Full Length Research Paper

ABSTRACT
![]() Studies have shown that pasture-raised pork is fresher and of higher nutritional value than its conventional counterpart. This study was to determine the effect of grazing systems on meat quality, carcass traits, and on lipid metabolism gene expressions. Control pigs were fed 100% commercial diet. Fifty/fifty (50/50) group was placed on 50% of the diet consumed by control group plus free access to ryegrass-clover pasture. The twenty-five/seventy-five (25/75) group was fed 25% of the diet consumed by the control plus access to free pasture. The overall meat quality (flavor, overall acceptability and carcass traits (marbling, color)) were scored significantly higher (P<0.05) in the 25/75 group than in control or 50/50 group. Back-fat thickness was lower in 25/75 group (P<0.05) than in control and 50/50 group. No differences were observed between the control and 50/50 in meat and carcass qualities. Real-time polymerase chain reaction (PCR) revealed that peroxisome proliferator-activated receptor α (PPARα), peroxisome proliferator-activated receptor γ (PPARγ), lipoprotein lipase (LPL), and sterol-regulatory-element binding protein 2 (SREBP-2) responded differently in muscle and adipose tissues. The results indicated that pasture-based pork production could positively influence lipid metabolism genes important in meat and carcass quality traits, with pasture exposure and feed allowance.
Studies have shown that pasture-raised pork is fresher and of higher nutritional value than its conventional counterpart. This study was to determine the effect of grazing systems on meat quality, carcass traits, and on lipid metabolism gene expressions. Control pigs were fed 100% commercial diet. Fifty/fifty (50/50) group was placed on 50% of the diet consumed by control group plus free access to ryegrass-clover pasture. The twenty-five/seventy-five (25/75) group was fed 25% of the diet consumed by the control plus access to free pasture. The overall meat quality (flavor, overall acceptability and carcass traits (marbling, color)) were scored significantly higher (P<0.05) in the 25/75 group than in control or 50/50 group. Back-fat thickness was lower in 25/75 group (P<0.05) than in control and 50/50 group. No differences were observed between the control and 50/50 in meat and carcass qualities. Real-time polymerase chain reaction (PCR) revealed that peroxisome proliferator-activated receptor α (PPARα), peroxisome proliferator-activated receptor γ (PPARγ), lipoprotein lipase (LPL), and sterol-regulatory-element binding protein 2 (SREBP-2) responded differently in muscle and adipose tissues. The results indicated that pasture-based pork production could positively influence lipid metabolism genes important in meat and carcass quality traits, with pasture exposure and feed allowance.
Key words: Pigs, lipid metabolism, meat quality, real-time polymerase chain reaction (PCR).
INTRODUCTION
Molecular biology approaches using gene expression have been reported that numerous genes are associated with leaner pigs carcasses, meat and carcass quality traits (Albuquerque et al., 2017; Ivanovic et al., 2015; Tous et al., 2014; Álvarez-Rodríguez et al., 2016). Many of these genes have also been found to be expressed during obesity-related diseases (Georgiadi and Kersten, 2012).
Currently, obesity is considered a public health disease. According to the World Health Organization (WHO), the obese population has doubled in number in less than 30 years (Mitchell and Shaw, 2015). Consumers today are more concerned about their health and physical fitness when seeking ways to get a healthier source of protein.
Therefore, the understanding of lipid metabolism in pigs could elucidate mechanisms underlining conditions such as hypertension, type 2 diabetes, cardiovascular diseases, cancer, and atherogenic dyslipidemia. This has led to increased interest in the study of grazing systems on meat quality and genes involved in lipid metabolism.
Animals raised in outdoor systems have been shown to contain higher intramuscular fat (IMF) (Högberg, et al., 2004), color (Estévez et al., 2003; Fraser et al., 2009)and leaner carcasses (Álvarez-Rodríguez et al., 2016; Galvão et al., 2006)than animals in confinement. The content of polyunsaturated fatty acids (PUFA) content on pastures, primarily essential linolenic acid (C18:3), and linoleic acid (C18:2) has been associated with pork of high nutritional value (Ventanas et al., 2006; Kim et al., 2009; Nilzén et al., 2001; Edwards, 2005).
However, the impact of grazing systems on genes involved in lipid metabolism is still poorly studied in swine. Dervishi (2012) reported changes in fatty acids profile of meat products and milk of grazing lambs as well as in genes responsible for stearoyl CoA desaturase (SDC), and other lipid regulatory genes: peroxisome proliferator-activated receptor α (PPARα), eroxisome proliferator-activated receptor γ (PPARγ), and sterol-regulatory-element binding protein 2 (SREBP). PPARs are a family of nuclear receptors acting as ligand-dependent transcription factors (PPARα, PPARδ, and PPARγ) involved in fatty acid synthesis in pigs (Wang, et al., 2016). PPARα regulates cellular fatty acid uptakes, transportation, oxidation, ketogenesis and gluconeogenesis (König et al., 2007). PPARγ is considered an important regulator of adipogenesis (Samulin et al. 2008). The increase in PPARγ activates lipogenic genes such as lipoprotein lipase (LPL) and SREBP-1 and SREBP-2 (Eberlé et al., 2004).
The objective of this study was to determine the effects of a pasture-based pork production system on the expression of genes involved in lipid metabolism (LPL, PPARα, PPARγ, and SREBP-2) and meat quality characteristics in pork.
MATERIALS AND METHODS
The experiment was conducted at Alcorn State University Swine Research and Development Farm, Churchill, Mississippi. The study was carried out during spring and summer (April-June). All the procedures of animal handling and treatment were approved by the Animal Use and Care Committee at Alcorn State University.
A total of 48 finishing pigs from Yorkshire and Hampshire crossbreed sows were randomly assigned to the three experimental groups. The commercial ration used for the experiment met the National Research Council (1998) requirements. The diet contains 15.5% CP of 12.04 KJ of energy. The animals were assigned into six groups (8 animals each). The control groups and its replica received a finishing diet ad libitum. The rest of the groups were raised under pasture conditions and supplemented with a 50/50 and 25/75 of the control diet/pasture combination, respectively.
The control groups were housed indoors on concrete slatted-floor pens (2.1 × 3.6 m). Two pens were used to allocate 8 pigs each, approximately 1.2 m2 per pig. The groups 50/50 and 25/75 were placed on a pasture fenced area of approximately 16200 m2 divided in 8 paddocks (about 1300 to 1600 m2 each). The pigs at the site were rotated among different paddocks and fed with a mixed pasture of ryegrass and red clover. A corrugated zinc hoop (4×3×2m) and an automatic refill water supply were established for each group. The animals were weighed weekly, body weights and feed intake recorded for later use in estimating growth performance.
Initial body weight (IBW), final body weight (FBW), average daily gain (ADG), average feed intake (AFI) and feed conversion ratio (FCR) were evaluated. Pigs were slaughtered at a FBW of approximately 100 kg in Southern University’s Agricultural Research and Extension Center (SUAREC) Meat Laboratory. The dressing percent was obtained using the final body weight and hot carcass weight. The carcass length was measured from the forward edge of the first rib to the forward edge of the aitch bone. Backfat thickness was measured at the 10th rib at 7 cm off midline at last rib of the split carcass with a Destron PG-100 fiber-optic probe. The estimated lean yield was calculated using the following formula: Estimated lean yield (ELY) = 55.07 - (0.377 × Fat depth) + (0.052 × Muscle depth). Measures of loin muscle meat quality include color, loin muscle score, loin muscle marbling score, and loin muscle firmness score. The major primal cuts from the carcass left side (shoulder, loin, ham, and belly) were trimmed to remove the fat and the weights were recorded to obtain net yield. The color was determined from chop cuts at the last rib level using a Chromameter (C-300; Minolta, Dietikon, Switzerland) and illuminant D65.
Thirty-three panelists were recruited to perform meat quality evaluation. Roasted pork samples from the same muscle were given to the panelist from each treatment. Additionally, red lights were used in the place to disguise color differences. The panelist evaluated the pork using the Five-Point Hedonic Scale (1 as extremely unpleasant and 5 extremely acceptable).
Sample collection and RNA processing
After slaughter, samples of muscle and adipose tissue (about 500 to 1,000 mg) were collected and immediately rinsed with DEPC-treated water, then snap-frozen in liquid nitrogen, and then transferred to a -80°C freezer for future lipid metabolism analysis. Total RNA was extracted from the frozen tissue sample using Trizol (Sigma cat# T-9424). Homogenization of the samples was done by Quiagen-tissue laser. The RNA precipitation was done following a standard protocol (Thermo Scientific). Purity and concentration were analyzed by using a Nanodrop ND-2000 spectrophotometer. The integrity of the samples was examined by gel electrophoresis.
Reverse transcription polymerase reaction and real-time polymerase chain reaction
RNA was treated with DNAase I. RT reaction was done using SuperScript® III Reverse Transcriptase. Then it was incubated at 50°C for 50 min. The reaction was inactivated by heating at 70°C for 15 min. The primers used were PPARα, PPARγ, SREBP, and LPL (Table 1). The reaction was developed using a modified protocol of Kary Mullis (1983). PCR reactions were run in an ICycler (Bio-Rad, Hercules, CA, USA) using PerfeCTa® SYBR® Green FastMix® for iQ (Quanta Biosciences, Gaithersburg, MD). The cDNA Cycling conditions were as follows: 3 min at 95°C, followed by 40 cycles of 15 s at 95°C, 1 min at 60°C; then followed by 80 cycles of 10 s at 55°C with increasing setpoint temperature after cycle 2 by 0.5°. Changes in gene expression were conducted according to the protocol described by Schmittgen and Livak (2008).
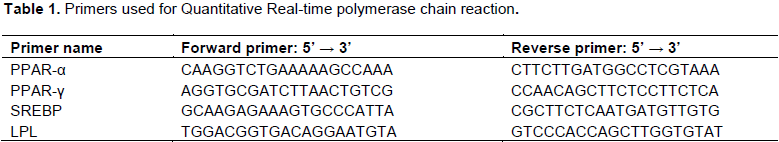
Statistical analysis
Statistical Analytic Software (SAS, 2013) was used to analyze the weights of the pigs and the relative expression levels of gene expression. Mean squares were determined using the general linear model (GLM) and mean separations were done using the Duncan multiple range tests (SAS Ins., 1985). Excel (Microsoft office, 2010) was used to analyze the relative expression of the genes.
The relative quantities of mRNA expression were normalized to the relative quantities of a housekeeping gene and these normalized ratios were used for the quantitative determination of gene expression.
RESULTS AND DISCUSSION
Growth performance
The rate of growth of the pigs across the treatment was evident (Table 2). The different treatment groups were not varied in body weight since all the groups were slaughtered at similar market weight. However, average daily gain and feed conversion ratio differed (P <0.05) reaching the market weight 30 and 60 days earlier than 50 and 25% of commercial diet group, respectively. Similar results were observed by Heyer et al. (2006) who noted lower performance in crosses of Duroc × Large White and Swedish Landrace × Large White when compared with indoor animals. Also, Hansen et al. (2006)reported a reduced average daily gain of the pigs under organic systems with restricted concentrate feeding. In contrast, Lebret and Guillard (2005) reported that outdoor sows showed a higher final body weight than the indoor sows.
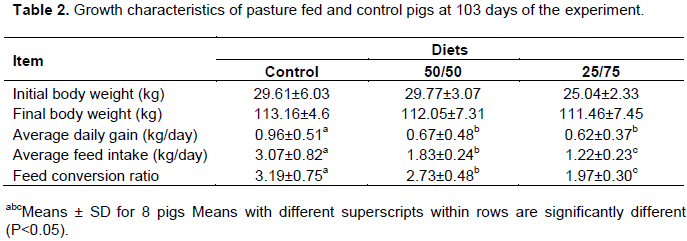
The reduced feed intake (P<0.05) in pigs from diets 50/50 and 25/75 groups was significantly lower than those of the controls due to the dietary fiber levels. Kerr and Shurson (2013) and Noblet ans Le Goff (2001) reported that fibrous diets affected energy and nutrient digestibility and are associated with energy losses as methane, hydrogen, and fermentation heat. Thus, the significantly lower FCR (P<0.05) observed on pasture fed animals were expected as long as the growth performance depends on the dietary energy-protein balance. Similar results were observed by Smit and Beltrana (2017) in pigs fed a diet with Camelina sativa cake inclusions, although a decrease in feed intake was observed, there was a significant increase of n-3fatty acids in adipose tissues.
Carcass and meat quality characteristics
As shown in Table 3, the diet had no effect (P>0.05) on primal cuts (length, shoulder, leg, rib). Nevertheless, dressing percentage and loin were significantly (P<0.05) higher in the control group than in carcasses from pigs raised on pasture. Similarly, Rey et al. (2006), reported lower carcass weight and yield from Iberian pigs raised on pasture in comparison to indoor animals. Contrary to our findings, Lebret and Guillard (2005) and Gentry et al. (2002) found that hot carcass weight, cold carcass weight, dressing percentage, and loin eye area were higher under organic feeding systems.
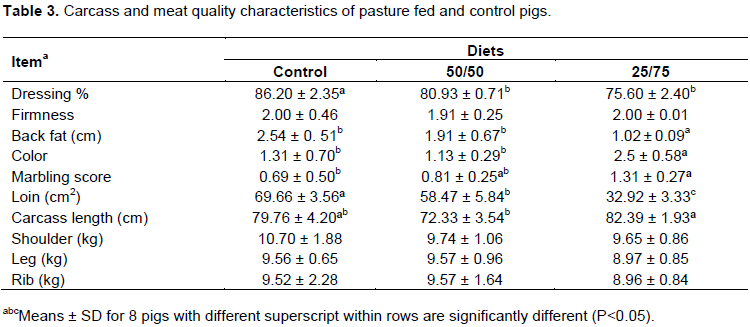
In agreement with Ahmed et al. (2016)who supplemented the diets of grower-finisher pigs using pomegranate, there was a significant reduction in backfat thickness while increasing the forage intake and restriction of feed. In the present study, the animals fed 25% of commercial diet developed leaner carcasses (P <0.05) with back fat results of 2.48, 2.22 and 1.01 cm for the control, 50/50 and 25/75 groups, respectively.Strudsholm and Hermansen (2005) also reported thicker back fat levels 1.7, 1.65 and 1.47 cm in control, outdoor with ad-libitum and outdoor with restricted intake, respectively. In addition, the significantly higher (P<0.05) intensity in color of pork from the 25/75 group consuming greater forage and potentially increasing the tocopherol levels was observed. Consequently, myoglobin levels and a number of oxidative fibers might have improved too (Högberg et al., 2004; Rodríguez-Estévez et al., 2009). These findings were also confirmed byGentry et al. (2002), Tartrakoon et al. (2012) and Álvarez-Rodríguez et al. (2016). However, Lebret and Guillard (2005) reported decreased in redness in Triceps brachii and bacon, whereas Hansen et al. (2006) observed no influence of the production system on meat color. The marbling score, also known as intramuscular fat content (IMF) presented a gradual tendency to increase in 25/75 (P>0.065). A possible explanation may be due to the monounsaturated fatty acids (MUFA) and polyunsaturated fatty acids (PUFA) in meat, commonly as a result of high content of linolenic acid (C18: 3ω-3) and linoleic acid (C18: 2ω-6) present in forages (Álvarez-Rodríguez et al., 2016; Nilzén et al., 2001). Moreover, marbling in outdoor animals has been associated with an increased protein turnover rate and proteolytic activity in muscle (Edwards, 2005). Lipoprotein lipase (LPL) and indeed, intramuscular fat (IMF) is the main determinant factor influencing sensory attributes of meat (Araújo et al., 2011; Lim et al., 2015). The pork aroma and tenderness was not significantly affected (P>0.05) among the treatments (Table 4). However, a tendency for higher tenderness was observed on meat from 25/75 groups. In addition, flavor and overall acceptability were higher in 25/75 group (P<0.05), while differences were not observed between 50/50 group and the control. These results emphasized the positive effect of PUFA content of the forages in the deposition in IMF. A similar effect of PUFA on pork quality was observed by Tartrakoon et al. (2016) and Nilzén et al. (2001)resulting in higher antioxidant levels and consequently increase oxidative stability. In a previous study, it was not observed that there was an association between PUFA content with changes in the sensory characteristics of pork from Korean native black pigs (Kim et al., 2009b).
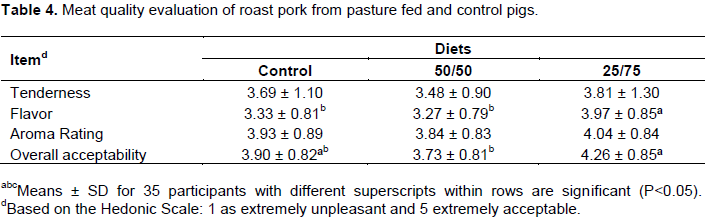
Gene expression levels of LPL, PPARα, PPARγ, and SREBP-2 in muscle and adipose tissue
Forages like clover and ryegrass, are rich in PUFA especially linolenic acid (C18: 3ω-3) and linoleic acid (C18: 2ω-6) (Wyss et al., 2006). As previously mentioned, the effect of PUFAs on meat quality has been widely studied at the molecular level (Ivanovic et al., 2015; Joven et al., 2014; Nilzén et al., 2001). Moreover, new implications of PUFA have been reported to influence genes involved in lipid metabolism and subsequently affect meat quality traits. In previous studies, Trifolium pretense and plant extracts of mulberry, banana, and ginseng were shown to have significant implications in the activation of signaling pathways influencing meat quality traits, principally mediated throughout the PPARs binding-activation (Park et al., 2005; Qiu et al., 2012).
In this study, the pasture-based pork production system was compared with pigs fed on commercial feed to evaluate the meat quality and related lipid gene expression. Real-time PCR was performed to detect gene expression variation of PPARα, PPARγ, LPL, and SREBP-2. The relative expression levels of these genes are presented in Figures 1 and 2. The expression levels differed significantly (P<0.05) among treatments and between tissue sample types. LPL, PPARα, PPARγ and SREBP-2 expression in adipose tissue were highly stimulated in 50/50 treatment than in the control and 25/75 group. However, LPL, PPARα, and PPARγ in muscle were significantly increased in 25/75 group (Figures 1 and 2).
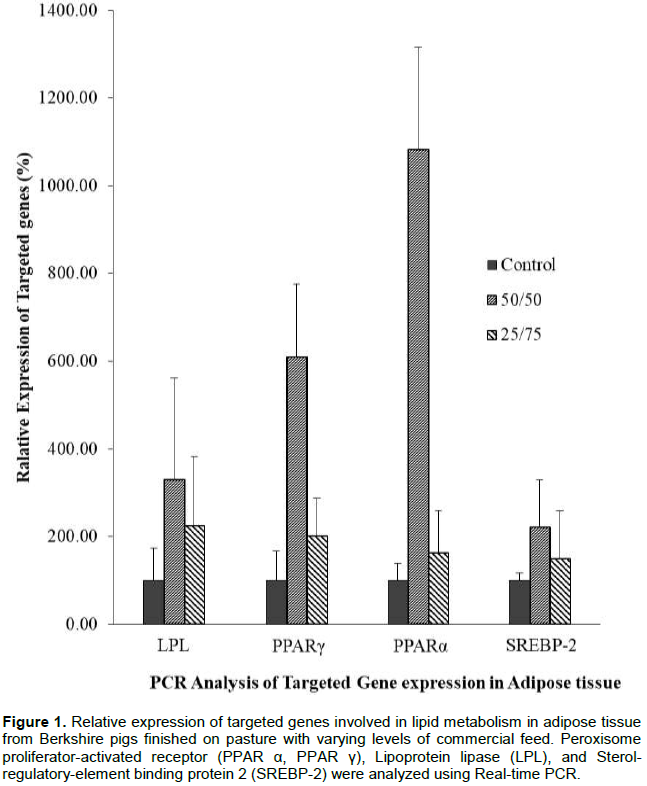
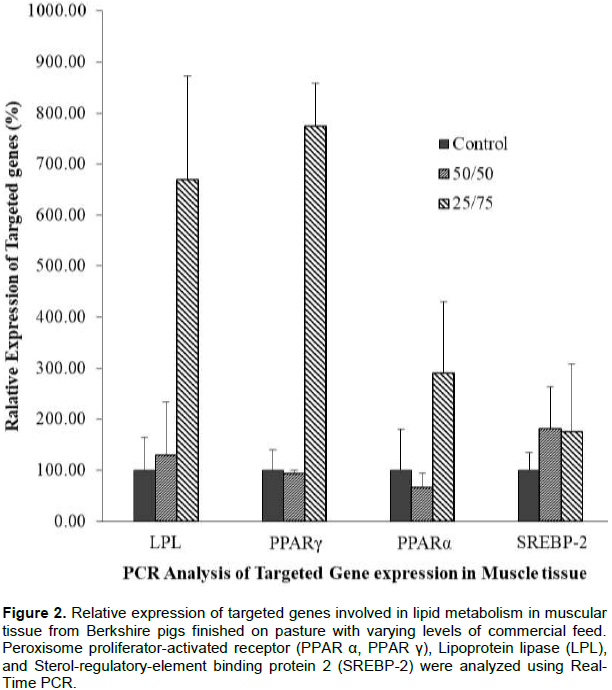
Peroxisome proliferator-activated receptor alpha (PPAR-alpha), also known as NR1C1, is a nuclear receptor protein which in human is encoded by the PPARα gene. PPARα is activated under conditions of energy deprivation and is necessary for the process of ketogenesis. Therefore, the increased relative expression level of PPARα (P<0.05) in pasture animals could be related to a metabolic fasting process due to the reduced energy obtained from forages (Yang et al., 2013). Likewise, PPARs signaling pathway is influential in meat quality traits (He et al., 2013; Wang et al., 2016). It is known that activation of PPARα promotes uptake, utilization and catabolism of fatty acids (Vahmani et al., 2014). The leaner carcasses (P<0.05) on free-rearing systems might be explained by the transport and utilization of fatty acids to catabolic routes. Luci et al. (2007), in their study using clofibrate, also confirmed an increase in PPARα relative expression in adipose tissue, suggesting a direct relationship between the beta-oxidation capacity of adipose tissue and PPARα expression.
On the other hand, a link between PPARα and sterol regulatory element-binding protein-2 (SREBP-2) has been found(König et al., 2009). Sterol regulatory element-binding proteins (SREBPs) are transcription factors that regulate lipid homeostasis. SREBP-2 is related to cholesterol synthesis (Eberlé et al., 2004). Therefore, SREBP-2 and its target genes relative expression could be attenuated by PPARα resulting in lower cholesterol concentration in plasma (König et al., 2007, 2009).
Peroxisome proliferator-activated receptor gamma (PPAR-γ or PPARG), also known as the glitazone receptor, or NR1C3 is a type II nuclear receptor, which in humans is encoded by the PPARγ gene. The relation between IMF content and PPARγ activation is probably due to its important role in reducing glucose concentrations favoring adipogenesis. Lipoprotein lipase (LPL) role is also considered essential in fat deposition (Albuquerque et al., 2017). Marbling score in 25/75 group was significantly higher than in the other two treatments. A possible explanation for increased IMF (marbling score) deposition in animals under higher pasture consumption could be a possible activation of PPARγ gene expression.
CONCLUSIONS
The findings from the present study suggested that a reduction in swine diet by 25 to 50% plus ad-libitum grazing supplementation of rye-grass/red clover can maintain a sustainable pasture pork production of finishing pigs. The intense grazing and reduced rate of gain resulted in upload of lipid metabolic genes in muscle and adipose tissue, reducing backfat thickness while improving meat quality characteristics. This study showed that pasture-based pork production system could up-regulate lipid metabolism genes expression that improved meat quality traits.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
This research was suppported in part by USDA-NIFA Grant #MISX3500. The research was completed in part by Nnamdi Okoli as part of the requirement for M.S. in Agricultural Biotechnology.
REFERENCES
|
Ahmed ST, Mun HS, Islam MM, Ko SY, Yang CJ (2016). Effects of dietary natural and fermented herb combination on growth performance, carcass traits and meat quality in grower-finisher pigs. Meat Science 122:7-15. |
|
|
Albuquerque A, Neves JA, Redondeiro M, Laranjo M, Félix MR, Freitas A, Martins JM (2017). Long-term betaine supplementation regulates genes involved in lipid and cholesterol metabolism of two muscles from an obese pig breed. Meat Science 124:25-33. |
|
|
Álvarez-Rodríguez J, Villalba D, Cubiló D, Babot DM, Tor M (2016). Organic practices and gender are effective strategies to provide healthy pork loin. Journal of Integrative Agriculture 15:608-617. |
|
|
Araújo WAG, Albino LFT, Sakomura NK, Paulino PVR, Campos AM (2011). Meat quality in indoor and outdoor production systems of poultry and swine. Open Journal of Animal Science 1:75-88. |
|
|
Dervishi E, Joy M, Alvarez-Rodriguez J, Serrano M, Calvo J (2012). The forage type (grazing versus hay pasture) fed to ewes and the lamb sex affect fatty acid profile and lipogenic gene expression in the longissimus muscle of suckling lambs. Journal of Animal Science 90:54-66. |
|
|
Eberlé D, Hegarty B, Bossard P, Ferré P, Foufelle F (2004). SREBP transcription factors: Master regulators of lipid homeostasis. Biochimie 86:839-848. |
|
|
Edwards SA (2005). Product quality attributes associated with outdoor pig production. Livestock Production Science 94:5-14. |
|
|
Estévez M, Morcuende D, Cava R (2003). Oxidative and colour changes in meat from three lines of free-range reared Iberian pigs slaughtered at 90 kg live weight and from industrial pig during refrigerated storage. Meat Science 65:1139-1146. |
|
|
Fraser MD, Davies DA, Vale JE, Nute GR, Hallett KG, Richardson RI, Wright IA (2009). Performance and meat quality of native and continental cross steers grazing improved upland pasture or semi-natural rough grazing. Livestock Production Science 123:70-82. |
|
|
Galvão DM, Abreu da Silva M, Borges De Medeiros R, De Saibro C, Pavan MA, Arrighi MA (2006). Effect of different grazing systems on the performance of pigs, in growing and finishing phases, kept on white clover (Trifolium repens L.) pasture. Revista Brasileira de Zootecnia 35:792-796. |
|
|
Gentry JG, McGlone JJ, Miller MF, Blanton JR (2002). Diverse birth and rearing environment effects on pig growth and meat quality. Journal of Animal Science 80:1707–1715. |
|
|
Georgiadi A, Kersten S (2012). Mechanisms of gene regulation by fatty acids. Advances in Nutrition 3:127-134. |
|
|
Hansen L, Claudi-Magnussen C, Jensen S, Andersen H (2006). Effect of organic pig production systems on performance and meat quality. Meat Science 74:605-615. |
|
|
He K, Wang Q, Wang Z, Pan Y (2013). Association study between gene polymorphisms in PPAR signaling pathway and porcine meat quality traits. Mammalian Genome 24:322-331. |
|
|
Högberg A, Pickova J, Stern S, Lundström K, Bylund, A (2004). Fatty acid composition and tocopherol concentrations in muscle of entire male, castrated male and female pigs, reared in an indoor or outdoor housing system. Meat Science 68:659-665. |
|
|
Ivanovic J, Pantic S, Dokmanovic M, Glamoclija N, Markovic R, Janjic J, Baltic, MZ (2015). Effect of Conjugated Linoleic Acids in Pig Nutrition on Quality of Meat. Procedia Food Science 5:105-108. |
|
|
Joven M, Pintos E, Latorre MA, Suárez-Belloch J, Guada JA, Fondevila M (2014). Effect of replacing barley by increasing levels of olive cake in the diet of finishing pigs: Growth performances, digestibility, carcass, meat and fat quality. Animal Feed Science and Technology 197:185-193. |
|
|
Kerr BJ, Shurson GC (2013). Strategies to improve fiber utilization in swine. Journal of Animal Science and Biotechnology 4:11. |
|
|
Kim DH, Seong PN, Cho SH, Kim JH, Lee J M, Jo C, Lim DG (2009a). Fatty acid composition and meat quality traits of organically reared Korean native black pigs. Livestock Science. 120:96-102. |
|
|
Kim DH, Seong PN, Cho SH, Kim JH, Lee JM, Jo C, Lim DG (2009b). Fatty acid composition and meat quality traits of organically reared Korean native black pigs. Livestock Science 120:96-102. |
|
|
König B, Koch A, Spielmann J, Hilgenfeld C, Hirche F, Stangl GI, Eder K (2009). Activation of PPARα and PPARγ reduces triacylglycerol synthesis in rat hepatoma cells by reduction of nuclear SREBP-1. European Journal of Pharmacology 605:23-30. |
|
|
König B, Koch A, Spielmann J, Hilgenfeld C, Stangl GI, Eder K (2007). Activation of PPARα lowers synthesis and concentration of cholesterol by reduction of nuclear SREBP-2. Biochemical Pharmacology 73(4):574-585. |
|
|
Lebret B, Guillard AS (2005). Outdoor rearing of cull sows: Effects on carcass, tissue composition, and meat quality. Meat Science 70:247-257. |
|
|
Lim D, Chai H, Lee S, Cho Y, Choi J, Kim N (2015). Gene Expression Patterns Associated with Peroxisome Proliferator-activated Receptor (PPAR) Signaling in the Longissimus dorsi of Hanwoo (Korean Cattle). Asian-Australian Journal of Animal Science 28(8):1075-1083. |
|
|
Luci S, Giemsa B, Kluge H, Eder K (2007). Clofibrate causes an upregulation of PPAR-γ target genes but does not alter expression of S REBP target genes in liver and adipose tissue of pigs. Lipids, pp. 16-88. |
|
|
Mitchell S, Shaw D (2015). The worldwide epidemic of female obesity. Best Practice and Research: Clinical Obstetrics and Gynaecology 29(3):289-299. |
|
|
Nilzén V, Babol J, Dutta P, Lundeheim N, Enfält AC, Lundström K (2001). Free range rearing of pigs with access to pasture grazing — effect on fatty acid composition and lipid oxidation products. Meat Science 58(3):267-275. |
|
|
Noblet J, Le Goff G (2001). Effect of dietary fibre on the energy value of feeds for pigs. Animal Feed Science and Technology 90(1-2):35-52. |
|
|
Park MY, Lee KS, Sung MK (2005). Effects of dietary mulberry, Korean red ginseng, and banaba on glucose homeostasis in relation to PPAR-??, PPAR-??, and LPL mRNA expressions. Life Sciences 77(26). |
|
|
Qiu L, Chen T, Zhong F, Hong Y, Chen L, Ye H (2012). Red clover extract exerts antidiabetic and hypolipidemic effects in db/db mice. Experimental and Therapeutic Medicine 4(4):699-704. |
|
|
Rey AI, Daza A, López-Carrasco, C, López-Bote CJ (2006). Feeding Iberian pigs with acorns and grass in either free-range or confinement affects the carcass characteristics and fatty acids and tocopherols accumulation in Longissimus dorsi muscle and backfat. Meat Science 73(1):66-74. |
|
|
Rodríguez-Estévez V, García A, Pe-a, F, Gómez AG (2009). Foraging of Iberian fattening pigs grazing natural pasture in the dehesa. Livestock Science 120:135-143. |
|
|
Samulin J, Berget I, Lien S, Sundvold H (2008). Differential gene expression of fatty acid binding proteins during porcine adipogenesis. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology 151:147-152. |
|
|
Smit MN, Beltrana E (2016). Increasing dietary inclusions of camelina cake fed to pigs from weaning to slaughter: Safety, growth performance, carcass traits, and n-3 enrichment of pork. Journal of Animal Science 95:2952-2967. |
|
|
Strudsholm K, Hermansen JE (2005). Performance and carcass quality of fully or partly outdoor reared pigs in organic production. Livestock Production Science 96:261-268. |
|
|
Tartrakoon W, Tartrakoon T, Kitsupee N (2016). Effects of the ratio of unsaturated fatty acid to saturated fatty acid on the growth performance, carcass and meat quality of finishing pigs. Animal Nutrition 2:79-85. |
|
|
Tous N, Lizardo R, Theil PK, Vilà B, Gispert M, Font-i-Furnols M, Esteve-Garcia E (2014). Effect of vitamin A depletion on fat deposition in finishing pigs, intramuscular fat content and gene expression in the longissimus muscle. Livestock Science16(7):392-399. |
|
|
Vahmani P, Glover KE, Fredeen AH (2014). Effects of pasture versus confinement and marine oil supplementation on the expression of genes involved in lipid metabolism in mammary, liver, and adipose tissues of lactating dairy cows. Journal of Dairy Science 97:4174-4183. |
|
|
Ventanas S, Estevez M, Tejeda J, Ruiz J (2006). Protein and lipid oxidation in Longissimus dorsi and dry cured loin from Iberian pigs as affected by crossbreeding and diet. Meat Science 72:647-655. |
|
|
Wang W, Xue W, Xu X, Jin B, Zhang X (2016). Correlations of genes expression in PPAR signalling pathway with porcine meat quality traits. Czech Animal Science Journal 61:333-339. |
|
|
Wyss U, Morel I, Collomb M (2006). Fatty acid content of three grass/clover mixtures. Grassland Productivity 11:348-350. |
|
|
Yang H, Li F, Xiong X, Kong X, Zhang B, Yuan X, Óvilo C (2013). Identification and characterization of novel peroxisome proliferator-activated receptor-gamma (PPAR-gamma) transcriptional variants in pig and human. Livestock Science 24:322-331. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


