Full Length Research Paper

ABSTRACT
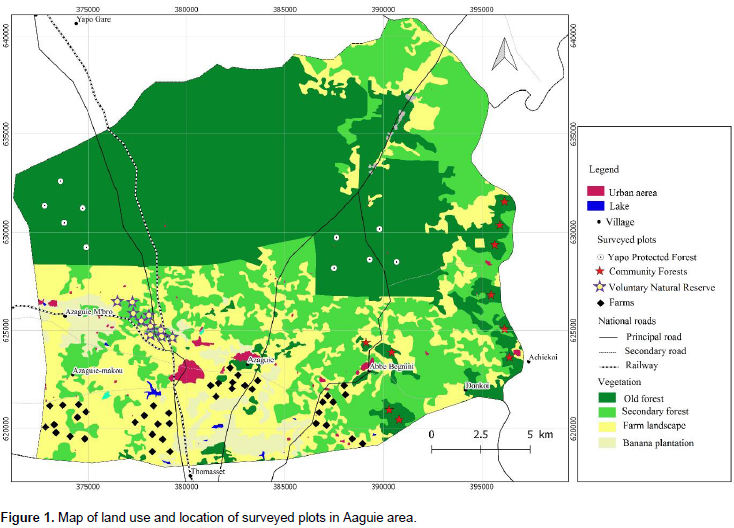
The potential of agricultural landscapes for tree species diversity conservation in perennial crop was examined in the Southeast of Côte d’Ivoire. Based on botanical survey of trees and shrubs species, the beta diversity has been assessed and diversity profile, species accumilation curves, and rank abundance have been compared from 70 plots of 500 m2 selected in the Yapo Protected Forest (YPF), a Voluntary Natural Reserve (VNR), the Community Forests (CF) and the four main crop systems: cocoa (Theobroma cacao), cola (Cola nitida), teck (Tectona grandis) and rubber (Hevea brasiliensis). Results show that 7.3% of stems recorded in old forests can be met in all types of farm habitats. The YPF is more diverse than the other habitats. VNR, CF, and cola-cocoa-rubber farms have the same trees species diversity level when we considered only the most abundant spceies. Farms of rubber contribute to decreasing tree species diversity. In all habitat types, most species are scarce. The highly abundant species are non pioneer species in YPF and VNR, pioneer species in CF and exotic species in farms. A substantial number of tree species can be found on farms that is increasing beta diversity in the study area. Further researchs are required to determine the drivers of these results in the study area.
Key words: Agroforestry, diversity profile, farmland, old growth forest, Côte d’Ivoire.
INTRODUCTION
METHODOLOGY
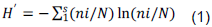
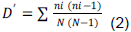
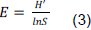
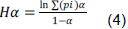
RESULTS
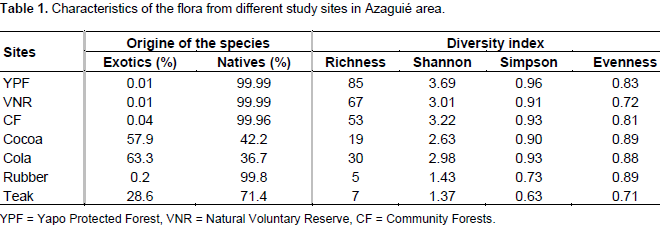
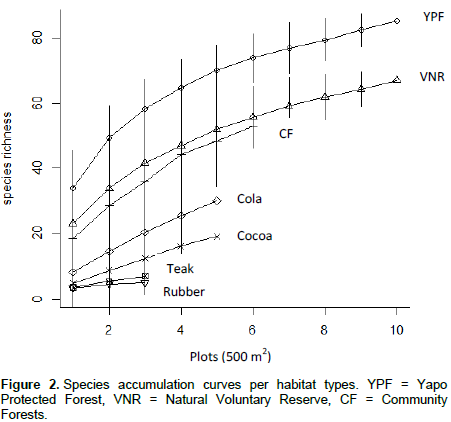
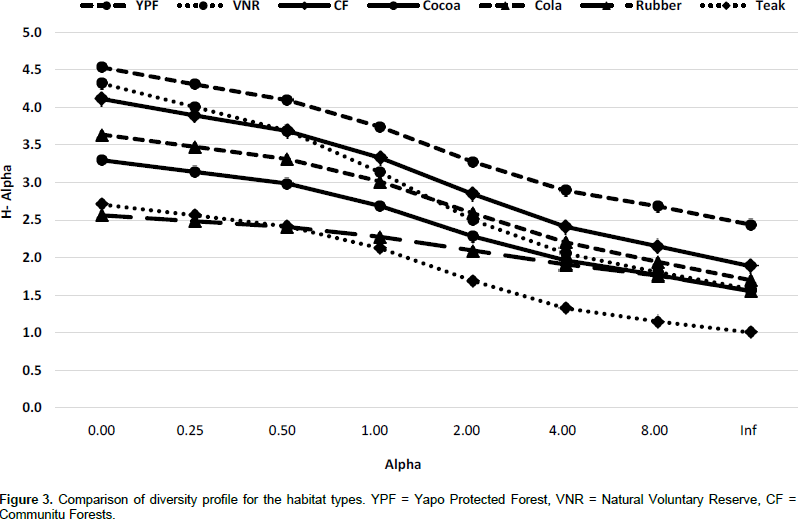
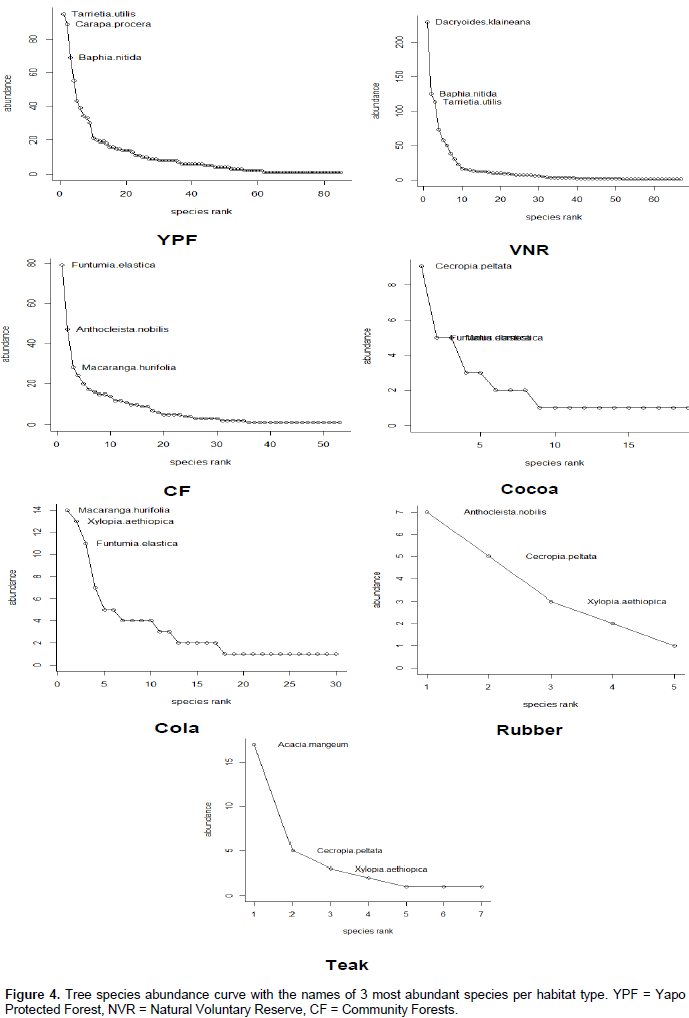
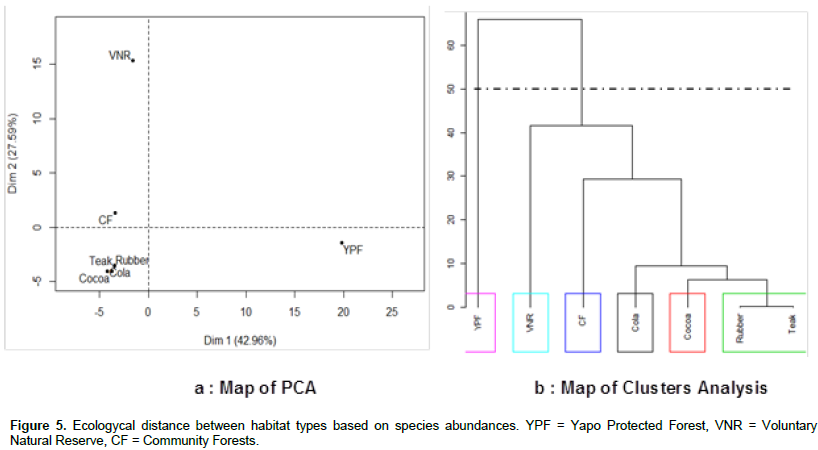
DISCUSSION
CONCLUSION
CONFLICT OF INTERESTS
REFERENCES
|
Adou Yao CY (2005). Pratiques paysannes et dynamiques de la biodiversité dans la forêt classée de Monogaga (Côte d'Ivoire). Thèse Doctorat unique, Département Hommes Natures et Société, Université MNHN, Paris, 233p. |
|
|
Adou Yao CY, Kpangui KB, Vroh BTA, Ouattara D (2016). Pratiques |
|
|
Adou Yao CY, Kpangui KB, Koffi BJC, Vroh BTA (2015). Farming practices, diversity and utilizations of associated species of cocoa plantations in a forest savannah transition zone, Center Côte d'Ivoire. Glob. J. Wood Sci. For. Wildlife 3(3):094-100. |
|
|
Aké-Assi L (2002). Flore de la Côte d'Ivoire 2, catalogue, systématique, biogéographie et écologie. Genève, Suisse : Conservatoire et Jardin Botanique de Genève. Boisseria 58:1-441. |
|
|
Albertin A, Nair PKR (2004). Farmers perspectives on the role of shade trees in coffee production systems: an assessment from Nicoya Peninsula, Costa Rica. Hum Ecol. 32:443-463. |
|
|
Alexandre DY (1982). Aspects de la régénération naturelle en forêt dense de Côte-d'Ivoire. Candollea 37:579-588. |
|
|
Arnold JEM, Dewees PA (1995). Tree Management in Farmer Strategies: Responses to Agricultural Intensification. Oxford University Press, London, U.K. |
|
|
Arnold JEM (1997). Framing the issues. In: Arnold JEM, Dewers PA, eds. Farms, trees and farmers: responses to agricultural intensification. London (UK), pp. 3-20. |
|
|
Berger WH, Parker FL (1970). Diversity of Planktonic foraminifera in deep sea sediments. Science 168:1345-1347. |
|
|
Bernhardt-Reversat F, Hutel H, Lemée G (1978). La fortêt sempervirente de basse Côte d'Ivoire. In Lamotte M, Bourliès F (eds) Problèmes d'écologie: Structure et fonctionnement des écosystèmes terrestres. Masson pp. 313-345. |
|
|
Bhagwat SA, Willis KJ, Birks HJB, Whittaker RJ (2008). Agroforestry: A refuge for tropical biodiversity? Trends Ecol. Evol. 23:261-264. |
|
|
Boffa JM, Kindt R, Katumba B, Jourget JG, Turyomurugyendo L (2008). Management of tree diversity in agricultural landscapes around Mabira Forest Reserve, Uganda. Afr. J. Ecol. 46(1):24-32. |
|
|
Carlo TA, Collazo JA, Groom MJ (2004). Influence of fruit diversity and abundance on bird useof two shaded coffee plantation. Biotropica 36:602-614. |
|
|
Chisholm AR, Muller-Landau HC, Rahman KA, Bebber DP, Bin Y, Bohlman SA, Bourg NA, Brinks J, Bunyavejchewin S, Butt N, Cao H, Cao M, Cardenas D, Chang LW, Chiang JM, Chuyong G, Condit R, Dattaraja HS, Davies S, Duque A, Fletcher C, Gunatilleke N, Gunatilleke S, Hao Z, Harrison RD, Howe R, Hsieh CF, Hubbell SP, Itoh A, Kenfack D, Kiratiprayoon S, Larson AJ, Lian J, Lin D, Liu H, Lutz JA, Ma K, Malhi Y, McMahon S, McShea W, Meegaskumbura M, Razman SM, Morecroft MD, Nytch CJ, Oliveira A, Parker GG, Pulla S, Punchi-Manage R, Romero-Saltos H, Sang W, Schurman J, Su SH, Sukumar R, Sun I-F, Suresh HS, Tan S, Thomas D, Thomas S, Thompson J, Valencia R, Wolf A, Yap S, Ye W, Yuan Z, Zimmerman JK (2013). Scale-dependent relationships between tree species richness and ecosystem function in forests. J. Ecol. 1-11. |
|
|
Cissé A, Aka JCK, Kouamé D, Vroh BTA, Adou Yao CY, N'guessan KE (2016). Caracterisation Des Pratiques Agroforestieres A Base De Cacaoyers En Zone De Foret Dense Semi-Decidue: Cas De La Localite De Lakota (Centre - Ouest, Cote d'Ivoire). Eur. Sci. J. 12(21):50-69. |
|
|
Collinet F (1997). Essai de regroupement des principales espèces structurantes d'une forêt dense humide d'après l'analyse de leur répartition spatiale (Forêt de Paracou, Guyane). Thèse de Doctorat. Université Claude Bernard, Lyon, France. |
|
|
Correia M, Diabaté M, Beavogui P, Guilavogui K, Lamanda N, de Foresta H (2010). Conserving forest tree diversity in Guinée Forestière (Guinea, West Africa): the role of coffee-based Agroforests. Biodivers. Conserv. 19:1725-1747. |
|
|
Corthay R (1996). Analyse floristique de la forêt sempervirente de Yapo (Côte d'Ivoire).Travail de diplôme, Université de Genève 152p. |
|
|
Gibson L, Lee TM, Koh LP, Brook BW, Gardner TA, Barlow J, Peres CA, Bradshaw CJA, Laurance WF, Lovejoy TE, Sodhi NS (2011). Primary forests are irreplaceable for sustaining tropical biodiversity. Nature 478:378-381. |
|
|
Gotelli NJ, Colwell RK (2001). Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecol. Lett. 4:379-391. |
|
|
Guillaumet JL (1967). Recherche sur la végétation et la flore de la région du Bas Cavally (Côte d'Ivoire). Mémoire ORSTOM 20. Paris, |
|
|
Kent M, Coker P (1992). Vegetation description and analysis: A practical approach. London: Belhaven Press. |
|
|
Khan F (1982). La reconstitution de la forêt tropicale humide Sud-ouest de la Côte d'Ivoire. ORSTOM, Collection mémoires, Paris, 151p. |
|
|
Kindt R, Coe R (2005). Tree diversity analysis. A manual and software for common statistical methods for ecological and biodiversity studies. Nairobi: World Agroforestry Centre (ICRAF). 196p. |
|
|
Kindt R, Van Damme P, Simon AJ (2006). Tree diversity in western Kenya: Using profiles to characterise richness and evenness. Biodivers. Conserv. 15:1253-1270. |
|
|
Kpangui KB, Vroh BTA, Goné Bi ZB, Adou Yao CY (2015). Diversités floristique et structurale des cacaoyères du « V Baoulé » : cas de la Sous-préfecture de Kokumbo (Centre, Côte d'Ivoire). Eur. Sci. J. 11(36):40-60. |
|
|
Legendre P, Legendre L (1998). Numerical Ecology. English 2nd edition. Elsevier Scientific Publishing Company, Amsterdam, 213p. |
|
|
Li L (2010). Impact of Human Activities on Vegetation Diversity in Agricultural Ecosystems: Evidence from Fengqiu County, China. J. Resour. Ecol. 1(4):353-360. |
|
|
Lundgren BO, Raintree JB (1982). Sustained agroforestry. In: Nestel B (eds) Agricultural Research for Development: Potentials and Challenges in Asia pp. 37-49. |
|
|
Magurran AE (1988). Ecological diversity and its measurement. Princeton University Press. |
|
|
Margurran AE (2004). Measuring biological diversity. Blackwell Publishing Company, United Kingdom, 256p. |
|
|
Mangenot G (1955). Etude sur les forêts des plaines et plateaux de la Côte d'Ivoire. Etudes éburnéennes 4:5-61. |
|
|
Martinot-Lagarde P (1961). Le Niangon en plantation serrées sous forêt en Côte d'Ivoire. Bois et Forêts des tropiques 80:13-26. |
|
|
Morris EK, Caruso T, Buscot F, Fischer M, Hancock C, Maier TS, Meiners T, Muller C, Obermaier E, Prati D, Socher SA, Sonnemann I, Waschke N, Wubet T, Wurst S, Rillig MC (2014). Choosing and using diversity indices: insights for ecological applications from the German Biodiversity Exploratories. Ecol. Evol. 4(18):3514-3524. |
|
|
Nair PKR (1985). Classification of agroforestry systems. Agrofor. Syst. 3:97-128. |
|
|
Nair PKR (1993). An Introduction to Agroforestry. Kluwer Academic Publishers, The Netherlands, 499p. |
|
|
Ordonez JC, Luedeling E, Kindt R, Tata HL, Harja D, Jamnadass R, van Noordwijk M (2014). Constraints and opportunities for tree diversity management along the forest transition curve to achieve multifunctional agriculture. Curr. Opin. Environ. Sustain. 6:54-60. |
|
|
Pascal J-P, Pélissier R (1996). Structure and floristic composition of a tropical evergreen forest in southwest India. J. Trop. Ecol. 12:195-218. |
|
|
Peet RK (1974). The measurement of species diversity. Ann. Rev. Ecol. Syst. 5:285-307. |
|
|
Perfecto I, Rice RA, Greenberg R, van Der Voort ME (1996). Shade coffee: a disappearing refuge for biodiversity. Bioscience 46:598-608. |
|
|
Purvis A, Hector A (2000). Getting the measure of biodiversity. Nature 405:212-219. |
|
|
Rice RA (2008). Agricultural intensification within agroforestry: the case of coffee and wood products. Agr. Ecosyst. Environ. 128:212-218. |
|
|
Rice RA, Greenberg R (2000). Cacao cultivation and the conservation of biological diversity. Ambio 29:81-87. |
|
|
Salennave P (1961). Niangon de Côte d'Ivoire et Niangon du Gabon. Bois et forêts des Tropiques 76:45-54. |
|
|
Scouppe M (2011). Composition floristique et diversité de la végétation de la zone Est du Parc National de Taï (Côte d'Ivoire). Master Université de Genève 194p. |
|
|
Shaw PJA (2003). Multivariate statistics for the environmental sciences. London: Hodder Arnold. |
|
|
Sinclair FL (1999). A general classification of agroforestry practice. Agrofor. Syst. 46:161-180. |
|
|
Somarriba E, Cerda R, Orozco L, Cifuentes M, Davila H, Espin T, Mavisoy H, Avila G, Alvarado E, Poveda V, Astorga C, Say E, Deheuvels O (2013). Carbon stocks and cocoa yields in agroforestry systems of Central America. Agric. Ecosyst. Environ. 173:46-57. |
|
|
Sonwa DJ, Bernard A, Nkongmeneck A, Weise F, Tchatat M, Akin AA, Marc Janssens JJ (2007). Diversity of plants in cocoa agroforests in the humid forest zone of Southern Cameroon. Biodivers. Conserv. 16:2385-2400. |
|
|
Tchouto GPM (2004). Plant diversity in Central African rain forest: implication for biodiversity conservation in Cameroon. PhD Thesis, Departement of Plant Sciences, Biosystematic Group, Wageningen University, 208p. |
|
|
Van Noordwijk M, Hoang MH, Neufeldt H, Oborn I, Yatich T (2011). How Trees and People can Co-adapt to Climate Change: Reducing Vulnerability Through Multifunctional Agroforestry Landscapes. Nairobi, World Agroforestry Centre (ICRAF). |
|
|
Van Steenis CGGJ (1958). Basic principals of rain forest sociology. Proceedings of the Kandy symposium. UNESCO, Paris, 163p. |
|
|
Vroh BTA, Adou Yao CY, Kouamé D, N'Da DH, N'Guessan KE (2010). Diversités Floristique et Structurale sur le Site d'une Réserve Naturelle Volontaire à Azaguié, Sud-est de la Côte d'Ivoire. Eur. J. Sci. Res. 45(3):411-421. |
|
|
Vroh BTA, Cissé A, Adou Yao CY, Kouamé D, Koffi KJ, Kpangui KB; Koffi BJ (2015a). Relations entre la diversité et la biomasse aérienne des espèces arborescentes dans les agroforêts traditionnelles à base de cacaoyers: Cas de la localité de Lakota (Côte d'Ivoire). Afr. Crop Sci. J. 23(4):311-326. |
|
|
Vroh BTA, Adou Yao CY, Kouamé D, Kpangui KB, Goné Bi ZB, N'Guessan KE (2015b). Trees species diversity and above ground biomass in three tropical forest types in Azaguié area, Côte d'Ivoire. Glob. Adv. Res. J. Plant Sci. 1(2):30-38. |
|
|
Vroh BTA (2013). Evaluation de la dynamique de la végétation dans les zones agricoles d'Azaguié (Sud-est Côte d'Ivoire). PhD Thesis, Université Félix Houphouët-Boigny. Abidjan, 162p. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


