post challenge. The correlation of antibody titer between day 10 and 21 was intermediate in broiler (r = 0.52; P = 4.1e-5) and local (r =0.4; p=0.006) chickens as compared to weak correlation in kuroiler (r = 0.1; p = 0.4) chickens.

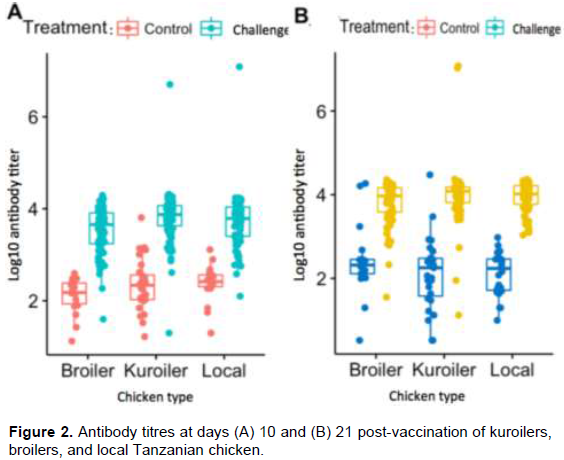
Although not supported by empirical studies, kuroilers are said to be resistant to infectious diseases (Sharma et al., 2015; Fleming et al., 2016). The higher antibody titres observed in kuroilers likely corroborated the previous reports, which suggest increased disease resistance in kuroilers.
The antibody titers were different (P < 0.0001) between control and challenged group in all chicken types at the two time points (Figure 2). The antibody titers were not different (P = 0.37) in the three chicken types at days 10 and 21 in the control group (Figure 2).
Generally, the observed antibody titers in the present study are relatively higher compared to the previous findings in other parts of the world. For example, Luo et al. (2013), reported mean antibody titer of 3.3 at 41 days after second immunization among Chinese-yellow broiler chickens. Another report found the mean antibody titer of 3.2 at 15 days post challenge among commercial meat chicken type (Tabidi et al., 2004). Generally, the comparison of the antibody responses in the present study with the previous reports is difficult because of the difference in environmental conditions, NDV strains used in vaccination, and detection tests involved.
CONCLUSION AND RECOMMENDATION
Kuroiler chickens demonstrated higher antibody titers following ND vaccination as compared to local Tanzania chickens and broilers raised under the same environmental and management condition. However, it remains to be established whether the high antibody titres observed in kuroiler chickens can be translated to an increased protective effect against virulent NDV infection in the field. Also, the growth performance of kuroilers and local Tanzanian chicken were not different, which suggests that breeds and feeding regiment employed in the present study had an influence on the observation. Overall, the finding contributes to on-going work on understanding chicken immune responses against NDV and informs breeding programs designed for developing chickens that have increased resistance to NDV. Also, findings suggest genetic characterisation of local chicken genetic resources and to establish the levels of gene flows between local chicken and improved commercial breeds existing in Tanzania.
ACKNOWLEDGMENTS
We thank the Bill & Melinda Gates Foundation for funding this research (grant #OPP1083453) through the Program for Enhancing Health and Productivity of Livestock (PEHPL) and the Nelson Mandela African Institution of Science and Technology in Arusha (NM-AIST), Tanzania for hosting the program.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Absalón AE, Diana V, Cortés-Espinosa E, Lucio P, Miller J, Afonso CL (2019). Epidemiology, control, and prevention of Newcastle disease in endemic regions: Latin America. Tropical Animal Health and Production 51(5):1033-1048. |
|
|
Adeleke MA, Peters SO, Ozoje MO, Ikeobi CON, Bamgbose AM, Adebambo OA (2011). Growth performance of Nigerian local chickens in crosses involving an exotic broiler breeder. Tropical Animal Health Production 43:643-650. |
|
|
Ahlers C, Alders R, Bagnol B, Cambaza A, Harun M, Mgomezulu R, Msami H, Pym B, Wegener P, Wethli E (2009). Improving village chicken production: a manual for field workers and trainers. ACIAR, Canberra, Australia, Series No. 139. |
|
|
Ahuja V, Dhawan M, Punjabi M, Maarse L (2008). Chickening out of poverty? Story of 'kuroiler'from India. |
|
|
Alders R, Awuni JA, Bagnol B, Farrell P, Haan N (2014). Impact of avian influenza on village poultry production globally. Ecohealth 11:63-72. |
|
|
Alexander DJ (2001). Newcastle disease. British Poultry Science 42:5-22. |
|
|
Chisoro P, Nkukwana TT, Mupangwa JF, Mabusela TP (2018). Feed intake, growth performance and carcass traits of broilers fed diets with various inclusion levels of baobab seed oilcake. South Africa Journal of Animal Science 48:284-294-294. |
|
|
Conan A, Goutard FL, Sorn S, Vong S (2012). Biosecurity measures for backyard poultry in developing countries: a systematic review. BMC Veterinary Research 8:240-250. |
|
|
Dessie T, Getachew F (2016) The Kuroiler Breed. African Chicken Genetic Gains Fact Sheet 2. Nairobi: ILRI. |
|
|
Fleming D, Koltes J, Markey A, Schmidt C, Ashwell C, Rothschild M, Persia M, Reecy J, Lamont S (2016). Genomic analysis of Ugandan and Rwandan chicken ecotypes using a 600k genotyping array. BMC Genomics 17(1):407-423. |
|
|
Gondwe TN, Wollny CBA (2007). Local chicken production system in Malawi: Household flock structure, dynamics, management and health. Tropical Animal Health Production 39:103-113. |
|
|
Jalil MA, Samad MA, Islam MT (2010). Evaluation of maternally derived antibodies against Newcastle disease virus and its effect on vaccination in broiler chicks. Bangladesh Journal of Veterinary Medicine 7:296-302. |
|
|
Kapczynski DR, Afonso CL, Miller PJ (2013). Immune responses of poultry to Newcastle disease virus. Devevlopmental and Comparative Immunology 41(3):447-453. |
|
|
Kitalyi AJ (1998). Village chicken production systems in rural Africa: Household food security and gender issues. FAO, Rome, Italy. |
|
|
Luo C, Qu H, Ma J, Wang J, Li C, Yang C, Hu X, Li N, Shu D (2013). Genome-wide association study of antibody response to Newcastle disease virus in chicken. BMC Genetics 14:42-50. |
|
|
Mapiye CM, Mwale JF, Mupangwa M, Chimonyo RF, Mutenje MJ (2008). A research review of village chicken production constraints and opportunities in Zimbabwe. Asian-Australasian Journal of Animal Sciences 21:1680-1688. |
|
|
Martin L, Jeremiah A, Michael D, Janet P (2015). Management, feeding and breeding practices of local chickens in the remote areas of Morobe province, Papua New Guinea. Journal of South Pacific Agriculture 18(2):1-52. |
|
|
Minga UM, Msoffe PL, Gwakisa PS (2004). Biodiversity (variation) in disease resistance and in pathogens within rural chicken populations. In International Health Netword for Family Poultry (INFD). World Poultry Congress, pp. 8-13. |
|
|
Ministry of Livestock and Fisheries Development (MLFD) (2015). Tanzania livestock modernization initiative. Dar es Salaam, Tanzania: Ministry of Livestock and Fisheries Development. |
|
|
Mpenda FN, Schilling MA, Campbell Z, Mngumi EB, Buza J (2019). The genetic diversity of local African chickens: A potential for selection of chickens resistant to viral infections. The Journal of Applied Poultry Research 28(1):1-12. |
|
|
Msoffe P, Minga U, Olsen J, Yongolo M, Juul-Madsen HR, Gwakisa P, Mtambo M (2001). Phenotypes including immunocompetence in scavenging local chicken ecotypes in Tanzania. Tropical Animal Health Production 33:341-354. |
|
|
Mtileni B, Muchadeyi F, Maiwashe A, Phitsane P, Halimani T, Chimonyo MK, Dzama K (2009). Characterisation of production systems for indigenous chicken genetic resources of South Africa. Applied Animal Husbandry for Rural Development 2:18-22. |
|
|
Muchadeyi FC, Dzomba EF (2017). Genomics tools for the characterization of genetic adaptation of low input extensively raised chickens. Poultry Science11:211-229. |
|
|
Munisi GH Katule AM, Mbaga SH (2015). Comparative growth and livability performance of exotic, indigenous chickens and their crosses in Tanzania. Livestock Research for Rural Development 27. |
|
|
Pevzner I, Stone H, Nordskog A (1981). Immune response and disease resistance in chickens. Selection for high and low titer to Salmonella pullorum antigen. Poultry Science 60:920-926. |
|
|
Sharif A, Ahmad T, Umer M, Rehman A, Hussain Z (2014). Prevention and control of Newcastle disease. International Journal of Agricultural Innovations Research 3(2):454-460. |
|
|
Sharma J, Xie J, Boggess M, Galukande E, Semambo D, Sharma S (2015). Higher weight gain by Kuroiler chickens than indigenous chickens raised under scavenging conditions by rural households in Uganda. Livestock Research for Rural Development 27(9). |
|
|
Tabidi MH, Makkawi A, Mahasin E, Ali AS (2004). Comparative evaluation of haemagglutination inhibition test and enzyme-linked immunosorbent assay for detection of antibodies Against Newcastle disease vaccine in broiler chicks. International Journal Poultry Science 3:668-670. |
|
|
Taha AE, El-Edel MA, El-Lakany HF, Shewita RS (2012). Growth performance and immune response against Newcastle and avian influenza vaccines in Egyptian chicken strains. Global Vetenaria 9:434-440. |
|
|
Team RC (2017). R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing; 2017. ISBN3-900051-07-0 |
|
|
Yakubu A, Ari M (2018). Principal component and discriminant analyses of body weight and conformation traits of Sasso, Kuroiler and indigenous Fulani chickens in Nigeria. Journal of Animal and Plant Science 28(1):46-55. |
|
|
Yune N, Abdela N (2017). Update on epidemiology, diagnosis and control technique of Newcastle Disease. Journal of Veterinary Science and Technology 8:429. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


