Full Length Research Paper

ABSTRACT
Strychnos camptoneura (Loganiaceae) is used in Congolese traditional medicine to treat various diseases such as malaria and diabetes, but not much is known about its effects on the immune system. Therefore, the purpose of this study was to investigate the immunomodulatory properties of S. camptoneura leaves. Mice were treated with the aqueous extract of S. camptoneura leaves (100 and 200 mg/kg); those administered with reverse osmosis water and β-1,3-glucan (150 mg/kg) were used as negative and positive controls, respectively. The immunomodulatory effects were measured 24 h post-treatment by quantifying the production of cytokines, and the number of myeloid and lymphoid cells. Treated mice with the aqueous extract showed a significant production of IL-4 and IL-10 compared to negative animals (P <0.001). Similar to β-1,3-glucan, both doses of aqueous extract markedly increased the number of macrophages, dendritic cells, CD4+ T and NK cells in treated-mice, but not of CD8+T cells. These findings suggest that the S. camptoneura leaves possess immunomodulatory properties. Further studies are needed to determine phytochemicals in these leaves and signaling pathways involved in the immunomodulation. This would help to better understand and valorize the therapeutic potential of S. camptoneura.
Key words: Strychnos camptoneura, immunomodulatory effects, cytokines, myeloid cells, lymphoid cells, mice.
INTRODUCTION
The immune system is a network of cells, chemicals (such as cytokines) and processes that function to protect host from foreign antigens such as microbes (such as bacteria, fungi and parasites), viruses, cancer cells and toxins (Marshall et al., 2018). The immune cells include myeloid lineage cells such as macrophages, dendritic cells, and neutrophils, which are involved in innate immunity. The immune system is also composed of lymphoid lineage cells that are B lymphocytes, CD4+ T and CD8+ T lymphocytes, and natural-killer (NK) cells.
The B lymphocytes produce antibodies and mediate the adaptative humoral immunity, while T lymphocytes support the adaptative cell-mediated immunity. After being activated, The CD4+T cells carry out multiple functions, ranging from activation of the cells of innate immune system, B lymphocytes, cytotoxic T cells, and they also play a critical role in the suppression of immune reaction (Luckheeram et al., 2012). CD8+ T and NK cells play a central role in the immunity to intracellular pathogens and cancer due to their ability to directly kill infected or malignant cells (Abel et al., 2018; Raskov et al., 2021).
Cytokines are the glycoproteins produced by a broad range of the immune cells and that act as messengers enabling these cells to communicate with one another and to generate a coordinated and robust immune response (O’Shea et al., 2019). Cytokines are classically subdivided into pro-inflammatory and anti-inflammatory cytokines. Pro-inflammatory cytokines, such as tumor necrosis factor-alpha (TNF-α), interleukin (IL)-1, IL-6 and interferon-gamma (INF-γ), generally promote the activation and proliferation of immune cells, and their homing to the sites of infection. Anti-inflammatory cytokines, such as IL-4 and IL-10, display the ant-inflammatory activities.
It is well known that the dysfunction of the immune system can cause various pathologies such as infectious diseases, parasitic diseases, inflammatory diseases, autoimmune diseases and cancer. Therefore, the search for immunomodulators to adjust immune responses to cope with diseases related to the immune system dysfunction has been gaining more and more interest for some time. The immunomodulators refer to any substance, natural or synthetic, that can suppress or stimulate immune responses (Behl et al., 2021). Immuno-suppressants are useful to treat the pathologies requiring immunosuppression, including autoimmune diseases and inflammatory disorders. By contrast, immunostimulants help to improve the immune functions of patients with immunodeficiency disorders, chronic infectious diseases and cancer.
Natural plant-based immunomodulators have gained increasing interest in recent decades as they have the potential to counteract the side effects and high cost of synthetic immunomodulators (Jantan et al., 2015). In addition, medicinal plants have long been recognized for their therapeutic properties, and up to 80% of people worldwide depend on them for some aspects of their primary health care (Zhang et al., 2015)A number of plants used in traditional medicine have proven immunomodulatory activities. These include Echinacea purpurea, Artemisia annua, Curcuma domestica, Aloe vera, Panx ginseng and Barleria prionitis, to name but a few (Alhazmi et al., 2021). Strychnos camptoneura (Loganiacea), named "Yindza" in the vernacular in the Republic of Congo, is a plant used by the tradi-practitioners to treat various pathologies, including malaria, parasitosis, diabetes, fever and microbial hernia, while little is known about its effects on the immune system. A previous study found that S. camptoneura possesses phytochemicals such as flavonoids and terpenoids, and antioxidant properties (Morabandza et al., 2016a). The purpose of this study was to investigate the immunomodulatory properties of this medicinal plant. Immunomodulatory effects of the aqueous extract of leaves on the production of cytokines (IL-4 and IL-10) and proliferation of immune cells were evaluated.
MATERIALS AND METHODS
Plant material and preparation of the aqueous extract
The plant material consisted of the leaves of S. camptoneura collected on March 2022 (drought period) from the forest of M'voula’s village, in Department of Cuvette-Ouest of the Republic of Congo. This village is located about 740 km from Brazzaville. The botanical authentication of plant material was made at the National Institute for Research in Exact and Natural Sciences (IRSEN) in comparison with the reference specimen registered under No. 271. The harvested leaves were cleaned with distilled water and then dried at room temperature for 14 days. Subsequently, the leaves were grounded into powder; 50 g of the powder were suspended in 500 ml of distilled water and thereafter macerated under agitation for 72 h. The macerated was filtered, and the filtrate was concentrated using rotavapor Buchi Switzerland apparatus at 60°C until excess of solvent was evaporated. Approximately 10 g of the extract was harvested and, subsequently, resuspended in distilled or reverse osmosis water before being used in mice.
Animals and experimental design
Swice mice aged from 4 to 5 weeks and weighing 15 to 20 g were used in this study. Animal experiments were conducted, on the one hand, at the pet store of the Faculty of Science and Technology of the Marien Ngouabi University (Republic of Congo) for the toxicity tests, and on the other hand, at the pet store of the University of Franche-Comté (France) for cytokine and leukocyte analysis. Animals used at the University of Franche-Comté were purchased from Janvier LABS Company (Genest-Saint-Isle, France). All animal study procedures were conducted in accordance with institutional guidelines. Mice were acclimatized to the environment for a week before starting the treatments. Animals were fed standard with free access to water and maintained at a night-day lighting rate (12 h of lighting, 12 h of darkness). After being acclimatized, the mice were divided into experimental groups and treated as presented in Table 1. All animals were treated by oral gavage.
Acute toxicity assessment
This was done in accordance with OECD Guideline No. 423 for the testing of chemical solutions, which determines the lethal dose (LD50) of 50% of experimental animals and the therapeutic dose (Jonsson et al., 2013). Briefly, the mice were left fasted for 24 h and thereafter treated with distilled water (Group 1) or with of 2000 mg/kg the aqueous extract of S. camptoneura leaves (Group 2). Animals were placed back in the cages and observed macroscopically for 30 min and then every hour for four hours. These observations aimed to assess signs of toxicity such as lethality, mobility, alertness, ptosis, aggressiveness, pilo-erection, animal response to external stimulus, vocalization, sleep and tremor.
Cytokine quantification
Mice were treated with reverse osmosis water or with the aqueous extract of S. camptoneura leaves (Groups 4 and 5). Twenty-four hours (24 h) post-treatment, blood samples were collected in an EDTA tube and centrifuged at 10000 rpm for 5 min at 4°C. Plasma from each animal was transferred in an Eppendorf tube and kept at -20°C until cytokines analysis. Plasma was diluted 1:4 with phosphate buffered saline (1× PBS) and the concentrations of IL-4 and IL-10 cytokines were determined using the commercial sandwich Enzyme-Linked Immunosorbent Assay (ELISA) according to the manufacturer's instructions. The IL-4 and IL-10 ELISA kits were purchased from Elabscience (Houston, Texas, USA).
Myeloid and lymphoid cell quantification
Mice were treated with reverse osmosis water (Group 3), with the aqueous extract (Groups 4 and 5) or with β-1,3-glucan (Group 6). Twenty-four hours after the treatment, approximately 75 μl of blood sample was taken from each animal in an ETDA tube and kept at 4°C. The immune cells were identified and quantified by flow cytometry analysis. Antibodies used for the myeloid lineage cells were: anti-CD45 for leukocytes, anti-CD11b for macrophages/ granulocytes, anti-CD11c for dendritic cells and anti-F4/80 for macrophages.
The antibody mixt included: 1.5 μl anti-CD45, 3 μl anti-CD11b, 3 μl anti-CD11c, 3 μl anti-F4/80 and 139.5 μl PBS 1X (mixte 1). Antibodies for the lymphoid lineage cells were: Anti-CD3 for T cells, anti-CD4 for CD4+ T cells, anti-CD8 for CD8+ T cells, NK-APC-A for NK cells and BV421-A for leukocytes. The mixte included: 1.5 μl anti-CD45, 3 μl anti-CD3, 3 μl anti-CD4, 1.5 μl anti-CD8, 1.5 μl anti-NK and 139.5 μl PBS 1X (mixte 2). To perform the flow cytometric analysis, 25 μl of blood from each animal was mixed with 50 μl of mixte 1 or 2. The mixture was incubated in darkness for 15 min at room temperature, 350 μl of red blood cell lysing buffer was added and then incubated in darkness for 20 min. Cell counting was performed using the BD FACSCanto™ II Flow Cytometry Systems (BD Biosciences, France). All antibodies were purchased from Elabscience (Houston, Texas, USA).
Statistical analysis
Data were analyzed using GraphPad Prism version 9.4.1. The Kruskal-Wallis one-way ANOVA test was performed to compare the experimental groups with each other. The results were presented as mean ± standard deviation (SD). P value < 0.05 was considered significant.
RESULTS
Toxicity of the aqueous extract of S. camptoneura leaves
As shown in Table 2, oral administration of the aqueous extract of S. camptoneura leaves at a high dose of 2000 mg/kg did not cause any deaths in mice. In addition, no change in psychomotor behaviors was observed in the treated animals compared to control animals that received 10 ml/kg of distilled water.

Effect of the aqueous extract of S. camptoneura leaves on cytokine production
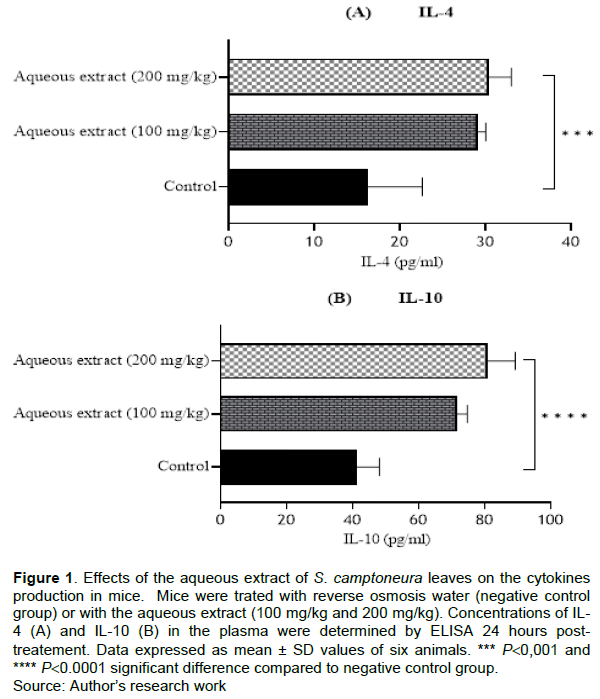
Effect of the aqueous extract of S. camptoneura leaves on IL-4 and IL-10 production was quantified by ELISA 24 h after the treatment. The concentrations of both cytokines in the plasma were significantly higher (p<0.001) in mice treated with the extract compared to control animals that were administered with reverse osmosis water. The mean concentrations of IL-4 in treated mice were 29.12 ± 0.89 and 30.38 ± 2.60 pg/ml at 100 and 200 mg/kg of the extract, respectively, whereas the mean contraction of this cytokine in negative control group was 16.20 ± 6.41 pg/ml (Figure 1A). For IL-10, the mean concentrations were 41.27± 6.86 pg/ml in control group and 71.56 ± 3.19 and 80.93 ± 8.37 pg/ml in treated mice with 100 and 200 mg/kg of the aqueous extract of S. camptoneura leaves, respectively (Figure 1B).
Effect of the aqueous extract of S. camptoneura leaves on myeloid and lymphoid cell proliferation
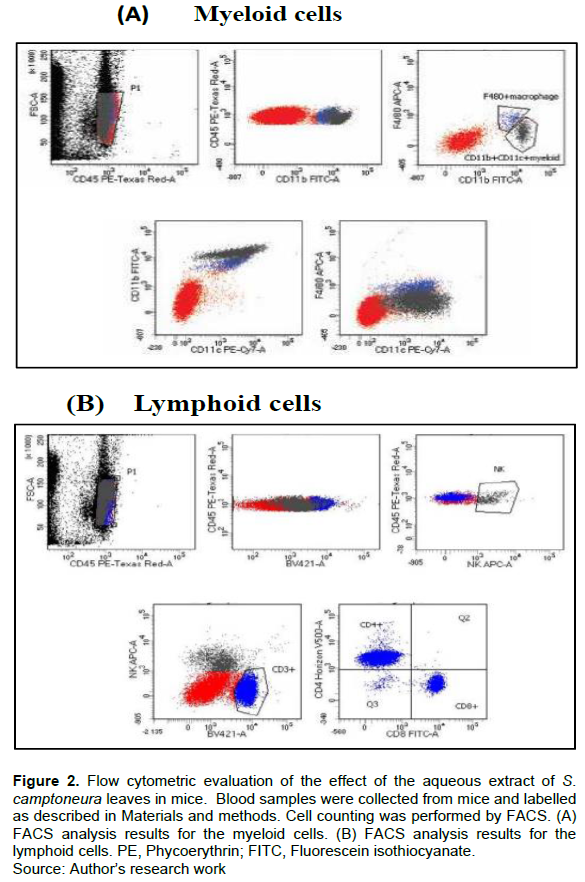
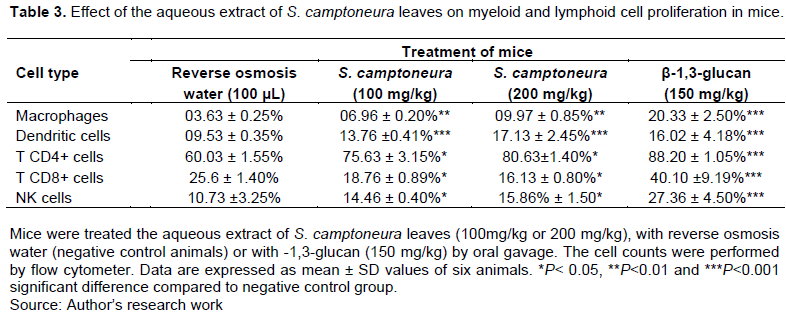
Analysis of the myeloid lineage cells (macrophages and dendritic cells) and lymphoid lineage cells (CD4+ T, CD8+T and NK cells) was carried out to investigate whether the aqueous extract of S. camptoneura leaves affects the proliferation of these cells. The results of flow cytometric analysis are reported in Figure 2 and Table 3. It was observed that the aqueous extract (at doses of 100 and 200 mg/kg) induced a significant increase (p<0.01) in the number of macrophages and dendritic cells in treated mice compared to negative control animals that were administered with reverse osmosis water. A significant increase in the number of CD4+ T and NK cells was also seen in animals treated with the aqueous extract. By contrast, a significant decrease in the number of CD8+ T cells was observed in treated-mice compared to negative control animals, and this decrease appeared to be proportional to the dose of treatment. On the other hand, the treatment of animals with β-1,3-glucan (150 mg/kg) strongly stimulated proliferation of macrophages, dendritic cells, CD4+ and CD8+T cells and NK cells compared to negative control animals (Table 3; p<0.001).
DISCUSSION
The aim of this present study was to investigate the immunomodulatory properties of S. camptoneura, a plant whose leaves, bark and stems are used in traditional Congolese medicine to treat a variety of pathologies. We evaluated effects of aqueous leaf extract of this plant on the production of anti-inflammatory cytokines, and on the proliferation of macrophages, dendritic cells, CD4+ and CD8+ T cells and NK cells in mice. We first examined the toxicity of this aqueous extract, and observed it did not cause any macroscopic signs of toxicity in the treated animals. Similar results had been obtained with the aqueous extract of S. camptoneura bark and stems (Morabandza et al., 2016b). Overall, and in accordance with the Harmonized Integrated System of Hazards to Human Health and the Environment of Chemical Substances and Mixtures (OECD, 2002), S. camptoneura can be classified as a non-toxic plant (OECD, 2002). Cytokine analysis showed that the aqueous extract of S. camptoneura leaves induced a very high production of IL-4 and IL-10 in treated mice. This indicated that this extract has anti-inflammatory properties. Indeed, IL-4 and IL-10, also known as Th2-type cytokines, are cytokines that in excess exert anti-inflammatory activities by inhibiting those of pro-inflammatory cytokines such as IL-1, TNF-α and IFN-γ (Maspi et al., 2016). IL-4 is a pleiotropic cytokine that influences Th cell differentiation as its early secretion leads to differentiation of naïve CD4 T cells (Th0) towards the Th2 phenotype. IL-4 can inhibit the production of Th1-type cytokines by reducing the production of IL-12 (Kaiko et al., 2008). IL-10 is considered the prototype of anti-inflammatory cytokines that contributes to the maintenance and reestablishment of immune homeostasis (Bedke et al., 2019). It downregulates the expression of Th1 cytokines by blocking the activity of the transcription factor NF-κB (Bedke et al., 2019). Previous studies also showed that some medicinal plants are able to induce the production of cytokines IL4 and IL-10. For example, Gholamnezhad et al. (2015)showed that Nigella sativa stimulated the production of IL4 in rats. The aqueous extracts of Combretum hereroense and Canthium mundianum leaves induced the production of IL-4 by the human peripheral blood mononuclear cells in vitro (Samie and Madzie, 2016). One the other hand, a study on the immunomodulatory effects of the Aloe vera peel extract in splenocyte cultures showed that this extract did not stimulate the production of IL-2 and INF-γ but did promote the production of IL-4 and IL-10 (Kwon et al., 2009). It was suggested that these effects could be due to the presence in Aloe vera of emodin which has an immunomodulatory effect by promoting Th2 cytokines and reducing Th1 cytokines (Liu et al., 2009). In the present study, we also observed that the aqueous extract of the S. camptoneura leaves stimulated production of IL-4 and IL-10. However, we were unable to identify phytochemicals responsible for the effects observed. We speculate that these effects may be due to the combined action of flavonoids and terpenoids, which were found within this plant (Morabandza et al., 2016b). Indeed, some flavonoids exert the anti-inflammatory activities by inhibiting transcription factors important for controlling mediators involved in inflammation (Maleki et al., 2019). On the other hand, studies have also shown that some terpenoids can inhibit the expression of NF-κB, a transcription factor that plays an important role in regulating immune and inflammatory responses, thereby promoting an anti-inflammatory response (Heras and Hortelano, 2009).
To examine in depth the immunomodulatory properties of S. camptoneura, we analyzed the effects of aqueous leaf extract on the production of myeloid and lymphoid cells in vivo. Myeloid cells include, among others, macrophages and dendritic cells. As the results showed, a significant increase in the number of macrophages and dendritic cells was observed in mice treated with the extract compared to negative control animals, which indicated its immunostimulatory effects on these cells as expected, a significant increase in the number of these cells was also observed in animals treated with β-1,3-glucan used in this study as positive control. Indeed, β glucans are well known as immunostimulants (Han et al., 2020). Since they are not produced by mammalian cells, β glucans are recognized as microbe-associated molecular patterns (MAMPs) by pattern recognition receptors (PRRs) expressed on the surface of innate immune (De Marco Castro et al., 2021). The binding of β-glucan to the receptor on dendritic cells and macrophages triggers their activation and maturation, increases their ability to present the antigen, and improves the production of pro-inflammatory cytokines that stimulate the polarization of Th1 and Th17 responses. It also induces the activation of CD8+ cytotoxic T lymphocytes (CTL) and NK cells (Bono et al., 2020). Exposed to external stimuli, macrophages can be differentiated into two subtypes, M1 and M2 macrophages (Saqib et al., 2018). M1 macrophages are activated mainly by IFN-γ, and they are responsible for inflammatory signaling and tissue damage. By contrast, M2 macrophages produce anti-inflammatory cytokines such as IL-4, IL-10 and TGF-β, which contribute to tissue healing and in the resolution of inflammatory process. Lipopolysaccharides (LPS) and IFN-γ drive the polarization of macrophages to the M1 phenotype, while IL-4 directs it to the M2 phenotype (Huang and Xu, 2019). It has been reported that phytochemicals such as polyphenols, flavonoids and terpenes have the ability to modulate the conversion of macrophages to the M2 phenotype (Saqib et al., 2018). Considering the fact that the aqueous extract of S. camptoneura leaves stimulates the production of IL-4 and contains flavonoids and terpenoids, we can conclude that this plant has the potential to promote the differentiation of M2 macrophages. Regarding dendritic cells, recent studies have shown that there are two conventional dendritic cell subpopulations, designated DC1 and DC2, whose differentiation strongly depends on the stimulus (Balan et al., 2019). Pro-inflammatory cytokines such as IFN-γ promote the generation of DC1, while anti-inflammatory cytokines (IL-4 and IL-10) drive the differentiation of DC2. Thus, the fact that the aqueous extract of S. camptoneura leaves induces the production of IL-4 and IL-10 suggests that these leaves contain phytochemicals that promote the differentiation of DC2. However, studies using the specific markers of dendritic cell subtypes are needed for verifying this assumption.
In this work, we also observed that the aqueous extract of S. camptoneura leaves stimulated the proliferation of CD4+ T and NK cells. The number of these cells was significantly increased in the blood of treated mice compared to negative control animals. CD4+ T lymphocytes, also known as helper T cells (Th), are divided into two major groups, designated Th1 and Th2 cells, mainly depending on the cytokines they produce (Zhu et al., 2010). Th1 cells produce IFN-γ as their signature cytokine and their proliferation evoke cell mediated immunity and inflammation. By contrast, Th2 cells fail to produce IFN-γ but do produce the anti-inflammatory cytokines such as IL-4, IL-5 and IL-13. From these facts, it can be assumed that the aqueous extract of S. camptoneura leaves promotes a Th2-type response in animals. Some previous studies have also been done regarding immunomodulatory effects of plants on NK cells. For example, it has been reported that the oral administration of white ginseng extracts enhances the cytotoxic activity and production of NK cells isolated from wild-type B6 mice but not from IFN-γ knockout mice, suggesting the involvement of IFN-γ in white ginseng’s immunostimulatory effect (Kim et al., 2022). NK cells are classically activated by Th1-type cytokines and, in turn, they secrete a large amount of IFN-γ that accelerates Th-type responses (Kiniwa et al., 2016). These authors also reported that overexpression of IL-4 induces the proliferation of tissue-resident macrophages and the production of IL-15, which contributes to the proliferation of a particular subpopulation of NK cells designated IL4-NK–like cells. This could explain the increase in the number of NK cells that we observed in our study.
Unlike treatment with β-1,3-glucan glucan, the treatment of mice with the aqueous extract of S. camptoneura leaves did not stimulate the proliferation of CD8+ T cells but did promote their decrease, which appeared to be dose-dependent. In addition, the amount of CD8+ T cells in treated mice was significantly lower compared to untreated animals. It is possible that at doses of 100 and 200 mg/kg, the aqueous extract of S. camptoneura leaves may cause TCD8+ cell death. It is also possible that this extract contains phytochemicals that have an immunosuppressive effect on CD8+ T cell proliferation.
One of the limitations of our study is that we were not able to analyze pro-inflammatory cytokines, including IFN-γ. We also did not examine the kinetics of cytokine and immune cell production. These analyses would also have made it possible to determine the level of production of pro-inflammatory cytokines and, consequently, to evaluate the immunomodulatory effects of the aqueous extract of S. camptoneura leaves over a long period of time.
CONCLUSION
This study demonstrates that the aqueous extract of S. camptoneura leaves have immune-modulatory effects on cytokines and leukocytes. The extract augments the production of IL-4 and IL-10, and the number of macrophages, dendritic cells, CD4+ T cells and NK cells. The immunologically active compounds present in leaves and their signaling pathways involved that immunomodulation need to be identified. This would help to valorize the therapeutic potential of S. camptoneura leaves, particularly in the treatment of inflammation-related diseases.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Abel AM, Yang C, Thakar MS, Malarkannan S (2018).Natural Killer Cells: Development, Maturation, and Clinical Utilization. Frontiers in Immunology 9:1869. |
|
|
Alhazmi HA, Najmi A, Javed SA, Sultana S, Al Bratty M, Makeen HA, Meraya AM, Ahsan W, Mohan S, Taha MME, Khalid A (2021). Medicinal Plants and Isolated Molecules Demonstrating Immunomodulation Activity as Potential Alternative Therapies for Viral Diseases Including COVID-19. Frontiers in Immunology 12:637553. |
|
|
Balan S, Saxena M, Bhardwaj N (2019). Dendritic cell subsets and locations. In International Review of Cell and Molecular Biology 348:1-68. |
|
|
Bedke T, Muscate F,Soukou S, Gagliani N, Huber S (2019). IL-10-producing T cells and their dual functions. Seminars in Immunology 44:101335. |
|
|
Behl T, Kumar K, Brisc C, Rus M,Nistor-Cseppento DC, Bustea C, Aron RAC, Pantis C, Zengin G, Sehgal A, Kaur R, Kumar A, Arora S, Setia D, Chandel D, Bungau S (2021).Exploring the multifocal role of phytochemicals as immunomodulators. Biomedicine and Pharmacotherapy 133:110959. |
|
|
Bono C, Martínez A, Megías J, Gozalbo D, Yáñez A, Gil M (2020). Dectin-1 Stimulation of Hematopoietic Stem and Progenitor Cells Occurs In Vivo and Promotes Differentiation Toward Trained Macrophages via an Indirect Cell-Autonomous Mechanism. MBio 11(3):e00781-20. |
|
|
De Marco Castro E, Calder PC, Roche HM (2021).β?1,3/1,6?Glucans and Immunity: State of the Art and Future Directions. Molecular Nutrition and Food Research 65(1):1901071. |
|
|
Gholamnezhad Z, Rafatpanah H, Sadeghnia HR, Boskabady MH (2015). Immunomodulatory and cytotoxic effects of Nigella sativa and thymoquinone on rat splenocytes. Food and Chemical Toxicology 86:72-80. |
|
|
Han B, Baruah K, Cox E, Vanrompay D, Bossier P (2020).Structure-Functional Activity Relationship of β-Glucans from the Perspective of Immunomodulation: A Mini-Review. Frontiers in Immunology 11:658. |
|
|
Heras B, Hortelano S (2009).Molecular Basis of the Anti-Inflammatory Effects of Terpenoids. Inflammation and Allergy-Drug Targets 8(1):28-39. |
|
|
Huang TL, Xu (2019).TET1 Knockdown Inhibits Porphyromonas gingivalis LPS/IFN-γ-Induced M1 Macrophage Polarization through the NF-κB Pathway in THP-1 Cells. International Journal of Molecular Sciences 20(8):2023. |
|
|
Jantan I, Ahmad W, Bukhari SNA (2015). Plant-derived immunomodulators: An insight on their preclinical evaluation and clinical trials. Frontiers in Plant Science 6:655. |
|
|
Jonsson M,Jestoi M,Nathanail AV,Kokkonen UM,Anttila M, Koivisto P, Karhunen P,Peltonen K(2013). Application of OECD Guideline 423 in assessing the acute oral toxicity of moniliformin. Food and Chemical Toxicology 53:27-32. |
|
|
Kaiko GE,Horvat JC,Beagley KW,Hansbro PM (2008).Immunological decision-making: How does the immune system decide to mount a helper T-cell response? Immunology 123(3):326-338. |
|
|
Kim JH, Kim DH, Jo S, Cho MJ, Cho YR, Lee YJ, Byun S (2022). Immunomodulatory functional foods and their molecular mechanisms. Experimental and Molecular Medicine 54(1):1-11. |
|
|
Kiniwa T, Enomoto Y, Terazawa N, Omi A, Miyata N, Ishiwata K, Miyajima A (2016).NK cells activated by Interleukin-4 in cooperation with Interleukin-15 exhibit distinctive characteristics. Proceedings of the National Academy of Sciences 113(36):10139-10144. |
|
|
Kwon KH, Hong MK, Hwang SY, Moon BY, Shin S, Baek JH, Park YH (2009). Antimicrobial and immunomodulatory effects of Aloe vera peel extract. Journal of Medicinal Plants Research 5:5384-5392. |
|
|
Liu YX., Shen NY, Liu C, Lv Y (2009). Immunosuppressive Effects of Emodin: An In Vivo and In Vitro Study. Transplantation Proceedings 41(5):1837-1839. |
|
|
Luckheeram RV, Zhou R, Verma AD, Xia B (2012). CD4 + T Cells: Differentiation and Functions. Clinical and Developmental Immunology 2012:1-12. |
|
|
Maleki SJ, Crespo JF, Cabanillas B (2019). Anti-inflammatory effects of flavonoids. Food Chemistry 299:125124. |
|
|
Marshall JS, Warrington R, Watson W, Kim HL (2018). An introduction to immunology and immunopathology. Allergy, Asthma and Clinical Immunology 14(S2):49. |
|
|
Maspi N, Abdoli A, Ghaffarifa F (2016). Pro- and anti-inflammatory cytokines in cutaneous leishmaniasis: A review. Pathogens and Global Health 110(6):247-260. |
|
|
Morabandza CJ, Amboyi GS, Matini L, Gouolali T, Ongoka PR, Abéna AA (2016a).Phytocheminal and antioxidant properties of bark and stems extract of S. camptoneura Gilg and Busse (Loganiaceae). Research Journal of Chemical Sciences 6(10):19-23. |
|
|
Morabandza CJ, Gombé Assoungou H, Ondélé R, Miguel L, Mokondjimobé E, Ongoka PR, Abéna AA (2016b). Usage traditionnel et étude de la toxicité aiguë et subchronique de l'extrait aqueux des écorces de tiges de S. camptoneura Gilg et Busse chez le rongeur. Afrique Science 12(5):34-42. |
|
|
OECD. (2002).Harmonised Integrated Classification System for Human Health and Environmental Hazards of Chemical Substances and Mixtures 1:5-15 OECD. |
|
|
O'Shea JJ, Gadina M, Siegel RM (2019). Cytokines and Cytokine Receptors. In Clinical Immunology pp. 127-155.e1. |
|
|
Raskov H, Orhan A, Christensen JP, Gögenur I (2021). Cytotoxic CD8+ T cells in cancer and cancer immunotherapy. British Journal of Cancer 124(2):359-367. |
|
|
Samie A, Madzie N (2016). Effects of Combretum hereroense and Canthium mundianum water extracts on production and expression of interleukin-4. African Journal of Traditional, Complementary and Alternative Medicines 14(1):302-309. |
|
|
Saqib U, Sarkar S, Suk K, Mohammad O, Baig MS, Savai R (2018). Phytochemicals as modulators of M1-M2 macrophages in inflammation. Oncotarget 9(25):17937-17950. |
|
|
Zhang J, Onakpoya IJ, Posadzki P, Eddouks M (2015).The Safety of Herbal Medicine: From Prejudice to Evidence. Evidence-Based Complementary and Alternative Medicine 2015:1-3. |
|
|
Zhu J, Yamane H, Paul WE (2010). Differentiation of Effector CD4 T Cell Populations. Annual Review of Immunology 28(1):445-489. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


