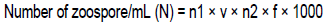ABSTRACT
Black pod is an important fungal disease of Theobroma cacao L. which causes significant yield losses in the field and consequently, inhibits the growth and development of the plant. For this purpose, the objective of this work was to evaluate the impact of phosphates solubilizing bacteria on growth of cocoa-tree and the biocontrol of Phytophthora megakarya, which is the most aggressive agent of black pod disease. Five strains of phosphate solubilizing bacteria (PSB) were characterized on petri dishes and in liquid media in order to determine their aptitude to solubilize different types of rock phosphate. The aptitude of the strains to inhibit P. megakarya was been evaluated on discs of leaves under in vitro and in vivo culture. The results shown that, Pantoea sp. (63B) with index of solubilization (IS) =5.5 and Enterobacter sp. (196B) with IS = 434.364 are the strains able to solubilize the five different types of rock phosphates used. Moreover, the antagonism test shown that Pantoea sp. (111B) with 90.35%, presented the greatest percentage of inhibition on plate. Bacillus sp. (104B) with 0.5 for the tolerant hybrid genotype and 1.75 for the sensitive hybrid genotype and Pantoea sp. (111B) with 1 for the tolerant hybrid genotype and 1.75 for the sensitive hybrid genotype are the highest foliar sensitivity reduction. In addition, the growth of cocoa tree has been very improved by the strain Enterobacter sp. (196B) for the tolerant hybrid genotype which had increased the plant high up to 50%, 35% for the number of leaves, 38.88% for the stem base diameter and 67.37% for the total dry mass compared to the negative control which did not receive microorganisms. With regard to the hybrids of sensitive cocoa-tree, Pantoea sp. 63B involves an increase of 43.28% of plant high and of 61.11% of the total dry mass, Enterobacter sp. (196B) involves an increase of 22.05% of number of leaves and 31.76% for the stem base diameter compared to the negative control. These strains can therefore be recommended to farmers in order to improve cocoa cultivation in the field.
Keys words: Rock phosphates, Phytophthora megakarya, biocontrol, black pod disease, Thebroma cacao.
Cocoa tree (Theobroma cacao L.) represents one of the major crops originating from tropical rainforests of Central and South America (Pokou et al., 2019). The seeds of this plant have an undeniable industrial and nutritional value and their commercialization constitutes a source of income for many peasants of the producing countries (Anushka and Dunwell, 2018; Morrissey et al., 2019). With an annual amount of the exchanges estimated at approximately 10 billion dollars, cocoa constitutes, according to the Belgian Agency of Development “Trade for Development centers”, the third world food market (Anonymous, 2014). The economy of Cameroon is mainly based on agriculture and the cocoa occupies a choice place among the agricultural products (Etoa, 2009). Cacao is one of the principal cultures of income with a production estimated at 225 000 metric tons in 2012 to 2013 and therefore the fifth world producer (Mfegue, 2012; ICCO, 2013).
In Cameroon, whatever this plant is economically important, its culture is subjected to the low productivity and several causes explain poor yield of the cocoa production: weak fertility of the soils, related to an unavailability of the assimilable phosphorus due to his precipitation by the mineral elements like iron and aluminum for acidic soils or calcium for alkaline soils (Fankem et al., 2008). Another fact is the black pod disease of cocoa tree (Ndoumbe-Nkeng, 2002). This fungal disease causes production losses estimated at nearly 30% on a worldwide scale (Takam, 2011). In Cameroon, these losses can reach 90 to 100% according to the area, the cultivar and the environmental conditions, in absence of treatments of plant (Ndoumbe-Nkeng et al., 2004; Nyadanu et al., 2012).
The means of fight developed by agricultural research to mitigate these constraints are based mainly on the application of chemical products (chemical fertilizers and fungicides). This solution does not answer inevitably any more waiting of the producer, who is in front of the incapacity to implement them, the cost of equipment and the financial means plant health product often exceeding his purchasing power. In addition, the requirements of the international market in terms of quality of the cocoa, the environmental concerns, the health of the consumers, are as many elements which do not support the use of the chemical products (Kébé et al., 2009). Moreover, the attention of these countries is carried more and more towards an “organic” production.
Therefore, agriculture must be directed towards durable cultivation systems with weak inputs. The use of plant growth promoting microorganisms (PGPM) and biological control agents are ways to exploit (BCA) (Nwaga et al., 2000; Tyler et al., 2008). PGPM like phosphate solubilizing bacteria (PSB) are able to ensure the growth of the plant by making available phosphorus and control the effect of phytopathogen. Introduced in the soil, at the opportune moment and the desired place, microorganisms judiciously selected according to their production characteristic of enzyme like bacteriocin, organic acids and others can exert a stimulating effect on protection and the growth of the plants (Robin, 2012).
In spite of many studies made on the cocoa-tree in Cameroon, none was interest under investigation of the microorganisms solubilizing phosphates like agent’s promoters growth of the cocoa-tree via phosphate solubilization and biocontrol of Phytophthora megakarya Brasier and Griffin.
Thereby, the present study aims to evaluate the inoculation effect of strains of PSB on the growth of the cocoa-tree in greenhouse and on the other hand, to characterize their aptitude to inhibit P. megakarya in vitro and in vivo conditions on leaves of cocoa-tree.
Aptitudes of bacteria to solubilize rock phosphates from different origins on plates and in liquid culture
Microorganisms
Five different strains from the strain bank of the Laboratory of Plant Biotechnology of the Faculty of Science of the University of Douala have been used. Pantoea sp. (63B), Klebsiella sp. (113B) and Enterobacter sp. (143B) from agro ecological zone I of Cameroon; Enterobacter sp. (79B) and Enterobacter sp. (196B), from agro ecological zone III of Cameroon. Five rock phosphates from different origins with mineral composition known (Table 1) have been used. To get rid of their soluble fractions, the different rock phosphates were washed 4 times with warm water following the cycle: 1 h to 24 h to 1 h to 24 h. They were then dried at 60°C until complete evaporation.
Preparation and evaluation of the concentration of the inoculum
The inoculum has been prepared by suspension of a pure colony of PSB in 10 mL of nutritive broth sterile having for composition per liter of solution: NaCl 3 g, yeast extract 3 g, peptone 5 g, pH 7 and incubated at 28°C, 150 rpm for 3 days. The evaluation of the concentration of the inoculum was made starting from decimal dilutions of the culture obtained, so 1 mL of culture was diluted in 9 mL of sterile water distilled until the dilution 10-7. One mL of dilution 10-7 was introduced into the Petrie dishes then mixed with approximately 12 mL of nutrient agar. After incubation at 28°C, during 3 to 4 days, the bacterial colonies were counted and the number of Colony Forming Unit (CFU) per ml recorded.
Bacterial rock phosphate solubilizing capacity on petri dishes and quantitative estimation of phosphate solubilization in liquid media
The aptitude of the PSB to solubilize the various types of phosphate was made on petri dish containing the National Botanical Research Institute’s Phosphate growth medium with some modifications (Fankem et al., 2014a) and containing per liter of distilled water: 20 g glucose, 5 g MgCl2·6H2O, 0.25 g MgSO4·7H2O, 0.2 g KCl, 0.1 g (NH4)2SO4, 15 g agar and one rock phosphate type (Algerian RP, Cameroonian RP, Malian RP, Mexico RP and Morocco RP) at 5 g/L plus 0.5% bromocresol green, pH 7.5. Five microliters of each bacterial suspension obtained as described above were transferred onto a single point of compartmented petri dish. The petri dishes were sealed and incubated at 28°C for 5 days and the phosphate solubilization recorded through the halo/yellow zone surrounding the bacterial colony. The index of solubilization (IS) as defined by Edi-Premono et al. (1996) was used as an indicator for the strains efficiency.
Bacteria were tested in liquid media to assess their capability in releasing phosphorus from insoluble rock phosphate sources. Fifty mL NBRIP medium were distributed into 250 mL Erlenmeyer flasks, individual rock phosphate types (Algerian RP, Cameroonian RP, Malian RP, Mexico RP and Morocco RP) were added to the medium at the concentration of 5 g/L and the pH adjusted to 7.5. After sterilization and cooling, 200 μL bacterial suspensions of 1 to 1.7x109 CFU/mL were used to inoculate flasks containing the different rock phosphates. Each treatment was performed in triplicate and non inoculated flasks supplemented with the different rock phosphates supplied with 200 μL 0.85% sterile NaCl served as controls. Incubation was made at 28°C, 150 rpm for 7 days. At end of the incubation time and in all cases, the cultures were transferred into sterile falcon tubes, centrifuged at 10.000 g for 10 minutes at 4°C, a part of the supernatant taken for pH measurement and another part for P determination following the method used by Fankem (2007).
Ability of PSB to inhibit the growth of Phytophthora megakarya on Petri dishes
Isolation and identification of Phytophthora megakarya from cocoa pod
The surface of cocoa pod was washed beforehand with tap water, and then underwent a series of disinfection with ethanol 95% during 30 s, sodium hypochlorite 10% during 2 min, then in ethanol 75% during 2 min to eliminate the microorganisms present on surface. Cocoa pods are then rinsed three times at sterile distilled water to eliminate the traces from disinfecting (Arnold, 1999; Evans et al., 2003; Rubini et al., 2005). A piece of infected cocoa pod was taken and deposited on a culture medium containing per liter of solution: 70 g of pea, 15 g of agar (Hugenin and Boccas, 1971). In this medium, the thallus develops in absence of bacteria. After the thallus was formed, a mycelia fragment was taken on the face of growth of the culture and transferred in the Perti disches on the same medium. Incubation was carried out in the darkness at 26°C during 4 to 5 days.
Test of antagonism between PSB - P. megakarya on Petri diches
The test of confrontation PSB - P. megakarya was carried out on Petri diches containing a medium having for composition per liter of solution: NaCl 3 g, yeast extract 3 g, peptone 5 g, agar 15 g and pea 70 g. It should be noted that the strains of bacteria used was also come from the strain bank of the Laboratory of Vegetable Biotechnology of the Faculty of Science of University of Douala. Their aptitude to solibilize phosphate had been tested before on soluble phosphates like Ca3 (PO4)2, AlPO4·H2O, FePO4·2H2O. Thus, a mycelial fragment of 8 mm in diameter is taken from the periphery of the culture of the mushroom, and then deposited in the center of the Petri dish. At a distance of 2 cm from the mycelial fragment, a strain of MSP is seeded as a line all around the fungus. The control Petri dish only received the mycelial fragment. Each of the bacterial strains (Pseudomonas sp. 1B, Pantoea sp. 22B, Burkholderia sp. 36B, Bacillus sp. 104B, Pantoea sp. 111B, Microbacterium sp. 130B, Pseudomonas sp. 179B, Enterobacter sp. 196B) was confronted with the P. megakarya at a rate of three repetitions; incubation was carried out in the darkness at 26°C. Twenty-four hours after the setting in culture, the mycelium growth was measured daily until the full one with control Petri dish. Thus the percentage of inhibition of the mycelium growth of P. megakarya was calculated according to the following formula: % inhibition = [(R control - R test)/R control] x100 with R = diameter (Wang et al., 2002). It is should be noted that to start from 20% one will speak about inhibition. The bacteria which had an inhibiting effect against P. megakarya were selected for the test on the foliar discs.
Greenhouse trials with phosphate-solubilizing bacteria on the cocoa-tree
A ground of a field in fallow was collected on the campus of the Faculty of Science of the University of Douala (Cameroon) and was used in this experiment. The physical characteristics of this soil were: clay 16.39%, Fine silt, 1.94%; Coarse silt, 1.67%; Fine sand, 31.24%; Coarse sand, 47.76% and chemical: pH H2O, 4.93; PH KCl, 4.34; Moisture, 2.99%; organic material, 31.74 g; organic C, 18.41 g/kg; total N, 2.37 g/kg; Ca2+, 3.57 molc/kg; Mg2+, 0.21 molc/kg; K+ , 0.05 molc/kg; Na+, 0.08 molc/kg; CEC, 4.34 molc/kg; Zn, 2.76 mg/kg; Cu, 853.07 mg/kg; Mn, 155.53 mg/kg; Pb, 9.48 mg/kg; Cr, 39.00 mg/kg; Fe, 714.81 mg/kg; Cd, 0.03 mg/kg; Ni, 8.07 mg/kg; assimilable phosphorus, 9.89 mg/kg.
To conclude this test in greenhouse, five microbial treatments (Pantoea sp. 63B, Enterobacter sp. 79B, Klebsiella sp. 113B, Enterobacter sp. 143B, Enterobacter sp. 196B), a positive control (+) having received soluble phosphorus and a negative control (-) having received rock phosphate were highlighted. This device was carried out in double because two hybrids of cocoa were used namely: SNK64 × UPA143 (hybrid sensitive) and UPA143 × T72/501 (hybrid tolerant) coming from Barombi-Kang (South-West, Cameroon). This fact the treatments for each hybrid were carried out in four repetitions from where a total of experimental units.
The experiment was made in the plastic bags of 2 L, the control (-) received 0.8 g of rock phosphate of Cameroon (C= 233.33 mgP/kg of ground) and the control (+) received soluble phosphorus 1.56 g. the days beforehand, each treatment was watered. The next days, each bag received a seed of cocoa tree and inoculated with 1 mL of suitable PSB having a concentration of 1-3.109 CFU/mL approximately. The bags were then watered every day with 80 to 100 mL of water. A second inoculation was carried out three weeks after the first in order to increase the microbial population.
The parameters of growth to knowing, stem base diameter, number of leaves and plant high of each seedling were evaluated each month as from the second month. At harvest (6 months after sowing), the total dry weight, aerial parts of the stem and roots were weighed using a balance (KERN EMB 600-2).
Incidence of the inoculation of P. megakarya on cocoa leaves under PSB control
Preparation and evaluation of the concentration of zoospores of P. megakarya
The inoculation of P. megakarya consists of a suspension of zoospores. It was obtained starting from the lesions made on surface of cocoa pod. Mycelium fragments of P. megakarya in culture on petri dishes were taken and deposited on the lesions. These lesions were covered using sterile cotton soaked with sterile distilled water. The whitish down consisting of sporocysts that will produce and release zoospores appeared on the surface of the pod was scraped and rinsed with 40 mL of sterile distilled water. Incubation was made less than 12 h photoperiod during one week then placed during 15 min at a temperature of 4°C. The culture was finally exposed in the light of an incandescent lamp during 45 min. The suspension of zoospores obtained was counted using a cell of Malassez and was adjusted with the concentration of 3.105 zoospores/mL (Nyassé et al., 1995) in order to carry out inoculations on the disc of leaves.
The number of zoospore is estimated according to the relation:
with:
n1= number of zoospore counted; v= volume of a rectangle;
n2= number of counted rectangle; f= factor of dilution.
Confrontation test between PSB - P. megakarya on foliar disc of cocoa-tree
The test was carried out on foliar discs of cocoa-tree and consists in measuring the foliar sensitivity to P. megakarya according to a scale which varies from 0 (absence of symptom) to 5 (true spot) (Blaha et al., 2000).
The foliar discs were taken on seedling of two hybrid families of cocoa-tree whose reaction to Phytophthora is known. Thus, the sensitive hybrid (SNK64 × UPA143) and the tolerant hybrid (UPA143 × T72/501) were tested.
Thus, 15 mm of diameter of leaves were soaked in a bacterial suspension (Pantoea sp. 22B, Bacillus sp. 104B, Pantoea sp. 111B, Microbacterium sp. 130B and Enterobacter sp. 196B) adapted during a minute, and then lay out in plastic dishes on toilet paper soaked with water. Each disc received 10 μL of a suspension of zoospores of P. megakarya of concentration 3.105 zoospores/mL prepared beforehand as describe previously. The controls do not undergo soaking in the bacterial suspension. Incubation was carried out with the darkness with 26°C during 7 days. The results were noted according of the scale of Blaha et al. (2000).
Statistical analyses
Statistical analyses were performed with Sigma plot 12.0. The analysis of variance (ANOVA) was run to find difference between factors and the HSD Turkey test to compare the different treatments.
Aptitudes of the PSB to solubilize rock phosphates from different origins on Petri dishes and in liquid media
Aptitude of the strains to solubilize on Petri dishes
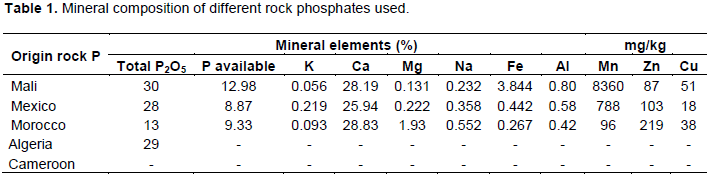
The results obtained (Figure 1) indicated that Pantoea sp. 63B was the one which had more the strong potential of solubilization with an index of solubilization (IS) equal to 5.501 followed by Klebsiella sp.113B which had an IS equal to 5.029; between these two strains there was no significant difference while the strains Enterobacter sp. 79B and Enterobacter sp. 196B presented a weak solubilizer aptitude with IS equal to 3.603 and 3.065 respectively.
Ability to solubilize different rock phosphates
The ability to solubilize differents rock phosphates in solid medium (Figure 2) was evaluated. All the rock phosphates do not have the same capacity of solubilization, the phosphate of Cameroon and Mali were solubilized by the various strains with a IS of 6.911 and 6.543, respectively, followed by the phosphate of Algeria with 4.803 for IS and finally of phosphates of Morocco and Mexico with IS 1.935 and 1.385, respectively. Rock phosphate of Mexico being most recalcitrant.
Capacity of solubilization of the strains compared to various rock phosphates in liquid medium
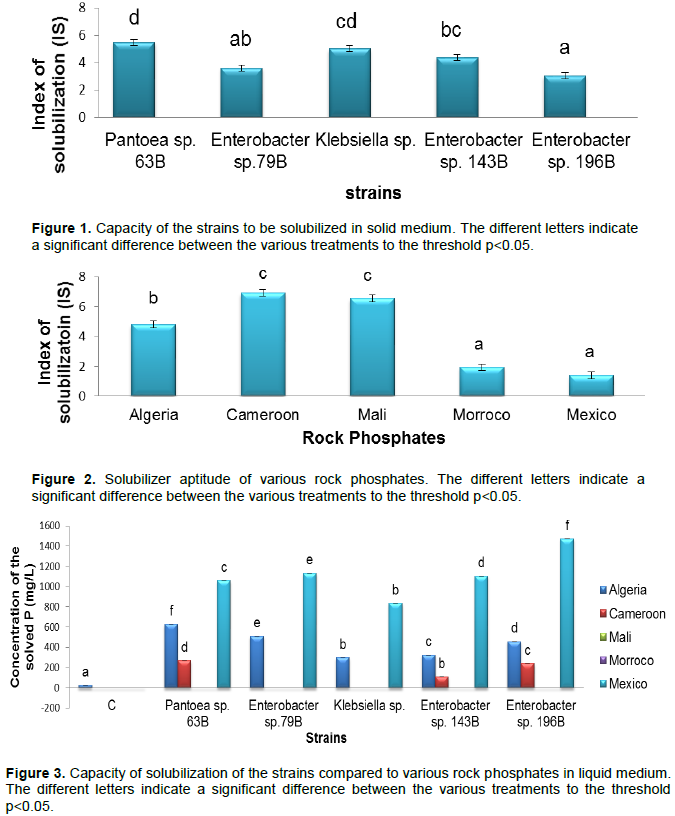
The results obtained (Figure 3) showed that, for rock phosphate of Algeria, Pantoea sp. 63B presented a greater quantity out of phosphorus solubilized with a concentration of 628.182 mg/L. With a concentration of 270.909 mg/L solubilized phosphorus, Pantoea sp. 63B also presented the greatest activity of solubilization for phosphate of Cameroon. As regards rock phosphate of Mexico, Enterobacter sp. 196B presented the greatest solubilized phosphorus concentration which was 1473.636 mg/L. On the other hand, the rock phosphates of Mali and of Morocco were not solubilized.
Aptitude of the strains to solubilize rock phosphates in liquid medium
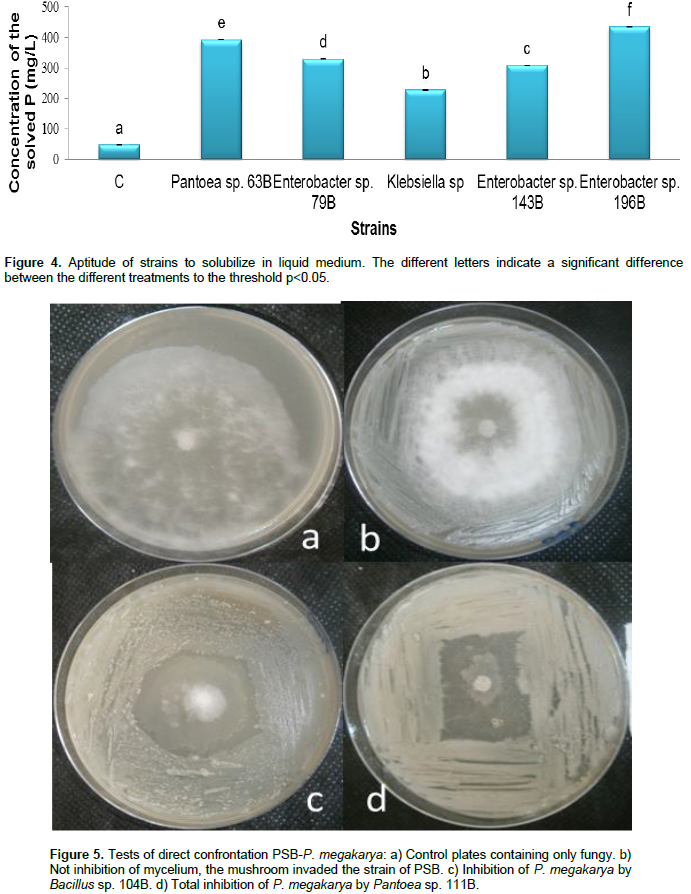
The Figure 4 showed that there was a significant difference between the aptitudes of the strains to solubilize the five types of rock phosphate in liquid medium. With an average concentration equalize with 434.364 mg/L solubilized phosphorus, Enterobacter sp. 196B was the strain which presented a highest solubilizing activity followed by Pantoea sp. 63B, Enterobacter sp. 79B, Enterobacter sp. 143B and Klebsiella sp. 113B had as average respective concentrations 392 mg/L, 328.727 mg/L, 308 mg/L and 227.455 mg/L. Klebsiella sp. 113B was the least efficient strain.
Aptitude of the PSM to inhibit the growth of Phytophthora megakarya on petri dishes
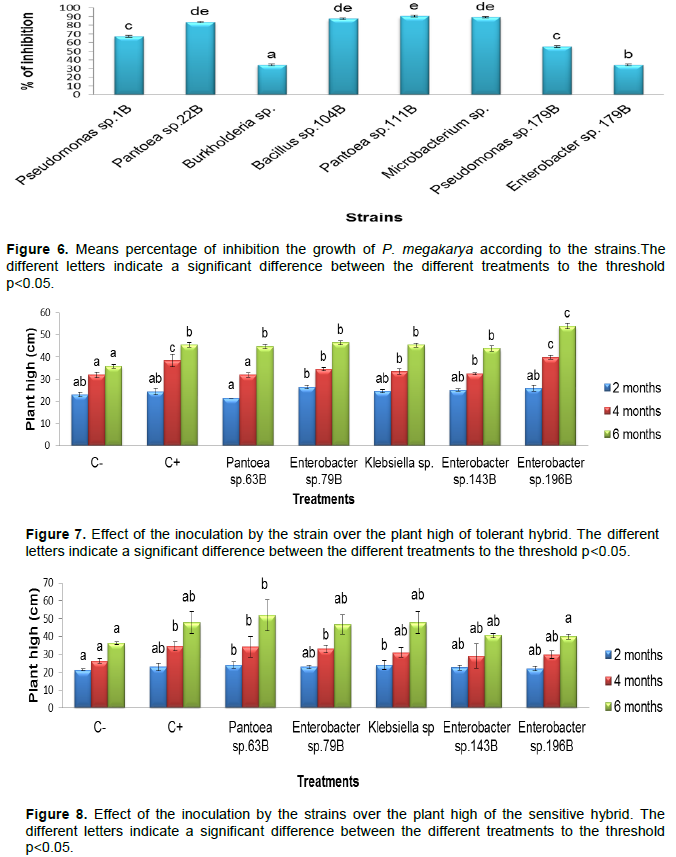
The direct confrontation of eight microbial strains (Pseudomonas sp.1B, Pantoea sp. 22B, Burkholderia sp. 36B, Bacillus sp. 104B, Pantoea sp. 111B, Microbacterium sp. 130B, Pseudomonas sp. 179B and Enterobacter sp. 196B) with P. megakarya, allowed highlighting the aptitude of these strains to inhibit the mycelial growth of pathogens (Figure 5).
The percentage of inhibition of the myelium growth varies from a strain with another (Figure 6). Thus the Pantoea sp. 111B strain presented the highest percentage (90.35%) followed by Microbacterium sp. 130B (89.15%), Bacillus sp. 104B (87.53%), Pantoea sp. 22B (83.68%), Enterobacter sp. 196B (79.91%), Pseudomonas sp.1 B (67.06%), Pseudomonas sp. 179B (55.41%) and Burkholderia sp. 36B (34.11).
On the basis of these result, the most powerful strains were retained for the test on the disc of leaves.
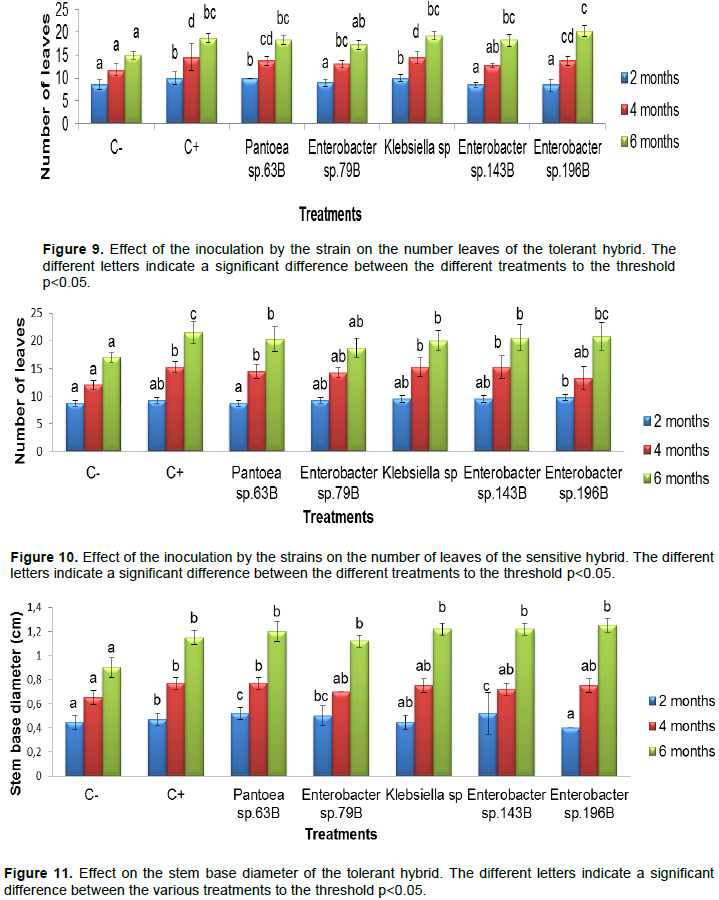
Effect over the plant high of the tolerant hybrid
The results obtained showed a significant difference between the treatments during month (Figure 7). Thus, in two months Enterobacter sp. 79B showed the highest growth which was 26.42 cm. At four months the various treatments were not significant different from each other but Enterobacter sp. 196B presented the highest growth is 38.82 cm. in the sixth month, Enterobacter sp. 196B presented also the greatest value which was 54 cm.
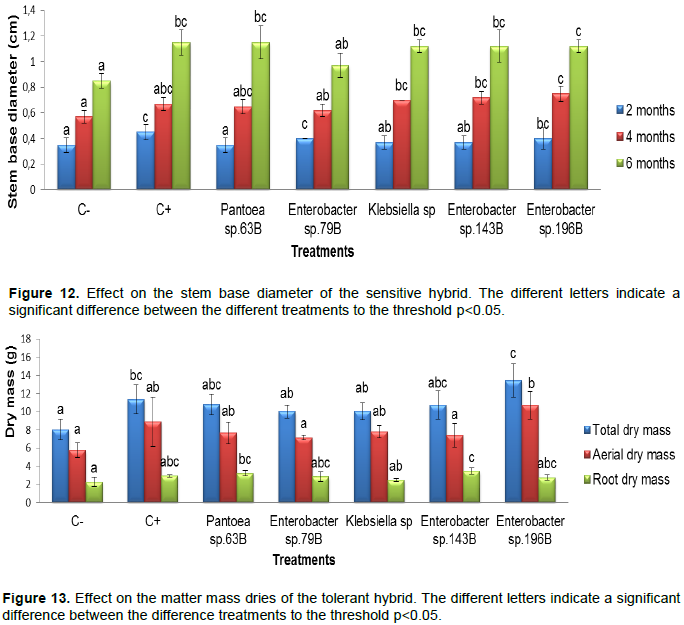
Effect over the plant high of the sensitive hybrid
At two months, the plant high of the plants inoculated with Pantoea sp. 63B (24 cm) and Klebsiella sp. 113B (24 cm) presented the highest values. At four months of inoculation, the result obtained showed that the control (+) presented the biggest plant high which was 34.5 cm followed by Pantoea sp. 63B (34 cm). At 6 months of inoculation, Pantoea sp. 63B presented the greatest value of the plant high which was 51.97 cm (Figure 8).
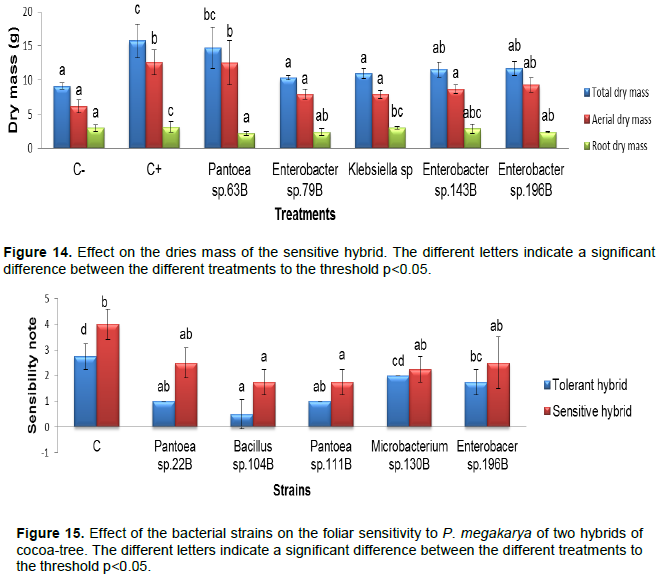
Effect of the number of leaves of the tolerant hybrid
At two months of inoculation, the number of leave varied between 10 (control+) and 8.5 (control-). Among the inoculated treatments, Pantoea sp. 63B and Klebsiella sp. 113B, also presented 10 leaves. At four months of inoculation, the results indicated that the control (+) and Klebsiella sp. 113B had 14.5 leaves while at six months more, a large number of leave was observed in Enterobacter sp. 196B (20.25) (Figure 9).
Effect on the number of leaves of the sensitive hybrid
The results ontained showed a variation amongst leaves between the different treatments during month (Figure 10). Thus, at two months, Enterobacter sp. 196B presented more a large number of leaves which was 9.75 while the largest number of leave in the four months was observed at the treatments control (+) of Klebsiella sp. 113B and Enterobacter sp. 143B (15.25). With regard to the six months the positive control (+) had the largest number of leaves with an average of 21.5 follow-ups of Enterobacter sp. 196B (20.75).
Effect on the stem base diameter of the tolerant hybrid
At two months of inoculation (Figure 11), the stem base diameter of the plants inoculated with Pantoea sp. 63B (0.52 cm) and Enterobacter sp. 143B (0.52 cm) presented high values of the stem base diameter. At four months of inoculation, Pantoea sp. 63B (0.77 cm) and the control (+) (0.77 cm) were significant different from the control (-)(0.65 cm). At six months of inoculation, the got results showed a significant difference between the microbial treatments and the control (-).
Effect on the stem base diameter of the sensitive hybrid
The stem base diameter varied according to the various treatments (Figure 12). Thus, after 2 months of inoculation the control (+) and Enterobacter sp. 79B had the largest stem base diameter (0.45 cm). The largest stem base diameter in the four months was also observed at Enterobacter sp.196B (0.75 cm). At six months, the great
values of the stem base diameter was observed at the control (+) and Pantoea sp. 63B (1.15 cm).
Effect on the dry matter mass of the tolerant hybrid
The results obtained (Figure 13) indicated a significant difference as regards the variation of the total dry mass, the shoot dry mass and root dry mass six months after sowing. For the variation of the total dry mass, the largest and the smallest mass were observed respectively in Enterobater sp. 196 (13.44 g) and control (-) (8.03 g). Enterobacer sp.196B also had the greatest shoot dry mass (10.69 g) while the greatest root mass was observed in Enterobacter sp. 143B (3.46 g).
Effect on the dry mass of the sensitive hybrid
For the total dry mass (Figure 14), the control (+) (15.74 g) presented the greatest value followed by Pantoea sp. 63B (14.71 g). Just as regards the shoot dry mass, the control (+) presented the greatest value (12.59 g) followed by Pantoea sp. 63B. For the root dry mass, the greatest (3.14 g) was in the control (+) followed by Klebsiella sp. 113B (3.04 g).
Incidence of the inoculation of P. megakarya on the leaves of cocoa-tree from presence and absence of PSB
The action of five bacterial strains on P. megakarya was evaluated by the test on leaves discs. The results showed that for the tolerant hybrid, the notes of foliar sensitivity varied from 0.5 to 2 while in the sensitive hybrid, the notes varied from 1.75 to 2.5.
In both cases, the most important effect was obtained with the strains Bacillus sp. 104B (0.5 for the tolerant hybrid and 1.75 for the sensitive hybrid) and Pantoea sp. 111B (1 for the tolerant hybrid and 1.75 for the sensitive hybrid). In addition, the statistical analyses showed that there was no significant difference between these two strains for the two hybrids (Figure 15).
PSMs are able to release phosphorus from the soil minerals play a key role in soil fertility. The same observations were made by Wakelin et al. (2012), Huanhuan et al. (2018) and Xiaohui et al. (2018).
Phosphate is a nutrient and its unavailability limits plant growth in many countries and regions. With the average solubilization index (5.5) of Pantoea sp.63B and its ability to solubilize all the different mineral phosphates (4.47 for phosphate of Algeria, 8.36 for phosphate of Cameroon, 8.76 for phosphate of Mali, 2.73 for Morocco and 3.1 for Mexico), Pantoea sp.63B can be considered as the most effective strain. This result approaches those obtained by Fankem et al. (2014b) during a study relating to the effect of bacteria solubilization of phosphates (Pantoea sp., Klebsiella sp. and Bacillus sp.) on the growth and the output of soybean. The capacity of solubilization of different types of phosphate showed that the rock phosphate of Cameroon (IS = 6.9) is solubilized more in solid medium, followed by rock phosphate of Mali (IS = 6.53), rock phosphate of Algeria (IS = 4.8), rock phosphate of Morocco (IS= 1.9) and finally by rock phosphate of Mexico (IS = 1.38). These values are contradictory with those obtained by Babana et al. (2013) and Fankem et al. (2014a) which obtained IS going from 1 to 3.6. However, they approach those obtained by Maliha et al. (2004) which obtained values of index of solubilization going from 2.16 to 6.23.
The contradiction of the results between the solid medium and the liquid medium was observed by many authors Deubel and Merbach (2005), Baig et al. (2010) and Fankem et al. (2014b). The absence of the zone of halation in the solid medium would be due to the absence or with the weak diffusion of the organic acids produced by the strain (Babana et al., 2013). It was thus judicious to combine the qualitative method with the quantitative method to lead the operations of selection of the PSB. Similar results were reported by Nautiyal (1999), Nautiyal et al. (2000) and Baig et al. (2010).
To select the strains of PSB in laboratory, the activities antagonistic with respect to P. megakarya as measured in vitro on mycelia growth. The results showed a clear reduction of the diameter of P. megakarya in presence of eight strains of PSB tested (Pseudomonas sp. 1B, Pantoea sp. 22B, Burkholderia sp. 36B, Bacillus sp. 104B, Pantoea sp. 111B, Microbacterium sp. 130, Pseudomonas sp. 179B and Enterobacter sp. 196B). The strongest inhibiting actions were observed with the strains Pantoea sp. 111B, Bacillus sp. 104B, Micobacterium sp. 130B, Enterobacter sp. 196B and Pseudomonas sp. 1B. Many authors studied the effect of fungi and bacteria on the mycelia growth of Phytophthora sp. on disc (Kébé et al., 2009; Mpika et al., 2009). According to Manjula et al. (2004), purified chitinases of Bacillus subtilis AF1 are highly antifungal. Kébé et al. (2009) showed that Bacillus sp. reduced the mycelial growth of Phytophthora sp. on Petri dishes. Similar results were found by Tondje et al. (2007) which announced a strong inhibiting action of Trichoderma asperellum on P. megakarya. Nwaga et al. (2007) reported that Pseudomonas sp. is agents of biological control against the fungi diseases. According to Weller et al. (2002; 2007), Pseudomonas produce many antifungal metabolites. Indeed, according to Haas and Défago (2005), most Pseudomonas produce antifungal such as phenazines, the pyoluteorine, the pyrrolnitrine and the DAPG (2, 4-diacetylphloroglucinol) which are the most antifungal frequently detected.
All the strains tested in greenhouse showed an aptitude to promote the growth of the plant in the presence of rock phosphate of Cameroon compared to the negative control. For the tolerant hybrids (UPA143 × T72/501), Enterobacter sp. 196B showed an increase of 50% height of the plant high, 35% amongst number of leaves, 38.88% of the stem base diameter and 67.37% of the dry weight total compared to the control (-). For sensitive hybrids (SNK64 × UPA143), Pantoea sp.63B showed an increase of 43.28% in stem height and an increase of 61.11% in total dry weight; Enterobacter sp.196B showed an increase of 22.05% in leaf number and an increase of 31.76% in stem base diameter compared to the control (-). These values are higher than those obtained by Tandon (1987) who showed that the inoculation with the PSM induces an increase in the growth from 10 to 15%, but approach those of Fankem et al. (2014b) who showed that, the inoculation of soybean with PSB induces an increase in the growth of approximately 35%. The results obtained showed that the strains solubilzed insoluble rock phosphate and make available of plants phosphorus necessary to their growth. Verma (1993) affirms that the use of the PSB can increase the output of the agricultural production to more than 70%.
The study of the action of the bacterial strains on P. megakarya by the test on foliar discs highlighted a reduction of the foliar sensitivity to P. megakarya of the tolerant (UPA143 × T72/501) or sensitive (SNK64 × UPA143) hybrids of cocoa tree. Many authors as Maurhofer et al. (1994) on the tobacco, Duijff et al. (1997) on tomato, Chen et al. (1998) on cucumber and Kébé et al. (2009) on the leaves of cocoa-tree reported similar results. The strongest effects were recorded with the bacterial strains Bacillus sp. 104B and Pantoea sp. 111B, which makes these two bacterial strains of the potential candidates to the biological fight against P. megakarya. According to Kébé et al. (2009), the Bacillus kind equipped with an aptitude to sporulate, which could predict fight. This study thus made it possible to highlight several potential antagonists of Phytophthora megakarya, likely to be used in the fight against the black pod disease of cocoa-tree.
At the end of this study, the results obtained on the characterization of the strains on petri dishes and in liquid medium showed the need for combining the two methods in order to select strains of qualified PSB. In addition, the results obtained revealed strains of PSB having an antagonistic effect proven against P. megakarya under in vitro and in vivo conditions on foliar discs. This effectiveness let’s consider the possibility of using these microorganisms in the fight the black pod disease in cocoa-tree and the promotion of the growth of this last. However, this hypothesis can only be considered if the efficacy observed under in vitro conditions is confirmed on cocoa tree in the field. This second stage of the study requires the massive knowledge of the production technic of the microorganisms; to carry out tests in fields in optics to confirm the results obtained in laboratory and greenhouse, finally to popularize the use of these microorganisms among farmers.
The authors have not declared any conflict of interests.
REFERENCES
|
Anonymous (2014). Monographic study on the cocoa sector in UEMOA. 33p.
|
|
|
|
Anushka MW, Dunwell JM (2018). Cacao biotechnology: current status and future prospects. Plant Biotechnology Journal 16(1):4-17.
Crossref
|
|
|
|
|
Arnold B (1999). Fungal endophytes of tropical trees: Methods and potential for biological control of fungal pathogens of cocoa. Research methodology in biocontrol of plantdiseases with special reference to fungal diseases of cocoa. Worshop manual, CATIE, Turrialba, Costa Rica. Edited by Ulrike Krauss and PrakashHebbar: pp. 44-54.
|
|
|
|
|
Babana AH, Dicko AH, Maïga K, Traoré D (2013). Characterization of rock phosphate-solubilizing microorganisms isolated from wheat (Triticum aestivum L.) rhizosphere in Mali. Journal of Mıcrobıology and Mıcrobıal Research 1(1):1-6.
|
|
|
|
|
Baig KS, Arshad M, Zahir ZA, Cheema MA (2010). Comparative efficacy of qualitative and quantitative methods for rock phosphate solubilization with phosphate solubilizing rhizobacteria. Soil and Environment 29(1):82-86.
|
|
|
|
|
Blaha G, Eskes AB, Kébé BI, Tahi GM, Nyassé S (2000). Early screening of resistance to Phytophthora spp. by means of disc inoculation. In: Eskes AB & Efron Y., Proceedings of the CFC/ICCO/IPGRI Project workshop on working procedures for cocoagermplasm evaluation and selection. 1-6 February 1998. Montpellier (France) pp. 103-107.
|
|
|
|
|
Chen C, Bélanger RR, Benhamou N, Paulitz TC (1998). Induced systemic resistance (ISR) by Pseudomonas spp. Impairs pre-and post-infection development of Pythium aphanidermatumon cucumber roots. The European Journal of Plant Pathology 104(9):877-886.
Crossref
|
|
|
|
|
Deubel A, Merbach W (2005). Influence of microorganisms on phosphorus bioavailability in soils. In: Microorganisms in Soils: Roles in Genesis and Functions. Soil Biology 3:177-191.
Crossref
|
|
|
|
|
Duijff BJ, Gianinazzi-Pearson V, Lemanceau P (1997). Involvement of the outer membrane lipopolysaccharides in the endophytic colonization of tomato roots by biocontrol Pseudomonas fluorescens strain WCS417r. New Phytologist 135:325-334.
Crossref
|
|
|
|
|
Edi-Premono, Moawad MA, Vleck PLG (1996). Effect of phosphate solubilizing Pesudomonas putida on the growth of maize and its survival in the rhizosphere. Journal of Crop Science11:13-23.
|
|
|
|
|
Etoa PA (2009). Round Table on the Sustainable World Coconut Economy in Cameroon 14 p.
|
|
|
|
|
Evans HC, Holmes KA, Thomas SE (2003). Endophytes and mycoparasites associated with an indigenous forest tree, Theobroma gileri, in Ecuador and preliminary assessment of their potential as biocontrol agents of cocoa diseases. Mycological Progress 2(2):149-160.
Crossref
|
|
|
|
|
Fankem H (2007). Occurrence and potentials of phosphate solubilizingmicroorganisms associated with oil palm (Elaeis guineensis Jacq.) rhizosphere in Cameroon. PhD Thesis, University of Yaoundé 135 p.
|
|
|
|
|
Fankem H, Ngo Nkot L, Deubel A, Quinn J, Merbach W, Etoa FX, Nwaga D (2008). Solubilization of inorganic phosphates and plant growth promotion by strains of Pseudomonas fluorescens isolated from acidic soils of Cameroon. African Journal of Microbiology Research 2:171-178.
|
|
|
|
|
Fankem H, Ngo Nkot L, Nguesseu NG, Tchuisseu TGV, Tchiazé IAV, Nwaga D (2014a). Rock phosphate solubilisation by strains of Penicillium spp. isolated from farm and forest soils of three agro ecological zones of Cameroon. American Journal of Agriculture and Forestry 2(2):25-32.
Crossref
|
|
|
|
|
Fankem H, Nguesseu NG, Ngo Nkot L, Tchuisseu TGV, Nwaga D, Etoa FX (2014b). Rock-phosphate solubilising bacteria and their effect on soybean (Glycine max) growth under pot grown conditions. American Journal of Agriculture and Forestry 2(4):100-109.
Crossref
|
|
|
|
|
Haas D, Défago G (2005). Biological control of soil-borne pathogens by fluorescentpseudomonas. Nature Reviews Microbiology 3:307-319.
Crossref
|
|
|
|
|
Huanhuan J, Peishi Q, Tong W, Mian W, Mingna C, Na C, Lijuan P, Xiaoyuan C (2018). Isolation and characterization of halotolerant phosphate-solubilizing microorganisms from saline soils. Biotech 8:461.
Crossref
|
|
|
|
|
Hugenin G, Boccas B (1971). Rôle de quelques facteurs dans la formation et la germination des oospores chez le phytophthora palmivora butl. Annales de Phytopathologie 3(3):353-371.
|
|
|
|
|
International CoCoa Organization (ICCO) (2013). The world cocoa market, (En ligne).
View (Page consultée le 2 août 2013).
|
|
|
|
|
Kébé IB, Mpika J, Kouamé FN, Prakash KH, Gary SS, Ake S (2009). Isolation and identification of indigenous microorganisms of cocoa trees in Côte d'Ivoire and demonstration of their antagonistic effects against Phytophthora palmivora, agent of black pod disease. The Science of Nature 6(1):71-82.
|
|
|
|
|
Maliha R, Khalil S, Ayub N, Alam S, Latif F (2004). Organic acids production and phosphate solubilization by phosphate solubilizing microorganisms (PSM) under in vitro conditions. Pakistan Journal of Biological Sciences 7:187-196.
Crossref
|
|
|
|
|
Manjula K, Kishore GK, Podile AR (2004). Whole cells of Bacillus subtilis AF 1 proved effective than cell free and chitinase-based formulations in biological control of citrus fruit rot and groundnut rust. Canadian Journal of Microbiology 50:737-744.
Crossref
|
|
|
|
|
Maurhofer M, Hase C, Meuwly P, Métraux JP, Défago G (1994). Induction of systemic resistance of tobacco to tobacco necrosis virus by the root colonizing Pseudomonas fluorescens strain CHAO: influence of the gacA gene and of pyoverdine production. Phytopathology 84:139-146.
Crossref
|
|
|
|
|
Mfegue CV (2012). Origins and mechanisms of dispersal of populations of Phytophthora megakarya, cocoa pathogen in Cameroon. PhD thesis in Integrative Plant Biology. International Center for Advanced Studies in Agricultural Sciences-Montpellier Supagro 186 p.
|
|
|
|
|
Morrissey J, Stack Conrad J, Valls R, Motamayor CJ (2019). Low-cost assembly of a cacao crop genome is able to resolve complex heterozygous bubbles. Horticulture Research 6(44):16-13.
Crossref
|
|
|
|
|
Mpika J, Kébé BI, Druzhinina IS, Komon- Zélazowka KM, Kubicek PC, Aké S (2009). Inhibition of Phytophthora palmivora, agent of black pod disease in Ivory Coast, by Trichoderma sp. Science of Nature 6(1):49-62.
Crossref
|
|
|
|
|
Nautiyal CS (1999). An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. Fems Microbiology Letters 170:265-270.
Crossref
|
|
|
|
|
Nautiyal CS, Bhadauria S, Kumar P, Lal H, Mondal R, Verma D (2000). Stress induced phosphate solubilization in bacteria isolated from alkaline soils. Fems Microbiology Letters 182:291-296.
Crossref
|
|
|
|
|
Ndoumbe-Nkeng M (2002). Impact of agro-ecological factors on the epidemiology of black pod disease in Cameroon: contribution to the establishment of an agricultural warning model. Paris, France: INA-PG, PhD thesis 189 p.
|
|
|
|
|
Ndoumbé-Nkeng M, Cilas C, Myuemb E, Nyassé S, Bieysse D, Flori A, Sache I (2004). Impact of removing diseased pods on cocoa black pods cause by Phytophthora megakarya and on cocaoa production in Cameroon. Crop Protection 23:415-424.
Crossref
|
|
|
|
|
Nwaga D, Fankem H, Essono OG, Ngo Nkot L, Randrianangaly JS (2007). Pseudomonads and symbiotic micro-organisms as biocontrol agents against fungal disease caused by Pythium aphanidermatum. African Journal of Biotechnology 6(3):190-197.
|
|
|
|
|
Nwaga D, Ngonkeu MEL, Oyong MM, Ngakou A, Abelong MP, Foko J (2000). Soil beneficial micro-organisms and sustainable agricultural production in Cameroon: current research and perspectives. In The biology and fertility of tropical soils, TSBF Report 1998, UNESCO-TSBF, Nairobi, Kenya pp. 62-65.
|
|
|
|
|
Nyadanu D, Akromah R, Adomako B, Kwoseh C, Dzahini-Obiatey H, Lowor ST, Akrofi AY, Assuah MK (2012). Host plant resistance to Phytophthora pod rot in cocoa (Theobroma cacao L.): The role of epicuticular wax on pod and leaf surfaces. International Journal of Botany 8(1):13-21.
Crossref
|
|
|
|
|
Nyassé S, Cilas C, Hérail C, Blaha G (1995). Leaf inoculation as an early screening test for cocoa (Theobroma cacao L.) resistance to Phytophthora black pod disease. Crop Protection 14(8):657-663.
Crossref
|
|
|
|
|
Pokou ND, Fister SA, Winters N, Tahi M, Klotioloma C, Sebastian A, Marden HJ, Maximova NS, Guiltinan JM (2019). Resistant and susceptible cacao genotypes exhibit defense gene polymorphism and unique early responses to Phytophthora megakarya inoculation. Plant Molecular Biology 99:499-516.
Crossref
|
|
|
|
|
Robin R (2012). Stimulate the growth of plants to better develop agricultural resources: Mineral fertilizers as hosts of microorganisms stimulating the growth of plants. ARTAS / AFCAS sugar congress, Reunion 7 p.
|
|
|
|
|
Rubini MR, Silva-Ribeiro RT, Pomella AWV, Maki CS, Araujo WL, dos Santos DR, Azevedo JL (2005). Diversity of endophytic fungal community of cacao (Theobroma cacao L.) and biological control of Crinipellis perniciosa, causal agent of Witches' Broom Disease. International Journal of Biological Sciences 1:24-33.
Crossref
|
|
|
|
|
Takam SP (2011). Estimation of the length of stay in the presence of a post-event censorship of interest. Application to the growth of cocoa fruits and modeling the risk of attack by black pod disease in Cameroon. PhD thesis in Applied Mathematics. University of Montpellier II (France) 190 p.
|
|
|
|
|
Tandon HL (1987). Phosphorus Research and Production in India. Fertilizer Development and Consultation Organisation New Delhi 160 p.
|
|
|
|
|
Tondje PR, Roberts DP, Bon MC, Widmer T, Samuels GJ, Ismaiel AD, Begoude AD, Tchana T, Nyemb-Tshomb E, Ndoumbe-Nkeng, Bateman R, Fontem D, Hebbar KP (2007). Isolation and identification of mycoparasitic isolates of Trichoderma asperellum with potential for suppression of black pod disease of cacao in Cameroon. Biological Control 43:202-212.
Crossref
|
|
|
|
|
Tyler JA, Valérie G, Hani A, Russell JT (2008). Multifaceted beneficial effects of rhizosphere microorganisms on plant health and productivity. Soil Biology and Biochemistry 40:1733-1740.
Crossref
|
|
|
|
|
Verma LN (1993). Biofertiliser in agriculture. In: PK Thampan (ed.) Organics in soil health and crop production. Peekay Tree Crops Development Foundation, Cochin, India pp. 152-183.
|
|
|
|
|
Wakelin S, Mander C, Gerard E, Jansa J, Erb A, Young S, Condron L, O'Callaghan M (2012). Response of soil microbial communities to contrasted histories of phosphorus fertilisation in pastures. Applied Soil Ecology 61(5):40-48.
Crossref
|
|
|
|
|
Wang SL, Hsaaiao WJ, Chakg WT (2002). Purification and characterization of an antimicrobial chitinaseextracellulary produces by Monoscuspurpureus CCR31499 in a Shirmp and crabs shell powder medium. Journal of Agricultural and Food Chemistry 50:2249-2255.
Crossref
|
|
|
|
|
Weller DM, Raaijmakers JM, McSpadden-Gardener BB, Thomashow LS (2002). Microbial populations responsible for specific soil suppressiveness to plant pathogens. Annual Review of Phytopathology 40:309-348.
Crossref
|
|
|
|
|
Weller DM, Landa BB, Mavrodi OV, Schroeder KL, de la Fuente L, Blouin-Bankhead SB, Allende-Molar R, Bonsall RF, Mavrodi DV, Thomashow LS (2007). Role of 2, 4-diacetylphloroglucinol-producing fluorescent Pseudomonas spp. in the defense of plant roots. Plant Biology 9:4-20.
Crossref
|
|
|
|
|
Xiaohui W, Changdong W, Junkang S, Zhaoyang L, Qian L, Chao J, Xin S, Yurong H, Changqian W, Rongbo S, Jiamiao Z, Jianfeng D, Xunli L (2018). Isolation and characterization of phosphofungi, and screening of their plant growth‑promoting activities. AMB Express 8:1-12.
Crossref
|
|