Full Length Research Paper

ABSTRACT
Knowledge of existing genetic variability is essential for initiating a successful breeding program. A set of 628 finger millet accessions comprising accessions from the core collection, farmer preferred and improved varieties released in Kenya, Tanzania and Uganda were profiled for nutrient content. Accessions showed very high variability for the different nutrient contents. Local cultivars and varieties released in the ESA region had significantly lower levels of the main essential nutrients (Ca, Fe, Zn) found in finger millet. Country of origin was highly significant for all the nutrients, with accessions from eastern and southern Africa having significantly lower nutrient contents. Grain color was associated with nutrient content with darker grains having higher compared to white colored. All nutrients were positively correlated (P<0.001) to each other. Grain yield was not significantly correlated to any nutrient content. The substantial variability for the grain nutrients observed in the finger millet core collection and local germplasm indicates the possibility for the selection of nutrient-rich accessions for use in the breeding programs.
Key words: Finger millet, micro and macro nutrients, diversity, grain color, glume cover, malnutrition.
INTRODUCTION
Finger millet (Eleusine coracana L. Gaertn.) indigenous to eastern Africa is a staple crop upon which millions of people depend on for food and income in rural households. It is adapted to adverse agro-ecological conditions and require minimal input (Adekunle, 2012). This African native crop probably originated in the highlands of Uganda and Ethiopia, where farmers have been growing it for thousands of years (Hilu and de Wet, 1976b). It ranks fourth in importance among millets in the world after sorghum (Sorghum bicolor), pearl millet (Pennisetum glaucum) and fox tail millet (Setaria italica) (Upadhyaya et al., 2007a). Finger millet is widely cultivated in Africa and south Asia under varied agro- climatic conditions (Dida et al., 2008). In Africa, it is extensively cultivated in Uganda, Kenya, Tanzania, Ethiopia, Rwanda, Burundi, Zambia and Malawi (Mnyenyembe and Gupta, 1998; Obilana et al., 2002). In south Asia, finger millet is widely cultivated in India and Nepal (Upadhyaya et al., 2007b). Wide adaptability (Upadhyaya et al., 2007b), higher nutritional quality (Gopalan et al., 2002), higher multiplication rate and longer shelf life (Iyengar et al., 1945), make finger millet an ideal crop for use as a staple food and for famine reserve. The crop has dual importance as source of food grain as well as straw for fodder. Finger millet is a rich source of calcium (Ca) (344 mg/100 g) which is 5 to 30 times more than in most cereals, making it the richest plant source (Gopalan et al., 2002; Gupta et al., 2017, Ceasar et al., 2018; Kumar et al., 2018). The grain has a fair amount of protein (7.3 g/100 g) (Malleshi and Klopfenstein, 1998; Sharma et al., 2017) and dietary fibre (15-20%) (Chethan and Malleshi, 2007). Finger millet carbohydrate has unique property of slower digestibility making it a food for long sustenance. All these, deficient in most cereals are crucial to human health and growth, qualify finger millet as an important crop against malnutrition.
Intake of diet poor in iron (Fe), zinc (Zn) and protein is the major cause for micronutrient and protein malnutrition. Fe deficiency leads to anemia. About 79% of the pre-school children between 6 and 35 months of age and 56% of women between 15 and 49 years of age are anaemic in poor countries (Krishnaswamy, 2009). Protein deficiency causes retarded physical and mental growth. Zinc deficiency leads to diarrhea, pneumonia and reduced immunity to diseases, and increased infant mortality (Gibson et al., 2008). Deficiencies of Fe and Zn are widespread worldwide (FAO/WHO, 2001; Cakmak, 2008) especially in sub-Saharan Africa and south and southeast Asia (Reddy Belum et al., 2005). In its report, FAO (2008) singled out sub-Sahara Africa as having the highest prevalence of under nutrition in the world, with one in three people being chronically hungry. A large proportion of people in this part of Africa especially in the rural communities are poor and live on a diet composed primarily of staple foods prepared from cereals (Oniang'o et al., 2003).
Finger millet being a promising source of micronutrients and protein (Malleshi and Klopfenstein, 1998) besides energy, can make a contribution to alleviating micronutrient and protein malnutrition (Underwood, 2000). Because of its high nutrient contents, finger millet is gaining importance in east and southern Africa for its potential use in the preparation of a variety of foods such as porridge, bread, biscuits, pastas, instant baby food, and composite flour (Dendy, 1993; Senthil et al., 2005). The high proportion of carbohydrates in form of non-starchy polysaccharides and dietary fibers in finger millet grain helps in reducing cholesterol. Slow release of glucose during digestion makes it suitable for diabetic patients. The nutritional quality of finger millet grain makes it an ideal food for expectant women, breast-feeding mothers, children, the sick, and diabetics (National Research Council, 1996). This in addition to the high quality protein content makes finger millet a “super crop” in nutritional terms.
The most cost effective approach for mitigating micronutrient and protein malnutrition is to introduce finger millet varieties selected and/or bred for increased Ca, Fe, Zn and protein contents. Plant breeding approach scores over others (such as food fortification, micronutrient supplements, dietary strategies and medical interventions) because it complements the existing approaches to combat micronutrient deficiency. It does not require any special program to change the behavior of farmers/consumers. Cultivars rich in Ca, Fe, Zn and protein with farmer preferred grain quality and adaptation traits are readily accepted (Welch and Graham, 2004; Graham et al., 2007; Pfeiffer and McClafferty, 2007; Prasad, 2010). Furthermore consumption of biofortified foods does not have side effects such as change in taste, bioavailability and risk of developing disease usually associated with inorganic fortification and taking of supplements (Bolland et al., 2010; Institute of Medicine, 2011).
Attempts to breed finger millet for enhanced grain micronutrient and protein contents are still in its infancy stage. Exploitation of existing variability among germplasm accessions is the first step and short-term strategy for developing and delivering micronutrient and protein-dense finger millet cultivars to address the micronutrient and protein malnutrition in the target population (Upadhyaya et al., 2010). International Crop Research Institute for Semi-Arid Tropics (ICRISAT) has the world mandate for finger millet research, with some of its core activities being the collection, characterization, preservation and distribution of germplasm. The ICRISAT genebank at Patancheru (India) holds 5940 accessions of finger millet from 23 countries. Using 14 quantitative traits data on these accessions, Upadhyaya et al. (2006) established a core collection in finger millet, which consists of 622 accessions representing geographical regions and biological races from the entire collection. Accessions from Africa (58.7%) and Asia (35.8%) were predominant in the core, while those from America and Europe were represented by 0.8 to 1.1% only. The subsp. coracona accessions were represented by 97.4%, while those from subsp. africana were 2.6% only (Upadhyaya et al., 2010). A set of 590 accessions from the core collection, 30 farmer preferred varieties and 8 released varieties from the east and southern Africa region were used in the study.
The objective of this present research study was to assess the finger millet core collection, farmer preferred varieties and improved varieties released in the east and southern Africa region for grain mineral content and prospects for breeding for increased Ca, Fe, Zn and protein grain content.
MATERIALS AND METHODS
The material for the study consisted of 590 accessions from the finger millet core collection (Upadhyaya et al., 2006), 30 farmer preferred cultivars and 8 released varieties. Of the released varieties, four are released in Kenya (Okhale 1, Nakuru FM 1, Ikhulule and P224), five in Uganda (Ending, Engeny, Gulu E, U15 and P224) and two in Tanzania (U15 and P224). The material was planted in Kenya at the ICRISAT-Kiboko, field station during the 2013 long rains for evaluation and generation of samples for analysis. ICRISAT-Kiboko is located at altitude 960 m above sea level, Latitude 2° 20’ S and Latitude 37° 45’ E. The accessions were planted in an augmented design (Federer, 1961) with released varieties P224 and U15 and farmer preferred varieties, Nakuru FM 1, Engeny and Okhale 1 used as checks. Each test entry was planted in a single row of 4 m length, with inter-row spacing of 0.4 m and intra-row of 0.1 m with two replications. The experimental plots were maintained weed and insect pest free. Fertilization was done at the rate of 20 kg N/ha and 50 kg P/ha at planting and 50 kg N/ha was applied at top dressing 30 days after planting.
In the field, the accessions were evaluated for the following agronomic traits-plant vigor, days to flowering (DAF), plant height, and number of productive tillers. After harvesting they were evaluated for grain related traits-grain color, glume cover and grain yield. Grain from the two replicates of each accession was homogenously mixed and a representative sample taken for analysis. Freshly harvested grain samples were sent to the Central Analytical Services laboratory in ICRISAT, Patancheru, India for the nutrient analysis using atomic absorption spectrophotometer (Sahrawat et al., 2002). Great care was taken to avoid contamination of grain samples during preparation, handling and shipping. Nitrogen content was determined, and converted to protein percentage weight (% wt) by a factor of 6.25 (Jones, 1941).
Analysis of variance was performed using PROC MIXED (sas V 9.4, SAS Institute Inc. 2017), considering both region and country as fixed. In order to pool the data across the countries, individual country variances were modelled to error distribution using restricted maximum likehood (REML) procedure. Pairwise comparison of means was performed for significant region effect. To analyze for association between grain nutrient contents and grain color, the accessions were classi?ed into ?ve categories as Purple Brown, Dark Brown, Copper Brown, Light Brown, and White. For association with geographical origin, the accessions were classi?ed into seven regions of collection as those from Asia, East Africa, Europe, South Africa, USA, West Africa and Unknown. Pearson correlations were estimated among various micronutrients and macronutrients and between the nutrients and different agronomic traits. Based on 10 nutrients (Ca, Cu, Fe, K, Mg, Mn, P, S, Zn and Protein) and 23 country origins, accessions were grouped into different clusters. This was done following Ward’s method based on Euclidean distance matrix using SAS cluster procedure (SAS V9.4) (Spark, 1973; Fundora et al., 2004).
RESULTS AND DISCUSSION
Nutrient content
Very high variability was observed in all the quality traits determined, iron ranged from 1.37 to 30.04 mg/100 g, potassium from 45.0 to 1427. 0 mg/100 g. Magnesium ranged from 53.0 to 217.8 mg/100 g, manganese from 4.40 to 22.07 mg/100 g, phosphorous from 7.10 to 380.3 mg/100 g, sulphur from 54.8 to 191.7 mg/100 g and zinc from 0.04 to 3.73 mg/100 g. Protein content ranged from 0.50 to 10.10% weight with a mean of 5.86% wt. The highest diversity was observed in potassium (σ2 =10721.0), followed by calcium (σ2 =3633.0), phosphorous (σ2 = 2111.0); while the least diversity was observed in Copper (σ2 = 0.08) and Zinc (σ2 = 0.20).
Calcium content
Calcium values ranged from 155.5 to 554.0 mg/100 g with a mean of 321.5 mg/100 g. A number of accessions recorded higher Ca content than levels cited in literature of 350 mg/100 g (United States National Research Council/National Academy of Sciences (1982). Accessions with the highest Ca content included: IE 2008 with 544 mg/100 g, IE 6541 with 483 mg/100 g, IE 3038 with 475 mg/100 g, IE 4491 with 471 mg/100 g, IE 2644 with 468 mg/100 g, KNE 392 with 466 mg/100 g, IE 595 with 466 mg/100 g, KNE 1149 with 466 mg/100 g, IE 3489 with 460 mg/100 g and IE 6013 with 460 mg/100 g (Table 1). The farmer preferred and released varieties Okhale 1 Ending, Nakuru FM 1, P224 and U15 had low Ca values of 230, 248, 293, 311 and 319 mg/100 g, respectively. They all fall below the conventionally accepted Ca values for finger millet and below the accessions mean of 321. 5 mg/100 g. Okhale 1 the variety with the lowest estimated Ca content is a popular variety in western Kenya. Ending variety with the second lowest Ca content is a farmer local variety preferred by many farmers in eastern and northern Uganda.
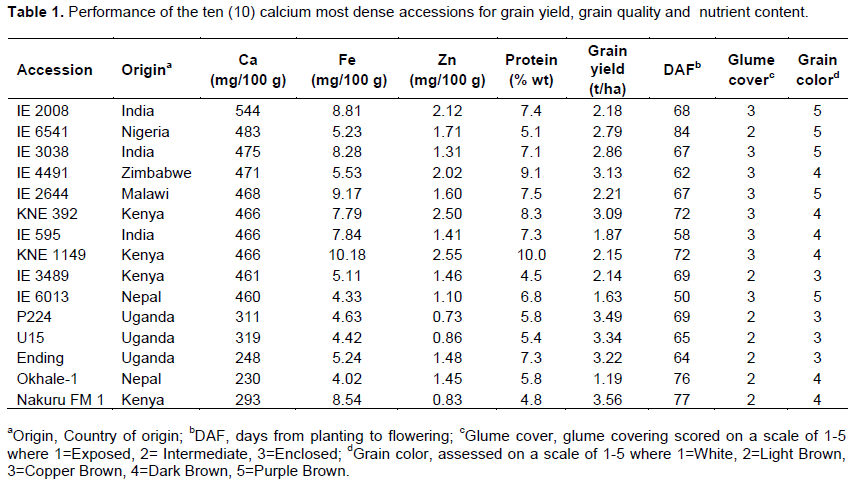
The high Ca content accessions also compared well with the adapted varieties in Zn and Protein content, maturity period (days to flowering (DAF)) and grain yield. These accessions are being tested further for adaptation for possible release and have also been incorporated into the Ca biofortification breeding programs in the region. The use of these accessions as donors is also likely to increase the amounts of Fe and Zn in the progeny. Upadhyaya et al. (2010), working on the same core collection in India found accessions to have Ca range of 386 to 489 mg/100 g with a mean of 430 mg/100 g, which is narrower than was found in this study but falls within the range. The higher range in the present study was mainly due to inclusion of the locally cultivated and released varieties which had low mineral content.
In several studies on estimation of calcium content in different genotypes of ?nger millet, high calcium values have been reported (Sharma et al., 2017). Calcium content was found to vary from 162 to 487 mg/100 g with a mean value of 320.8 mg/100 g grain in 36 genotypes of ?nger millet (Vadivoo et al., 1998), 293 to 390 mg/100 g in six varieties of ?nger millet (Babu et al., 1987), and 50 to 300 mg/100 g in another set of six varieties (Admassu et al., 2009). Furthermore, very high calcium content, 450 mg/100 g (Panwar et al., 2010) and 489 mg/100 g (Upadhyaya et al., 2011) has been reported in few ?nger millet genotypes. These studies are in agreement with the current study that high variability in Ca exists in finger millet germplasm.
Iron content
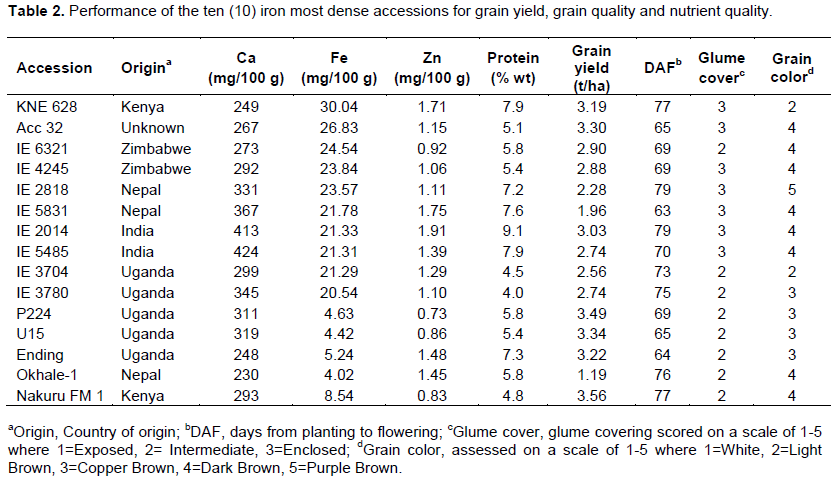
Very high Fe values were estimated for the accessions as compared to the often cited value of 3.9 mg/100 g (United States National Research Council/National Academy of Sciences, 1982). Ten of the best accessions for Fe content had values five times higher than the cited Fe content (Table 2). They include KNE 628 (30.04 mg/100 g), Acc 32 (26.83 mg/100 g), IE 6321 (24.54 mg/100 g), IE 4245 (23.84 mg/100 g), IE 2818 (23.57 mg/100 g), IE 5831 (21.78 mg/100 g), IE 2014 (21.33 mg/100 g), IE 5485 (21.31 mg/100 g), IE 3704 (21.29 mg/100 g) and IE 3780 (20.54 mg/100 g). Released and farmer preferred varieties had values higher than the accepted, but four times lower than those of the high Fe content accessions. Three of the high Fe content accessions Acc 32, KNE 628 and IE 2014 compared favorably well in agronomic traits (yield, DAP, plant height) with the released and farmer preferred varieties and are now being fast tracked for release. Ten high Fe content accessions have been incorporated into the breeding program.
Zinc content
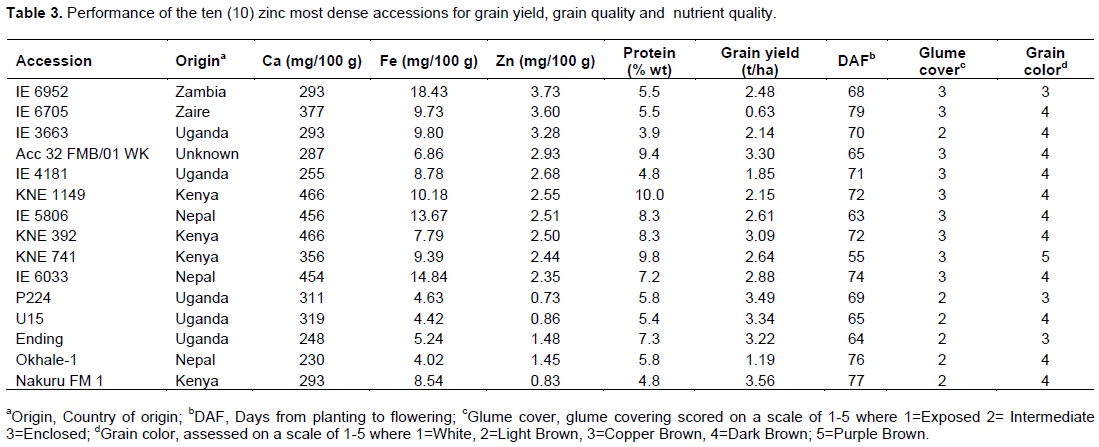
Highest ten Zn content accessions had values ranging from 2.35 to 3.73 mg/100 g with the highest being IE 6952, IE 6705, IE 3663 Acc 32 FMB/01 WK and IE 4181 with concentrations of 3.73, 3.60, 3.28, 2.93 and 2.68 mg/100 g, respectively (Table 3). As with previous nutrients, released and farmer preferred varieties had lower nutrient levels with P224 having the lowest concentration of 0.73 mg/100 g.
The aforementioned results for the micro-nutrients compare well with other crops. CIMMYT Scientists assessed 132 wheat accessions in Mexico and found grain Fe concentrations to range from 2.88 to 5.65 mg/100 g with a mean of 3.72 mg/100 g (Graham et al., 1999; Monasterio and Graham, 2000). Zinc concentrations ranged from 2.52 to 5.33 mg/100 g with a mean of 3.50 mg/100 g. They observed that there was enough genetic variation existing within the wheat germplasm to increase Fe and Zn concentrations substantially in wheat grain. Scientists at IRRI working on 939 rice accessions found Fe concentration to range from 0.75 to 2.44 mg/100 g. Zinc on the other hand ranged from 1.35 to 5.8 mg/100 g (Graham et al., 1999). Within the genotypes tested, there was about a 4-fold difference in Fe and Zn concentrations suggesting genetic potential to increase the concentrations of these micronutrients in rice grain. They also found that the varieties with high Fe concentrations also contained the highest grain-Zn. Additional data demonstrated that high-Fe and high-Zn grain traits are expressed in all rice environments tested and imply the two can be increased concurrently (Welch and Graham, 2004). Researchers at CIAT (Beebe et al., 2000) evaluated a core collection of over 1000 common bean (Phaseolus vulgaris L.) accessions in the field in one season. They found Fe concentrations to range from 3.4 to 8.9 mg/100 g with average of 5.5 mg/100 g (Graham et al., 1999). They observed that there was sufficient genetic variability to increase Fe concentrations significantly (by about 80%) and Zn (by about 50%) in common beans.
Protein content
Protein content ranged from 2.80 to 10.10% weight with a mean of 5.86% wt. Finger millet is reported to have on average 7% protein but large variations in protein content from 5.6 to 12.70% have been noted (Ravindran, 1991; Rao, 1994; Marimurthu and Rajagopalan, 1995; Antony et al., 1996; Vadivoo et al., 1998; Mushtari, 1998; Gautam, 2000; Bhatt et al., 2003). Singh and Srivastava (2006) analyzed 16 finger millet varieties and found out that protein content ranged from 4.88 to 15.58% wt with a mean value of 9.73% wt. Vadivoo et al. (1998) analyzed 36 genotypes of finger millet and reported their protein content in the range of 6.7 to 12.3 mg/100 g with the mean of 9.7 mg/100 g. These show that the protein content in the test material was lower than that reported earlier and need to be enhanced. However, a number of accessions possessed high protein content and are being used as donor parents.
The ?rst step in breeding crops for better nutrition is to evaluate the genetic diversity of available germplasm for target nutritional traits. This study revealed very high variability in the main micro nutrients available in finger millet namely Ca, Fe and Zn. It also revealed that the commonly cultivated varieties are low in these nutrients, which seems to be the trend with other staple crops, like the case of pearl millet in India (Rai et al., 2013). Identified accessions with high nutrients have been used as donors to improve nutrient levels in adapted low nutrient farmer preferred varieties. Accessions IE 2008, IE 6541, IE 3038, IE 4491, IE 2644 and KNE 392 have been incorporated into the biofortification program as donors for Ca. Accessions KNE 628, Acc. 32, IE 6321, IE 4245 and IE 2014 for Fe; and IE 6952, IE 3663, Acc 32 FMB/01 WK, KNE 1149 and IE 5806 for Zn. Three accessions IE 2034, KNE 1149 and KNE 741 are being used as donors for protein.
Nutrient composition by region of collection (origin)
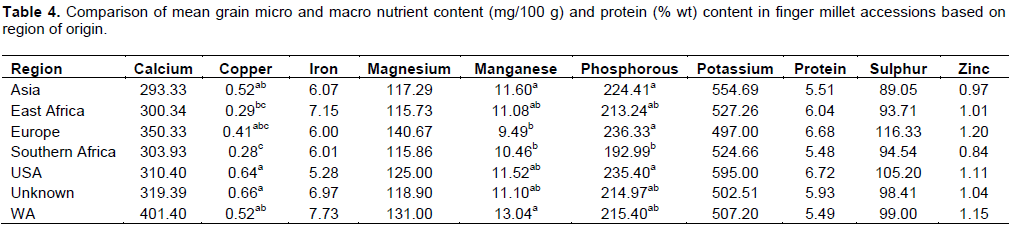
Nutrient concentrations differed with region of collection (Region), though no particular region had accessions superior in all the nutrients (Table 4). Of the main micro nutrient of interest in finger millet (Ca), accessions from West Africa had the highest mean concentration of 401.4 mg/100 g followed by Europe (350.3 mg/100 g) and Unknown (319.4 mg/100 g), while east and southern Africa regions had about the lowest of 300.0 and 303.9 mg/100 g, respectively. Accessions collected from West Africa had the highest mean Fe concentration of 7.73 mg/100 g with east Africa faring well with 7.15 mg/100 g. Not much diversity was expressed in the Zn concentration. Accessions collected from USA had the highest average protein content (6.72% wt) while those from West Africa (5.79% wt) and southern Africa (5.48% wt) had the lowest content (Table 4). Upadhyaya et al. (2010) working on similar material in India found that comparison of mean grain nutrients contents of the accessions classi?ed by geographic origin indicated poor evidence for the relationship of grain nutrient contents with geographical origin. The highest diversity for grain nutrient contents is likely to be present in Africa and Asia, the primary and secondary centers of origin of ?nger millet, respectively (Hilu and de Wet, 1976a; Dida et al., 2008). Barbeau and Hilu (1993) found significantly higher Ca content (515 mg/100 g) in finger millet accessions from Ethiopia compared to those from Kenya (401 mg/100 g) and India (375 mg/100 g).
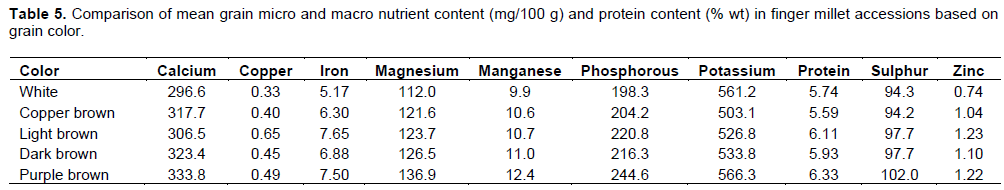
Effect of color on nutrient composition
Mean values of the different nutrient contents showed color to be associated with their concentration, with dark colored grains having highest concentration (Table 5). White colored grains had a mean of 296.6 mg/100 g Ca compared to 333.8 mg/100 g in purple colored grains, 0.33 mg/100 g Cu in white and 0.49 mg/100 g in purple colored while Fe was 5.17 mg/100 g in white and 7.50 mg/100 g in purple brown colored grains. Magnesium was 112.0 mg/100 g and 136.9 mg/100 g, Mn 9.9 mg/100g and 12.4 mg/100 g, P 198.3 mg/100 g and 244.6 mg/100 g in white and purple brown colored grains respectively. On the other hand K had 561.2 and 566.3 mg/100 g, S 94.3 and 102.0 mg/100 g and Zn 0.74 and 1.22 mg/100 g for the white colored and the purple brown colored grains, respectively. Protein content was estimated at 5.74% wt for the white colored grains and 6.33% wt for the purple brown colored grains. In all the nutrients there was a gradual increase in grain nutrient content from white to purple brown colored grains indicating that color can be a good indicator for grain micro and macro nutrient and can be used for selecting for all the nutrient traits. Vadivoo et al. (1998) working on 36 finger millet genotypes reported that the protein content of brown seeded types was higher than white seeded type. Similar findings were reported by Samantaray and Samantaray (1997).
Glume covering
Glume covering was found to be associated with nutrient content. Accessions with enclosed grains had higher nutrient content (Ca=336 mg/100 g, Fe=6.80 mg/100 g, Zn=1.13 mg/100 g and protein=6.00% wt) compared to intermediate covered (Ca=313 mg/100 g, Fe=6.32 mg/100 g, Zn=1.03 mg/100 g and protein=5.60% wt).
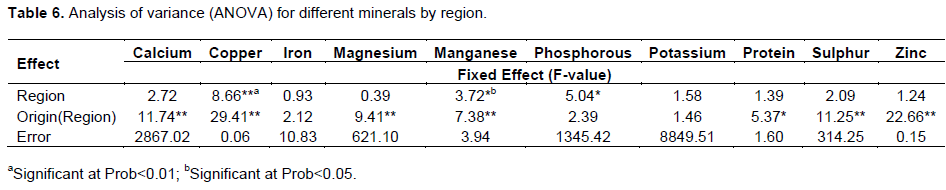
Analysis of variance (ANOVA)
ANOVA revealed country of origin (Country) to be highly significant (P≤0.01) for calcium, copper, magnesium, manganese, sulphur and zinc. It was significant (P≤0.05) for protein and not significant for Manganese (Table 6). Region was highly significant (P<0.01) for copper and was significant at P<0.05 mangenese and phosphorous. Lee et al. (2016), working on variability of nutrient composition of cereal grains from different origin found that country of origin was significant (P<0.05) for all the nutrients and concluded that, source of material was important when considering nutrient composition.
Trait correlation
Correlation analysis was performed among the nutrients and between the nutrients and grain related traits. Highly significant (P<0.001) values were observed among the nutrients calcium, copper, iron, potassium, magnesium, manganese, potassium, sulphur, zinc, and protein (Table 7). Ngu’ni et al. (2011) working on southern African sorghum observed significant positive correlations between grain minerals. Quoting Kumar et al. (2010) who concluded that it indicated that either genetic factors for each pair of minerals are associated, or physiological mechanisms were interconnected for their uptake/ translocation in the grains. Positive correlation between Fe and Zn grain content have also been established in pearl millet (Velu et al., 2007, 2011; Gupta et al., 2009; Govindaraj et al., 2013; Rai et al., 2015; Upadhyaya et al., 2016). This implies that there is potential of simultaneous genetic improvement for two or more grain minerals. Grain color was highly correlated (P≤0.01) to iron, zinc and protein, with no significant association to calcium, indicating that color can be used as morphological marker for the nutrients. The low nutrient content in the farmer preferred varieties and the new released varieties is likely a result of selection against the dark colored varieties as most farmers prefer light and copper brown colored grains. Glume covering was highly correlated to calcium, zinc and protein but not correlated to iron. It was also highly correlated to grain color, an indication that grain color and glume covering can both or interchangeably be used for selecting.
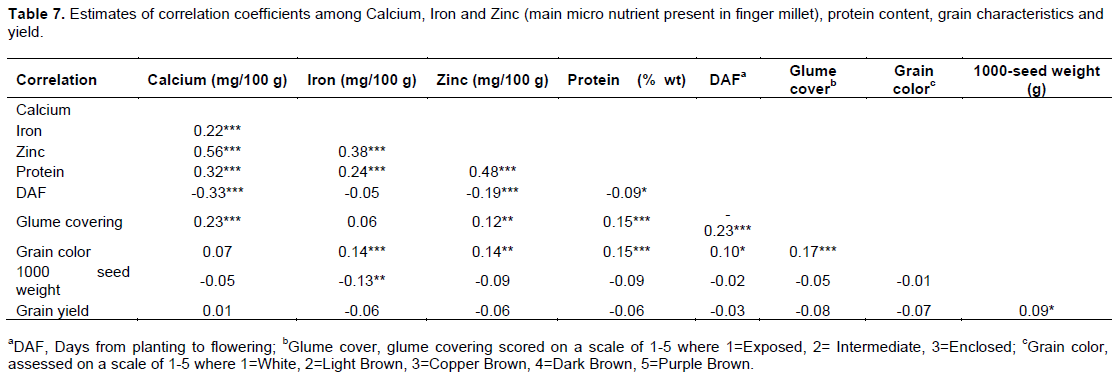
Weak positive and negative correlation exists between grain yield and calcium; and grain yield and iron, zinc and protein, respectively, implying that breeding for improved nutrient content of the micro and macro nutrients will not affect yield and vice versa. Significant negative correlation was established between DAF and Ca (-0.33), Zn (-0.19), and protein (-0.09) and not with Fe (-0.05). Seed size (1000-seed weight) had a negative association with all the nutrients of interest in finger millet, but the association was only significant with Fe (-0.13). Earlier studies have demonstrated the weak or negative correlation between grain yield and grain nutrient traits in different crops. Upadhyaya et al., (2010) working on core finger millet collection in India found weak and non-signi?cant correlations between grain yield and Fe (0.03), Zn (0.05), Ca (0.001) and protein (0.09) contents. Bänzinger and Long (2000) found weak relation between grain Fe and Zn contents in maize with yield. Kumar et al. (2010) and Ng’uni et al. (2011) demonstrated Fe and Zn weak negative relationships with yield in sorghum. Both crops are cereal staple crops in sub Saharan Africa. In pearl millet, a close relative of finger millet no significant association has been established between Fe and Zn grain content; and grain yield and seed size (Velu et al., 2007, 2011; Gupta et al., 2009; Govindaraj et al., 2013). This implies you can increase Fe and Zn grain contents without interfering with grain yield.
The positive correlation among the nutrients implies that several nutrients can be improved concurrently. On the other hand, the weak or lack of association between the different nutrient values and yield related traits (grain yield and seed size) allows for selection for nutrient content without necessarily affecting yield and vice versa. These findings do show that biofortification of finger millet grain can be done easily without comprising yield. According to FAO/WHO (2013), conventional breeding research has demonstrated that micronutrient density can be increased in food staples without negative effects on other farmer-preferred traits. HarvestPlus has developed high Fe pearl millet by conventional breeding (HarvestPlus, 2009). In India, Fe content of a commercially cultivated pearl millet variety was increased by more than 9% and had 11% more grain yield than the parental control (Rai et al., 2013). Rai et al. (2014) screened seed parent progenies and restorer parent progenies in a biofortification program and found that mean Fe content in the progenies increased by 5 to 66% compared to the control cultivars.
The present work is in agreement with previous studies in the different crops that (1) there is high variability with germplasm for increased nutrient content, (2) selecting for one of the nutrients will most likely lead to increase of the other micro nutrients of interest (Ca, Fe and Zn), and (3) selecting for high yield will not necessary lead to reduction of the micro nutrients and vice versa.
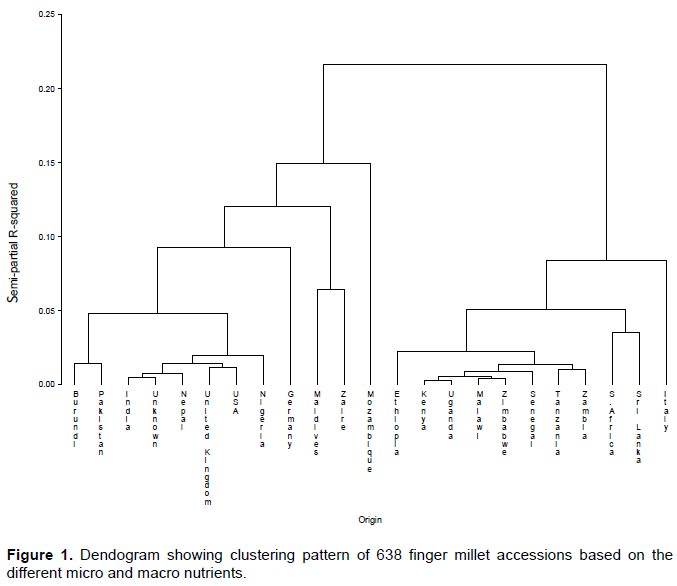
Cluster analysis
To establish the level of diversity among the accessions, cluster analysis was done and five clusters were established (Figure 1). Cluster one consisted of varieties from Burundi and Pakistan; cluster two from India, Unknown, Nepal, United Kingdom, USA and Nigeria; cluster three from Maldives and Zaire; Cluster four had accessions from Ethiopia, Kenya, Uganda, Malawi, Zimbabwe, Senegal, Tanzania and Zambia; and cluster five accessions from South Africa and Sir Lanka. There were however two main clusters; the largest, cluster four consisted of varieties from countries of finger millet origin, Uganda and Ethiopia (Hilu and de Wet, 1976a; Dida et al., 2008) and the main finger millet growing countries in East and Southern Africa region-Tanzania, Kenya, Malawi, and Zimbabwe. The second largest cluster contains accessions from countries from the secondary diversity region and which are among the largest finger millet producers, India and Nepal (Hilu and de Wet, 1976a; Dida et al., 2008). These two clusters represent the largest diversity for the different nutrient traits for future improvement of finger millet. The results are in agreement with accepted phenomena that, the highest diversity usually exists in centers of origin.
The value of biofortified cultivars has been demonstrated by various past researches. The fractional absorption of iron in biofortified pearl millet was found to be similar to a low iron variety when fed to young children in India (Hambidge et al., 2013) and adult women in Benin (Cercamondi et al., 2013). Therefore, the biofortified pearl millet provided significantly more total iron to the individuals who consumed it. Consistent with these results, an efficacy trial completed recently with school children in rural India demonstrated that biofortified pearl millet is efficacious in improving iron status with 64% of the iron deficiency at baseline being resolved in the intervention group after 3 months of daily pearl millet consumption with respect to the low iron pearl millet group. The recommended daily nutrient intake of an adult is 800 g of Ca; 10 mg Fe for males and 15 mg Fe for females; and 15 mg Zn for male and 12 mg Zn for females (FAO/WHO, 2000). With the current accepted finger millet nutrient values of Ca (350 mg/100 g), Fe (3.9 mg/100 g) and Zn (65.9 mg/100 g), Ca and Fe requirements can completely be met and a large portion of Zn by consuming 230 g of finger millet grain daily. With biofortification to the level of the high nutrient accessions found in the germplasm, the amount of grain required will be reduced to 150 g. This represents 35% reduction in the amount of grain required to meet the daily requirement. This can have a huge positive effect in areas with an agriculture-based economy where large segments of the population typically dependent on what they produce.
As biofortification efforts move forward, developing cultivars that have multiple micronutrients should be pursued (Nuss, 2010). For example, common beans and pearl millet already display simultaneous increases in zinc when bred for higher iron concentrations; and quality protein maize often has increased levels of zinc (Nuss, 2011). The synergistic effects between vitamin A and zinc lead to enhanced overall nutrient metabolism (Tanumihardjo et al., 2010). Therefore, a variety that has quality protein, enhanced zinc, and increased provitamin A carotenoids may supply better nutrition than any single nutrient approach for populations that have high intake of the crop. Simulations have demonstrated that adoption of zinc-biofortified rice could readily increase zinc intakes of women and children in Bangladesh (Arsenault et al., 2010). The same held true in adults following a traditional eating pattern in China (Qin et al., 2012). Manipulating both iron and zinc is feasible (Sperotto el al., 2010).
CONCLUSION
A number of conclusions can be made. Accessions showed very high variability for the different nutrient contents. The local cultivars and varieties released in the ESA region had significantly lower levels of the main essential nutrients (Ca, Fe, Zn) found in finger millet. Country of origin was highly significant for all the nutrients, with accessions from eastern and southern Africa having significantly lower nutrient contents. Grain color was associated with nutrient content with darker grains having higher compared to white colored. All nutrients were positively correlated to each other and grain yield was not significantly correlated to any nutrient content. The substantial variability for the grain nutrients observed in the finger millet core collection and local germplasm indicate the possibility for the selection of nutrient-rich accessions for use in the breeding programs.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Adekunle AA (2012). Agricultural innovation in sub-saharan Africa: experiences from multiple stakeholder approaches. Forum for Agricultural Research in Africa, Ghana. ISBN 978-99881. 8373-24. |
|
|
Admassu S, Teamir M, Alemu D (2009). Chemical composition of local and improved finger millet (Eleusine coracana (L.) Gaertn) varieties grown in Ethiopia. Ethiopian. Journal of Health Science 19(1):1-8. |
|
|
Antony U, Sripriya G, Chandra TS (1996). Effect of fermentation on the primary nutrients in finger millet (Eleusine coracana). Journal of Agriculture and Food Chemistry 44(9):2616-2618. |
|
|
Arsenault JE, Yakes EA, Hossain MB, Islam MM, Ahmed T, Hotz C, Lewis B (2010). The current high prevalence of dietary zinc inadequacy among children and women in rural Bangladesh could be substantially ameliorated by zinc biofortification of rice. Journal of Nutrition 140(9):1683-1690. |
|
|
Babu BV, Ramana T, Radhakrishnan TM (1987). Chemical composition and protein content in hybrid varieties of finger millet. Indian Journal of Agricultural Science 57(7):520-522. |
|
|
Banziger M, Long J (2000). The potential for increasing the iron and zinc density of maize through plant breeding. Food and Nutrition Bulletin 21(4):397-400. |
|
|
Barbeau WE, K. Hilu W (1993). Protein, calcium, iron, and amino acid content of selected wild and domesticated cultivars of finger millet. Plant Foods for Human Nutrition 43(2):97-104. |
|
|
Beebe S, Gonzalez A, Rengifo J (2000). Research on trace minerals in the common bean. Food and Nutrition Bulletin, 21(4):387-391. |
|
|
Bhatt A, Singh V, Shrotria PK, Baskheti DC (2003). Coarse Grains of Uttaranchal: Ensuring sustainable Food and Nutritional Security. Indian Farmer's Digest. pp. 34-38. |
|
|
Bolland MJ, Avenell A, Baron JA, Grey A, MacLennan GS, Gamble GD (2010). Effect of calcium supplements on risk of myocardial infarction and cardiovascular events: meta-analysis. BMJ 341:c3691. |
|
|
Ceasar AS, Maharajan T, Ajeesh Krishna TP, Ramakrishnan M, Victor Roch G, Satish L, Ignacimuthu S (2018). Finger millet [Eleusine coracana (L.) Gaertn.] Improvement: current status and future interventions of whole genome sequence. Frontier in Plant Science 9:1054. |
|
|
Cakmak I (2008). Enrichment of cereal grains with zinc: agronomic or genetic biofortification? Plant Soil 302(1):1-17. |
|
|
Cercamondi CI, Egli IM, Mithchikpe E, Tossou F, Zeder C, Hounhouigan JD (2013). Total iron absorption by young women from iron-biofortified pearl millet composite meals is double that from regular millet meals but less than that from post-harvest iron-fortified millet meals. The Journal of Nutrition 143(9):1376-1382. |
|
|
Chethan S, Malleshi NG (2007). Finger millet poly phenols: characterization and their nutraceutical potential. American Journal of Food Technology 2:618-629. |
|
|
Dendy DAV (1993). Opportunities for non-traditional uses of the minor millets. In: Riley, K.W., Gupta, S.C., Seetharam, A., Mushonga, J.N. (Eds.), Advances in Small millets. Oxford and IBH Publishing Co., New Delhi, India. pp. 259- 270. |
|
|
Dida MM, Wanyera N, Dunn MLH, Bennetzen JL, Devos KM (2008). Population structure and diversity in finger millet (Eleusine coracana) germplasm Tropical Plant Biology 1(2):131-141. |
|
|
Federer WT (1961). Augmented designs with one-way elimination of heterogeneity. Biometrics 17(3):447-473. |
|
|
Food and Agriculture Organization (FAO) (2008). The state of food insecurity in the world: High food prices and food security - threats and opportunities. Rome, Italy. |
|
|
Food and Agricultural Organisation/World Health Organisation (FAO/WHO) (2000). Preliminary report on recommended nutrient intakes. Joint FAO/WHO Expert Consultation on Human Vitamin and Mineral Requirements, FAO, Bangkok, Thailand, September 21-30, 1998, revised July 13, 2000. Food and Agricultural Organization of the United Nations Rome, Italy and World Health Organization, Geneva, Switzerland. |
|
|
Food and Agricultural Organisation/World Health Organisation (FAO/WHO) (2001). Human vitamin and mineral requirements, 2nd ed. Geneva, Switzerland. |
|
|
Food and Agricultural Organisation/World Health Organisation (FAO/WHO) (2013). Biofortification: Evidence and lessons learned linking agriculture and nutrition. Geneva, Switzerland. |
|
|
Fundora MZ, Hernandez M, Lopez R, Fernandez L, Sanchez A, Lopez J, Ravelo I (2004). Analysis of the variability in collected peanut (Arachis hypogea L.) cultivars for the establishment of core collections. Plant genetic Resources Newsletter 137:9-13. |
|
|
Gibson RS, Hess SY, Hotz C, Brown K.H (2008). Indicators of zinc status at the population level: a review of the evidence. British Journal of Nutrition 99(Suppl. 3):S14-S23. |
|
|
Gopalan C, Rama Sastri BV, Balasubramanian SC (2002). Nutritive Value of Indian Foods. National Institute of Nutrition, Indian Council of Medical Research, Hyderabad, India. |
|
|
Govindaraj M, Rai KN, Shanmugasundaram P, Dwivedi SL, Sahrawat KL, Muthaiah AR (2013). Combining ability and heterosis for grain iron and zinc densities in pearl millet. Crop Science 53(2): 505-517. |
|
|
Graham RD, Senadhira D, Beebe S, Iglesias C, Monasterio I (1999). Breeding for micronutrient density in edible portions of staple food crops: conventional approaches. Field Crops Research 60(1-2):57-80. |
|
|
Graham RD, Welch RM, Saunders DA, Ortiz Monasterio I, Bouis HE, Bonierbale M, de Haan S, Burgos G, Thiele G, Liria R, Meisner CA, Beebe SE, Potts MJ, Kadian M, Hobbs PR, Gupta RK, Twomlow S (2007). Nutritious subsistence food systems. Advances in Agronomy 92:1-74. |
|
|
Gupta SM, Arora S, Mirza N, Pande A, Lata C, Puranik S (2017). Finger millet: a "certain" crop for an "uncertain" future and a solution to food insecurity and hidden hunger under stressful environments. Frontier in Plant Science 8:643. |
|
|
Gupta SK, Velu G, Rai KN, Sumalini K (2009). Association of grain iron and zinc content with grain yield and other traits in pearl millet (Pennisetum glaucum (L.) R.Br.). Crop Improvement 36(2):4-7. |
|
|
Hambidge MK, Bellad RM, Mahantshetti NS, Kodkany BS, Boy E, Westcott JE, Rai KN, Cherian B, Miller LV, Krebs NF (2013). Bioavailability of iron (Fe) and zinc (Zn) from Fe and Zn biofortified pearl millet. The FASEB Journal |
|
|
HarvestPlus (2009). Breeding Crops for Better Nutrition: General Brochure. Washington, DC: HarvestPlus. |
|
|
Hilu KW, de Wet JMJ (1976a). Domestication of Eleusine coracana. Economic Botany 30(3):199-208. |
|
|
Hilu KW, de Wet JMJ (1976b). Racial evolution of finger millet, Eleusine coracana. American Journal of Botany 63(10):1311-1318. |
|
|
Institute of Medicine (US) Committee to Review Dietary Reference Intakes for Vitamin D and Calcium (2011). Dietary Reference Intakes for Calcium and Vitamin D. eds. Ross AC, Taylor CL, Yaktine AL, Del Valle HB. Washington DC: National Academies Press. |
|
|
Iyengar KG, Doraisami LS, Iyengar RS (1945). Ragi (Eleusine coracana). Mysore Journal of Agricultural Sciences 24:33. |
|
|
Jones DB (1941). Factors for converting percentages of nitrogen in foods and feeds into percentages of proteins. United States Department of Agriculture, Washington 183:1-22. |
|
|
Kumar A, Metwal M, Kaur S, Gupta AK, Puranik S, Singh S, Singh M, Gupta S, Babu BK, Sood S, Yadav R (2018). Nutraceutical value of Finger millet (Eleusine coracana (L.) Gaertn.), and their improvement using omics approaches. Frontier in Plant Science 7:1-1447. |
|
|
Kumar AA, Reddy BVS, Sahrawat KL, Ramaiah B (2010). Combating micronutrient malnutrition: Identification of commercial sorghum cultivars with high grain iron and zinc. Journal of SAT Agricultural Research 8(1). (ejournal.icrisat.org) |
|
|
Lee J, Nam DS, Kong C (2016). Variability in nutrient composition of cereal grains from different origins. SpringerPlus 5(1):419. |
|
|
Krishnaswamy K ( 2009). The problem of double burden of disease and its consequences-a brief overview. In: Programme and abstracts, Symposium on Nutritional Security for India-Issues and Way Forward, Indian National Science Academy, New Delhi, 3-4 August, pp. 5-6. |
|
|
Malleshi NG, Klopfenstein CF (1998). Nutrient composition, amino acid and vita- min contents of malted sorghum, pearl millet, finger millet and their rootlets. International Journal of Food Science and Nutrition 49(6):415-422. |
|
|
Marimurthu R, Rajagopalan R (1995). Protein stability in ragi. Madras Journal 82(11):616-617. |
|
|
Mushtari BJ (1998). Nutritive value of Ragi (Eleusine coracana Gaertn) before and after malting. Beverage Food World 25(5):38-42. |
|
|
Mnyenyembe PH, Gupta SC (1998). Variability for grain yield and related traits in finger millet germplasm accessions from Malawi. Africa Crop Science Journal 6(3):317-322. |
|
|
Monasterio I, Graham RD (2000). Breeding for trace minerals in wheat. Food and Nutrition Bulletin 21(4):392-396. |
|
|
National Research Council (1996). Finger millet. pp. 39-58. In F.R. Ruskin (ed.) Lost Crops of Africa. Vol. I: Grains. National Academy Press, Washington, DC. |
|
|
Ng'uni D, Geleta M, Johansson E, Fatih M, Bryngelsson T (2011). Characterization of the Southern African sorghum varieties for mineral contents: Prospects for breeding for grain mineral dense lines. African Journal of Food Science 5(7):436-445. |
|
|
Nuss ET, Tanumihardjo SA (2010). Maize: A paramount staple crop in the context of global nutrition. Compressive Reviews on Food Science and Food Safety 9(4):417-436. |
|
|
Nuss ET, Tanumihardjo SA (2011). Quality protein maize for Africa: Closing the protein inadequacy gap in vulnerable populations. Advances in Nutrition 2(3):217-224. |
|
|
Obilana AB, Manyasa EO, Kibuka JG, Ajanga S (2002). Finger millet blast (fmb) samples collection in Kenya: Passport data, analyses of disease incidence and report of activities. ICRISAT, Nairobi, Kenya. |
|
|
Oniang'o RK Mutuku JM Malaba SJ (2003). Contemporary African food habits and their nutritional and health implications. Asia Pacific Journal of Clinical Nutrition 12(3):231-236. |
|
|
Panwar P, Nath M, Kumar V, Kumar A (2010). Comparative evaluation of genetic diversity using RAPD, SSR and cytochrome P450 gene based markers with respect to calcium content in finger millet (Eleusine coracana L. Gaertn). Journal of Genetics 89(2):121-133. |
|
|
Pfeiffer WH, McClafferty B (2007). HarvestPlus: breeding crops for better nutrition. Crop Science 47:S88-S105. |
|
|
Prasad R (2010). Zinc biofortification of food grains in relation to food security and alleviation of zinc malnutrition. Current Science 98:1300-1304. |
|
|
Qin Y, Melse-Boonstra A, Yuan B, Pan X, Dai Y, Zhou M, Wegmueller R, Zhao J, Kok FJ, Shi Z (2012). Zinc biofortification of rice in China: a simulation of zinc intake with different dietary patterns. Nutrients 4(6):517-28. |
|
|
Rai KN, Patil HT, Yadav OP, Govindaraj M, Khairwal IS, Cherian B (2014). Dhanashakti: a high-iron pearl millet variety. Indian Farming 64(7):32-34. |
|
|
Rai KN, Velu G, Govindaraj M, Upadhyaya HD, Rao AS, Shivade H (2015). Iniadi pearl millet germplasm as a valuable genetic resource for high grain iron and zinc densities. Plant Genetic Resources 13(1):75-82. |
|
|
Rai KN, Yadav OP, Rajpurohit BS, Patil HT, Govindaraj M, Khairwal IS (2013). Breeding pearl millet cultivars for high iron density with zinc density as an associated trait. e-Journal of SAT Agricultural Research 11:1-7. |
|
|
Rao PU (1994). Evaluation of protein quality of brown and white ragi (Eleusine coracana) before and after malting. Food Chemistry 51(4):433-436. |
|
|
Ravindran G (1991). Studies on millets: proximate composition, mineral composition, phytate, and oxalate contents. Food Chemistry 39(1):99107. |
|
|
Reddy Belum VS, Ramesh S, Longvah T (2005). Prospects of breeding for micronutrients and carotene-dense sorghums. e-Journal of SAT Agricultural Research 1(1):1-4. |
|
|
Sahrawat KL, Ravi Kumar G, Rao JK (2002). Evaluation of triacid and dry ashing procedures for determining potassium, calcium, magnesium, iron, zinc, manganese and copper in plant materials. Communications in Soil Science and Plant Analysis 33(1-2):95-102. |
|
|
Samantaray GT, Samantaray BK (1997). X-ray Diffraction Study of Ragi (Eleusine coracana) Starch. Journal of Food Science and Technology 34(4):343-344. |
|
|
Senthil N, Kumari A, Nirmala Joel A, Selvi B, Raveendran TS, Ramamoor-thy K, Ramanathan A (2005). Small Millets for Nutritional Security, first ed. Kalaiselvam Pathipagam, Coimbatore, Tamil Nadu, India. |
|
|
Sharma D, Jamra G, Singh UM, Sood S, Kumar A (2017). Calcium Biofortification: Three Pronged Molecular Approaches for Dissecting Complex Trait of Calcium Nutrition in Finger Millet (Eleusine coracana) for Devising Strategies of Enrichment of Food Crops. Frontiers in Plant Science 7:2028. |
|
|
Singh P, S. Srivastava O (2006). Nutritional composition of sixteen new varieties of finger millet. Journal of Community Mobilization and Sustainable Development 1(2):81-84. |
|
|
Spark DN (1973). Euclidean cluster analysis. Algorithm As. 58. Applied Statistics 22:126-130. |
|
|
Sperotto RA, Boff T, Duarte GL, Santos LS, Grusak MA, Fett JP (2010). Identification of putative target genes to manipulate Fe and Zn concentrations in rice grains. Journal of Plant Physiology 167(17):1500-1506. |
|
|
Tanumihardjo SA, Palacios N, Pixley KV (2010). Provitamin A carotenoid bioavailability: What really matters? International Journal of Vitamins and Nutrition Research 80(4):336-350. |
|
|
Underwood RA (2000). Overcoming micronutrient deficiencies in developing countries: is there a role for agriculture?. Food and Nutrition Bulletin 21(4):356-360. |
|
|
Upadhyaya HD, Gowda CLL, Pundir RPS, Reddy VG, Singh S (2006). Development of core subset of finger millet germplasm using geographical origin and data on 14 quantitative traits. Genetic Resources and Crop Evolution 53(4):679-685. |
|
|
Upadhyaya HD, Gowda CLL, Reddy GV (2007a). Morphological diversity in finger millet germplasm introduced from southern and eastern Africa. Journal of SAT Agricultural Research 3(1):1-3. |
|
|
Upadhyaya HD, Pundir RPS, Gowda CLL (2007b). Genetic resources diversity of finger millet-a global perspective. In: Mgonja, M.A., Lenne, J.M., Manyasa, E., Srinivasaprasad, S. (Eds.), Finger Millet Blast Management in East Africa. Creating Opportunities for Improving Production and Utilization of Finger Millet. Proceedings of the First International Finger Millet Stakeholder Workshop, Projects R8030 and R8445 UK Department for International Development-Crop Protection Programme, 13-14 September 2005, Nairobi. International Crops Research Institute for the Semi-Arid Tropics, Patancheru 502324, Andhra Pradesh, India pp. 90-101. |
|
|
Upadhyaya HD, Ramesh S, Sharma S, Singh SK, Varshney SK, Sarma NDRK, Ravishankar CR, Narasimhuhu Y, Reddy VG, Sahrawat KL, Dhanalakshmi TN, Mgonja MA, Parzies HK, Gowda CLL, Singh S (2011). Genetic diversity for grain nutrient contents in a core collection of finger millet (Eleusine coracana (L.) Gaertn.) germplasm. Field Crops Research 121:42-52. |
|
|
Upadhyaya HD, Reddy KN, Ahmed MI, Ramachandran S, Kumar V, Singh S (2016). Characterization and genetic potential of African pearl millet named landraces conserved at the ICRISAT genebank. Plant Genetic Resources 15(5):438-452. |
|
|
Upadhyaya HD, Sarma NDRK, Ravishankar CR, Albrecht T, Narasimhudu Y, Singh SK, Varshney SK, Reddy VG, Dwivedi SL, Wanyera N, Oduori COA, Mgonja MA, Kisandu DB, Parzies, HK, Gowda CLL ( 2010). Developing a mini-core collection in finger millet using multi-location data. Crop Science 50(5):1924-1931. |
|
|
Vadivoo AS, Joseph R, Garesan NM (1998). Genetic variability and calcium contents in finger millet (Eleusine coracana L.Gaertn) in relation to grain colour. Plant Foods and Human Nutrition. 52(4):353- 364. |
|
|
Velu G, Rai KN, Muralidharan V, Kulkarni VN, Longvah T, Raveendran TS (2007). Prospects of breeding biofortified pearl millet with high grain iron and zinc content. Plant Breeding 126(2):182-185. |
|
|
Velu G, Rai KN, Muralidharan V, Longvah T, Crossa J (2011). Gene e?ects and heterosis for grain iron and zinc density in pearl millet (Pennisetum glaucum (L.) R. Br). Euphytica. 180(2):251-259. |
|
|
Welch RM, Graham RD (2004). Breeding for micronutrients in staple food crops from a human nutrition perspective. Journal of Experimental Botany 396(55):353-364. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


