Full Length Research Paper

ABSTRACT
The need for a complementary short-term mutagenicity bioassay with robust endpoints to the Ames assay has become increasingly crucial to in order to avoid false negative results. The alternative short-term test (STT) used in conjunction with the Ames increases the validity and decreases the number of false positive outcomes. As a result, Caco-2 cells (Human intestinal epithelial cell model) and RAW264.7 cells (mouse microphage-like cell line) were treated for 24 h with graded doses of hydrogen peroxide (0, 5, 10, 20, and 40 µM) (oxidative stress-inducing mutagen). Single- and double-strand DNA damage was quantified using single-cell gel electrophoresis (Comet assay). The head intensity, tail intensity, tail migration, and tail moment of the damaged DNA were analysed using an epifluorescence microscope with a gated camera and installed comet IV image analysis software. In Caco-2 and RAW264.7 cells, a significant drop in head intensity and a corresponding dose-dependent increase in tail intensity, tail migration, and tail moment are seen when were quantified w compared to the solvent control. The single cell gel electrophoresis (Comet assay) is a very sensitive, robust, and statistically reliable method for determining DNA damage utilising many parameters. As such, the comet assay is advised as a complement to existing short-term bioassays for mutagenicity, such as the Ames assay.
Key words: Mutagenicity, hydrogen peroxide, RAW264.7 cells, Caco-2 cells, single cell gel electrophoresis (Comet assay)
INTRODUCTION
The demand for technologies capable of rapidly predicting chemical carcinogens at a lower cost in terms of animal life and money continues to be a research priority. Historically, the convergence of fundamental genetic research on chemically induced mutagenesis and the Millers' work on electrophilic, DNA reactive chemical carcinogens compelled the scientific community to prioritise mutation-based short-term tests (STTs) over alternative methodologies (Zeiger, 2004; Benigni and Bossa, 2011). Due to the fact that no single approach is capable of detecting all conceivable genotoxic events, a diverse array of test systems has been developed and is being utilised globally in regulatory schemes.
These include bacterial mutation tests for detecting gene mutations or chromosomal aberrations, bone-marrow cytogenetics assays (Mortelmans and Zeiger, 2000; Benigni and Cecilia, 2011) and micronucleus assays (Hayashi, 2016). However, weaknesses in current testing methodologies have been recognised, and as a result, regulatory agencies have modified their requirements worldwide (Benigni and Cecilia, 2011). Among these weaknesses are: a dearth of assays capable of detecting non-genotoxic carcinogens, an increased rate of false-positive results in in vitro mammalian cell STTs; and the extremely low sensitivity of in vivo mutagenicity STTs (Zeiger, 1998; Benigni and Bossa, 2011).
All these challenges have prompted the idea of developing novel assays.
The single-cell gel electrophoresis (Comet assay) is a robust in vitro methodology (Møller and Loft, 2014) that has the potential to model gene mutation and chromosomal aberration endpoints in mammalian cells and should complement the well-established Ames Salmonella assays in the screening for mutagenicity caused by oxidative stress and/or other chemical inducers (Ames et al., 1973, Mortelmans and Zeigler, 2000; Brendler-Schwaab et al., 2005). Single cell electrophoresis (Comet assay) has been suggested as the most popular method in genetic toxicology (Brendler-Schwaab et al., 2005) and is employed in the evaluation of oxidative DNA damage in HepG2 cell lines (Benhusein et al., 2010).
HeLa, TK6 and V79 cell lines (Speit et al., 2016) and the prediction of bladder cancer in ecogenotoxicological studies and mutagenesis (Rojas et al., 1999; Moneef et al., 2003).
The DNA-damage on Caco-2 (adenocarcinoma cells) and RAW264.7 (macrophage) cell lines induced by hydrogen peroxide was characterised in this study (a modelled oxidative stress chemical inducer) and the comet assay presented to complement the existing short-term bioassays.
Hydrogen peroxide is a well-known oxidative stress inducer (Benhusein et al., 2010; Petersen et al., 2000). The RAW264.7 cells are appropriate model macrophages produced from Abelson leukaemia virus transformed cell line derived from BALB/c mice (Hartley et al., 2008). They are capable of pinocytosis and phagocytosis and can kill target cells by antibody dependent cytotoxicity, hence, an important model for immune studies (Fuentes et al., 2014).
The Caco-2 cells are immortalised cell line of human colorectal adenocarcinoma cells with the ability to differentiate into an heterogenous mixture of gastro-epithelial cells under culture condition. They are important intestinal models for drug bioavailability and absorption assessment (Sambuy et al., 2005; Angelis and Turco, 2011).
MATERIALS AND METHODS
Chemicals and laboratory consumables
Dulbecco Modified Eagle Medium (DMEM, high glucose D5796), Penicillin, Streptomycin and Glutamine (PSG 100x), Trypsin (0.25%), Fetal bovine serum (FBS), Sodium pyruvate, Gelred, Hydrogen peroxide (H2O2), Phosphate buffer saline (PBS), Comet lysis buffer (2.5 M sodium chloride, 100 mM Na2EDTA, 10 mM Tris, pH 10, 1% sodium sarcosinate, 1% Triton X-100), electrophoresis solution (1 mM Na2EDTA, 300 mM NaOH, pH 13), and Tris buffer (0.4 M Tris, pH 7.5). All chemicals were purchased from Sigma chemicals Co. UK.
Cell culture and treatments
Caco-2 cells, a human colorectal adenocarcinoma cell line, were maintained in DMEM media containing 20% fetal bovine serum albumin (ATCC), penicillin (100 U/ml) and streptomycin (100 µg/ml) at 37°C in a 5% CO2 in air atmosphere. Similar recipes were used for RAW264.7 cell medium except with the addition of sodium pyruvate. 2 ml of Caco-2 cells (2.0 x 105 cell/well) were plated in a 24-well plate and incubated for 24 h at 37°C under humidified condition for the adherence of the cells. Afterwards, the medium was replaced with 2 ml aliquot of DMEM medium containing 0, 5, 10, 20 and 40 µM hydrogen peroxide and allowed for 24 h. Similar treatment was used for RAW264.7 cells. The comet assay was performed under alkaline conditions essentially following the procedure of Singh et al. (1988).
Comet slide preparation, electrophoresis, staining and analysis
An aliquot of 50 µl of the Caco-2 cell suspension from each well was mixed with 450 µl of 0.5%(v/v) low melting point agarose dissolved in PBS and held at 37°C. From this mixture, 50 µl aliquot was taken and placed onto a pre-treated and pre-warmed 20-well Trevigen microscope slide (Trevigen, #4250-050-03, Gaithersburg, MD). This was repeated for each treatment. The slide was incubated at 4°C for 15 min. The slide was submerged in a freshly prepared lysis solution (2.5 M NaCl, 100 mM Na2EDTA, 10 mM Tris, pH 10, 1% sodium sarcosinate, 1% Triton X-100) and incubated at 4°C for 4 h. Afterwards, the slides were transferred into a horizontal gel electrophoresis tank filled with freshly prepared electrophoresis solution (1 mM Na2EDTA, 300 mM NaOH, pH 13) maintained at 4°C for 30 min followed by voltage application for 30 min (0.74 V/cm, 300 mA). After the electrophoresis, the slides were rinsed with Tris buffer (0.4 M Tris, pH 7.5) for 10 min and rinsed with distilled water for 5 min. The slides were then transferred into ethanol solution (80%) for 5 min to remove excess water. The slides are further transferred into an incubator set at 37°C for drying. Before the slide image analysis, the slides were stained with Gelred [Sigma-Aldrich, #9Q05FE, (10000x)] for 30 min, rinsed and dried in an incubator set at 37°C. Slides were examined at x200 magnification using an epifluorescence microscope (LEICA, DMLB) equipped with excitation filter of 515-560 nm, connected through a gated CCD camera to installed Comet IV image analysis software (Instem, Stone, UK). Images of 100 cells per treatments were analysed and head intensity (%), tail intensity (%), tail moment and tail migration (expressed in arbitraty units) generated autonomously. The advantage of tail moment as an index of DNA damage is that both the amount of damage DNA and the distance of migration of the genetic material in the tail are represented by a single number. Data are presented as mean ± SE. The One-way ANOVA test was used to compare the means of each treatment using GraphPad Prism statistical software.
RESULTS
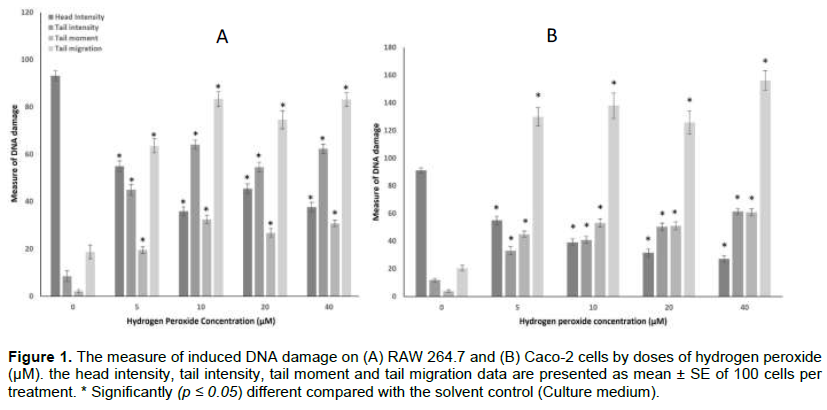
The extent of DNA damage on the RAW 264.7 (A) and Caco-2 cells (B) is presented in Figure 1. The degree of DNA damage in both cell lines is represented by Comet parameters such as head intensity (%), tail intensity (%), tail moment, and tail migration.
The RAW264.7 untreated control cells showed a background value for mean head intensity (93.18 ± 2.12%), tail intensity (8.54 ± 2.29%), tail moment (2.13 ± 0.70) and tail migration (18.75 ± 2.93). There was a significant (p < 0.05) increase in the tail intensity, tail moment, and tail migration of the hydrogen peroxide treatments compared with the control in both cell lines. A significant (p < 0.05) dose-dependent decrease was observed on the head intensity at 5 and 10 µM which normalised at 20 and 40 µM. Similarly, there was a significant (p < 0.05) and corresponding dose-dependent increase in the tail intensity, tail moment, and tail migration at 5 and 10 µM. The tail intensity and tail moment at 20 and 40 µM were not significantly (p >0.05) different. However, the tail migration showed a significant (p <0.05) increase at 40 µM over the 20 µM treatment.
Similarly, the Caco-2 cells showed a background mean value for the head intensity (91.18 ± 1.81%), tail intensity (11.95 ± 1.27%), tail moment (3.92 ± 0.90) and tail migration (20.47 ± 2.02). A significant (p < 0.05) reduction in the head intensity was observed at 5 and 10 µM which was normalised at 20 and 40 µM. In the same way, a corresponding significant (p < 0.05) increase in the tail intensity, tail moment, and tail migration was observed at 5 and 10 µM. However, the difference in the tail intensity, tail moment and tail migration at 20 and 40 µM was not statistically different.
DISCUSSION
The Ames test is the most widely used short-term genotoxicity assay with robust genetic endpoints. The Ames assay exhibits significant association with carcinogenicity and 80-84% interlaboratory reproducibility (Zeigler, 1987; Mortelsmans and Zeigler, 2000; Zeigler, 2010). Despite its usefulness, the Ames assay has drawbacks, such as its inability to detect non-genotoxic carcinogens.
The lack of a short-term no-mutation test to complement the Ames assay forced the search for an alternate bioassay that encompassed DNA damage. This study presents the Comet assay as a suitable complement of the Ames assay in the effort to validate the screening for mutagenicity using important mammalian cell model (Caco-2 cells and RAW 264.7, a model for gastrointestinal cells and immunity respectively) and hydrogen peroxide (an oxidative stress inducing chemical mutagen) (Figure 1).
Hydrogen peroxide produces oxidative stress by rapidly entering the cytoplasm and damaging DNA by producing hydroxyl-free radicals (Henzler and Steudle, 2000; Wang et al., 2005; Jaruga and Dizdaroglu, 1996). Toxic free radicals damage the DNA sugar residue, causing single- and double-strand breaks (Cadet et al., 1999; Phaniendra et al., 2015).
Also, they can convert purines and pyrimidines to their hydroxyl derivatives (Wang et al., 2005). Hydrogen peroxide may also cause C:G to T:A and C:G to G:C transversions in E. coli supF gene (Akasaka and Yamamoto, 1994). These genetic mutations are expressed by the Comet's head, tail, tail moment, and tail movement. To complement the comet experiment, all of these characteristics (Figure 1) depict oxidative stress-induced cell damage in Caco-2 and RAW264.7 cell lines. This study corroborates the findings of Collins (1999) and Kleiman et al. (1990) on oxidative damages of hydrogen peroxide of mammalian cells.
The tail moment is the most reported derived parameter (Ahnstrom, 1988; Piperakis et al., 2009; Azqueta et al., 2011).
A healthy cell has a head intensity of 100%. However, when DNA damage increases, the head intensity drops, resulting in an increase in tail intensity, tail migration, and tail moment (Figure 1). Because both cell lines are adherent, the background value in tail intensity, tail migration, and tail moment for solvent control is likely due to cell handling. After 24 h of treatment with hydrogen peroxide, both RAW264.7 and Caco-2 cells demonstrated dose-dependent DNA damage (Figure 1) except for 20 and 40 µM treatments. DNA repair mechanism likely interfered with the expressed DNA-damaged in RAW264.7 (Davies, 2000; Gasiorowski and Brokos, 2001; Rosignoli et al., 2001). However, the influence of the DNA repair mechanism seems to be reduced in Caco-2 cells (Figure 1). When cells are damaged by DNA, they activate a variety of response pathways. These processes include base excision repair (BER), nucleotide excision repair (NER), mismatch repair (MMR), homologous recombination (HR), and non-homologous end-joining (NHEJ) (Frenzilli et al., 2000; Clancy, 2008; Chatterjee and Walker, 2017).
The Comet bioassay employing RAW264.7 and Caco-2 cells is a robust and strong short-term experiment with good statistical applicability and can be considered as complement for the Ames assay in evaluating the genotoxicity of mutagens and carcinogens.
CONFLICT OF INTERESTS
The author has not declared any conflict of interests.
REFERENCES
|
Ahnstrom G (1988). Techniques to measure DNA single-strand breaks in cells - A review. International Journal of Radiation Biology 54(5):695-707. |
|
|
Akasaka S, Yamamoto K (1994). Hydrogen peroxide induces G:C to TA and G:C to C:G transversions in the supF gene of Escherichia coli. Molecular and General Genetics 243(5):500-505. |
|
|
Ames BN, Durston WE, Yamasaki E, Lee FD (1973). Carcinogens are mutagens: a simple test system combining liver homogenates for activation and bacteria for detection. Proceedings of the National Academy of Sciences (PNAS) of the United States of America 70(8):2281-2285. |
|
|
Angelis TD, Turco L (2011). Caco-2 cells as a model for intestinal Absorption. Current protocol in Toxicology, Chapter 20(1), Unit 20.6. |
|
|
Azqueta A, Gutzkow KB, Brunborg G, Collins AR (2011). Towards a more reliable comet assay: Optimising agarose concentration, unwinding time and electrophoresis conditions. Mutation Research-Genetic Toxicology and Environmental Mutagenesis 724(1-2):41-45. |
|
|
Benhusein GM, Mutch E, Aburawi S, Williams FM (2010). Genotoxic effect of induced by hydrogen peroxide in human hepatoma cells using the comet assay. Libyan Journal of Medicine 5(1):4637. |
|
|
Benigni R, Bossa C (2011). Alternate strategies for carcinogenicity assessment: an efficient and simplified approach based on in vitro mutagenicity and cell transformation assays. Mutagenesis 26(3):455-460. |
|
|
Brendler-Schwaab S, Hartmann A, Pfuhler S, Speit G (2005). The in vivo comet assay: use and status in genotoxicity testing. Mutagenesis 20 (4):245254 |
|
|
Cadet JD, Douki T, Gasparutto T, Pouget D, Ravanat JP, Sauvaigo JL (1999). Hydroxyl radicals and DNA base damage. Mutation Research/Fundamental and Molecular Mechanisms of Mutagenesis 424(1-2):9-21. |
|
|
Chatterjee N, Walker GC (2017). Mechanisms of DNA damage, repair, and mutagenesis. Environmental and Molecular Mutagenesis 58(5):235-263 |
|
|
Clancy S (2008). DNA damage and repair: Mechanisms for maintaining DNA integrity. Nature Education 1(1):103. |
|
|
Collins A (1999). Oxidative DNA damage, antioxidants, and cancer. Bioassays 21:238-46. |
|
|
Davies K (2000). Oxidative stress, antioxidant defences and damage removal, repair and replacement systems. IUBMB Life 50:279-89. |
|
|
Frenzilli G, Bosco B, Barale E (2000). Validation of single cell gel assay in human leukocytes with 18 reference compounds. Mutat Res. 468: 93-108. |
|
|
Fuentes AL, Millis L, Vapenik J, Sigola L (2014). Lipopolysaccharide-mediated enhancement of zymosan phagocytosis by RAW 264.7 macrophages is independent of opsonins, laminarin, mannan, and complement receptor 3. Journal of Surgical Research 189(2):304-312. |
|
|
Gasiorowski K, Brokos B. (2001). DNA repair of hydrogen peroxideinduced damage in human lymphocytes in the presence of four antimutagens. A study with alkaline single cell gel electrophoresis (Comet assay). Cellular and Molecular Biology Letters 6(2):141-159. |
|
|
Hartley JW, Evans LN, Green KY, Naghashfar Z, Macias AR, Zerfas, PM, Ward JM (2008). Expression of infectious murine leukemia virus by RAW264.7 cells, a potential complication for studies with a widely used mouse macrophage cell line. Retrovirology 5:1. |
|
|
Hayashi M (2016). The micronucleus test-most widely used in vivo genotoxicity test. Genes and Environment 38(1):1-6. |
|
|
Henzler T, Steudle E (2000). Transport and metabolic degradation of hydrogen peroxide in Chara corallina: model calculations and measurements with the pressure probe suggest transport of H2O2 across water channels. Journal of Experimental Botany 51(353):2053-2066. |
|
|
Jaruga P, Dizdaroglu M (1996). Repair of products of oxidative DNA base damage in human cells. Nucleic Acids Research 24(8):1389-1394. |
|
|
Kleiman NJ, Wang RR, Spector A (1990). Hydrogen peroxide-induced DNA damage in bovine lens epithelial cells. Mutat Res. 240: 35-45. |
|
|
Møller P, Loft S (2014). Statistical analysis of comet assay results. Frontiers in genetics, 5(292). |
|
|
Moneef MA, Sherwood BT, Bowman KJ, Kockelbergh RC, Symonds RP, Steward WP, Mellon JK, Jones GD (2003). Measurements using the alkaline comet assay predict bladder cancer cell radiosensitivity. British Journal of Cancer 89(12):2271-2276. |
|
|
Mortelmans K, Zeiger E (2000). The Ames Salmonella/microsome mutagenicity assay. Mutation Research 455(1-2)29-60. |
|
|
Petersen AB, Gniadecki R, Vicanova J, Thorn T, Wulf HC (2000). Hydrogen peroxide is responsible for UVA-induced DNA damage measured by alkaline comet assay in HaCaT keratinocytes. Journal of Photochemistry and Photobiology B: Biology 59(1-3):123-131. |
|
|
Phaniendra A, Jestadi DB, Periyasamy L (2015). Free radicals: properties, sources, targets, and their implication in various diseases. Indian Journal of Clinical Biochemistry 30(1):11-26. |
|
|
Piperakis M, Karanastasi G, Lakovidou-Kritsi Z, Piperakis M (2009). The use of comet assay in measuring DNA damage and repair efficiency in child, adult, and old age populations. Cell Biology and Toxicology 25(1):65-71. |
|
|
Rojas E, Lopez M, Valverde M (1999). Single cell gel cell electrophoresis assay: methodology and applications Journal of Chromatography B: Biomedical Sciences and Applications 722(1-2):225-254. |
|
|
Rosignoli P, Fabiani R, De Bartolomeo A, Spinozzi F, Agea E, Pelli MA (2001). Protective activity of butyrate on hydrogen peroxide-induced DNA damage in isolated human colonocytes and HT29 tumor cells. Carcinogenesis 22:1675-80. |
|
|
Sambuy Y, De Angelis I, Ranaldi G, Scarino ML, Stammati A, Zucco F (2005). The Caco-2 cell line as a model of the intestinal barrier: influence of cell and culture-related factors on Caco-2 cell functional characteristics. Cell Biology and Toxicology 21(1):1-26. |
|
|
Speit G, Schütz P, Bausinger J (2016). Different sensitivities of cultured mammalian cells towards aphidicolin-enhanced DNA effects in the comet assay. Mutation research. Genetic Toxicology and Environmental Mutagenesis, 803-804, 22-26. |
|
|
Wang K, Hong YJ, Huang ZQ (2005). Protective effects of silybin on human umbilical vein endothelial cell injury induced by H2O2 in vitro. Vascular Pharmacology 43:198-206. |
|
|
Zeiger E (1987). Carcinogenicity of mutagens: predictive capability of the Salmonella mutagenesis assay for rodent carcinogenicity. Cancer Research 47:1287-1296 |
|
|
Zeiger E (1998). Identification of rodent carcinogens and noncarcinogens using genetic toxicity tests: premises, promises, and performance. Regulatory Toxicology and Pharmacology 28:85-95. |
|
|
Zeiger E. (2004). History and rationale of genetic toxicity testing: an impersonal, and sometimes personal, view. Environmental Health Perspectives 44:363-371 |
|
|
Zeiger E. (2010). Historical perspective on the development of the genetic toxicity test battery in the United States. Environmental and Molecular Mutagenesis 51(8?9):781-791. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


