Full Length Research Paper

ABSTRACT
With climatic change, and despite their protection by the Malian forest code, species characterizing the natural forest galleries of rivers in southern Mali are threatened with extinction because of the development of agricultural parcels. A study of seeds germination and the growth of branch cuttings of Pterocarpus santalinoides L’Hér. ex DC. were carried out for multiplication of this typical gallery species. Seeds and branch cuttings were collected from the banks of the river Bagoé. The study revealed that scarification of seeds increases the germination rate (up to 5 times) for P. santalinoides L’Hér. ex DC. The branch budding rate and the total number of buds varied with cutting diameter size. The larger the diameter, the better were the results. Studies such as this one might be carried out on other typical forest gallery species for a better understanding of the functioning of these particular ecosystems with a view to identify how best they might be protected and conserved.
Key words: Southern Mali, woody vegetation, forest gallery, diversity, plant multiplication.
INTRODUCTION
In the Sahelian African countries, plant resources play an important role in the survival of populations. Ecosystems are rich with various plant species and constitute a heritage that deserves to be safeguarded. Unfortunately, they are subject to severe degradation due to the combined effects of climatic hazards and human pressure (MEATEU, 2000).
In southern Mali, socio-economic surveys reveal that a reservoir of biological resources is the forest gallery (Arbonnier, 2000), a part of the closed forest formation according to the classification of Yangambi (Letouzey, 1982).
Before the 1970s, forest galleries played an important role in the protection of the banks, in the fight against the evaporation of river waters and were considered as a reservoir of biological diversity in general and plant species in particular. Due to the favorable water conditions, some plant species are encountered exclusively in forest galleries (Yossi, 1996).
During the last decades, forest galleries in the Sudano - Sahelian zone become increasingly at risk because of agricultural development (Karembé, 1996). The forest galleries are threatened with extinction in favor of agricultural parcels despite their protection by the Malian forest code. This explains, partly, the denudation and filling of several rivers in Mali (Yossi, 1996).
Mali's vegetation has been the subject of several studies, (PIRT, 1983; Boudet, 1979; Penning de Vries and Djitèye, 1982; Breman and De Ridder, 1991; Yossi, 1996; Dembélé, 1996; Sakiliba, 2006). These studies have sought to characterize the country's soil and vegetation resources, namely pastoral resources, specifically woody, or fallow land. However, none of these studies, to our knowledge, has focused on the plant diversity of forest galleries and their eventual restoration. In order to re-introduce the gallery species for the restoration and conservation of the banks and thus participate in the search for an adequate solution toward limiting the threat of development, a study was undertaken on the biology of plant species in the forest galleries of rivers in southern Mali.
The results presented here focused on the multiplication of Pterocarpus santalinoides L’Hér. ex DC. which is of vital importance from both a food and medicinal point of view, (Arbonnier, 2000) and especially for the conservation of the forest galleries. P. santalinoides L’Hér. ex DC. is located in the West Tropical Africa (Senegal to Gabon); South America (Paraguay, Brazil, Bolivia, Peru, Ecuador, Colombia, Venezuela) and the Guyanas (Caribbean – Trinidad) according to (http://tropical.theferns.info/), accessed on May 29, 2018.
Germination was carried out using scarified and non-scarified seeds. No other pre-treatment was used as in (Amusa, 2011) for Afzelia africana Sm. and in (Pathak et al., 2013) for Psidium guajava L. All seeds were from the same location in comparison to the study reported in (Fredrick et al., 2015) for Faidherbia albida (Delile) A.Chev. The growth of branch was done using various cuttings diameter and height in comparison with the value-year-old tree used as parameter in (Ambebe et al., 2017) for the growth responses of Cordia Africana Lam. We recorded the germination score data in order to assess the effect of scarification on the germination rate of P. santalinoides L’Hér. ex DC. The influence of the branch cuttings diameter and height sizes were analyzed on the budding rate and the total number of buds. Seeds germination and growth of branch cuttings of P. santalinoides L’Hér. ex DC. were done in natural conditions in order to better understand the behavior of this species typical of galleries in common environments.
MATERIALS AND METHODS
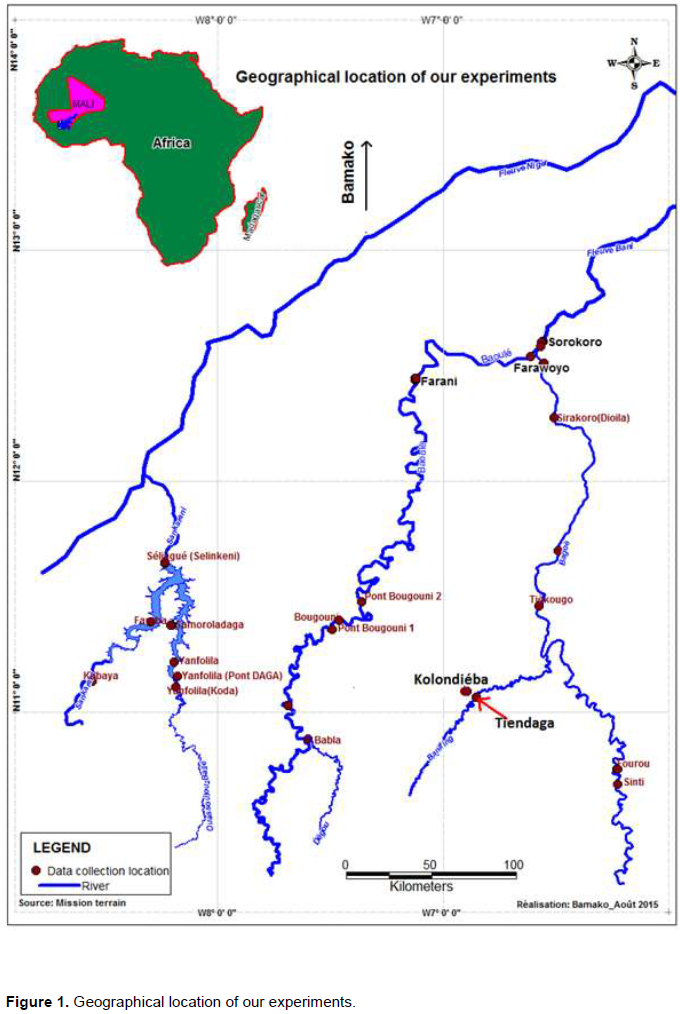
This study was carried out at a site located on latitude 10°03’35’’N, longitude 6°50’59’’W and altitude 311 m above the sea, in the Guinea savannah with an average annual temperature of 27°C. The average rainfall recorded between 1984 and 2014 in the town Konlondiéba, near to the site, is 1090 mm. Seeds of P. santalinoides L’Hér. ex DC. were gathered on August 6, 2012, from the banks of the river Bagoé near the bridge of the village Tiendaga, (Figure 1), using polystyrene bags. The collection did not take into account either the types of formations or the characteristics of the river. Axes were also used to cut the branches of P. santalinoides L’Hér. ex DC. in the banks of the river Bagoé on August 6, 2012. Branch cuttings of different sizes were obtained with secateurs and packed in polystyrene bags. An objective of this study was to introduce the species under fairly natural conditions (Figure 1).
Germination experiment
Germination experiments were carried out with scarified and non-scarified seeds that we termed T1 and T2 treatments, respectively. For T1, a knife was used to make a crack on seed shell. For each type of seed, three parcels of land (repetitions) were used and in each parcel of land 10 pots (replicas) each filled with manure. The manure comprised of 6 wheelbarrows of clay soil from the river Sikoro (Bamako) and 4 wheelbarrows of sifted rubbish heap. Each pot contained a single seed and 60 pots were prepared: 30 for T1 and 30 for T2 seeds. The diameter and height of a pot were 8 cm and 20.5 cm, respectively, filled with 1030 cm3 of manure. After installation, the 60 pots were watered with 30 liters of water using a watering can at 8.00am and at 6.00pm during one year and a half. Observations were then made with each new sprout being noted. For each treatment (scarified and non-scarified seeds) the number of sprouts and their occurrence dates were recorded. Measurements were recorded in a Microsoft Excel spreadsheet.
Branch cuttings experiment
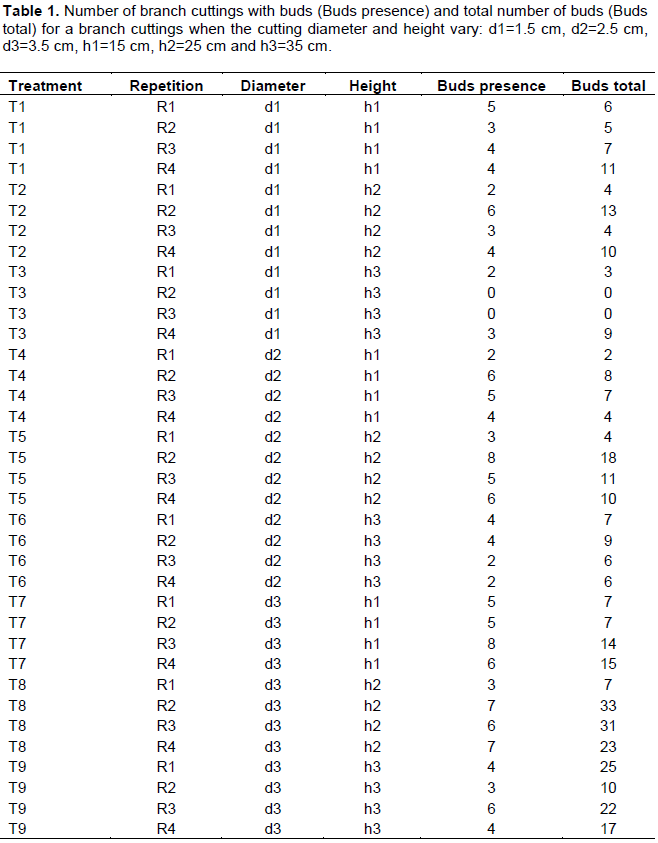
The branch cuttings were measured, sorted and stored on the ground under shade of trees two days before the installation. Two principal factors were studied: the cutting height (h=15, 25 and 35 cm) and the cutting diameter (d=1.5, 2.5 and 3.5 cm). We thus have 9 model treatments (permutations) and adopted the randomized complete block design with 4 repetitions leading to 36 plots. In each parcel of land, 10 pots (replicas) were used, leading to a total of 360 branch cuttings used in the experiment. Each branch of P. Santalinoides L’Hér. ex DC. is in its own bag filled with manure. The manure was made up as follows: 6 wheelbarrows of clay soil from the river Sikoro (Bamako) and 4 wheelbarrows of sifted rubbish heap. We used a soil designed to encourage the development of young plants derived from cuttings. Each bag had 17.5 cm and 35 cm as diameter and height, respectively, filled with 6020 cm3 of manure. The experiment was followed to observe and note new and dead buds. Figure 2 shows a partial image (shot on April 25, 2013) of the branch cuttings experiment with treatment T9 in the front. For each branch cuttings two values were recorded: presence of bud and the total number of buds. For each diameter 120 (4 repetitions x 3 height sizes x 10 replicas) branch cuttings were made. The data were analyzed to understand the budding of Pterocarpus santalinoides L’Hér. ex DC.

Statistical analysis
The data obtained were categorical count values. We used an exact binomial distribution test for the small size germination data (n=30). The proportion test was used to compare percentages of cuttings group’s data. For the larger size cuttings data for which different diameters and/or heights were used (n>50), the normal distribution approximation applies. Hence, we used an analysis of variance (ANOVA) through the linear model framework, in the comparison of branch cuttings data. For the post-hoc analysis, we used the Newman-Keuls procedure with the Student t-test or the proportion test at 5% threshold and the Bonferroni correction. The calculations were performed on the same computer (a laptop equipped with i7-2640M processor, 8GB of RAM, under Microsoft Windows Professional 7) and R version 3.3.3.
RESULTS
Germination
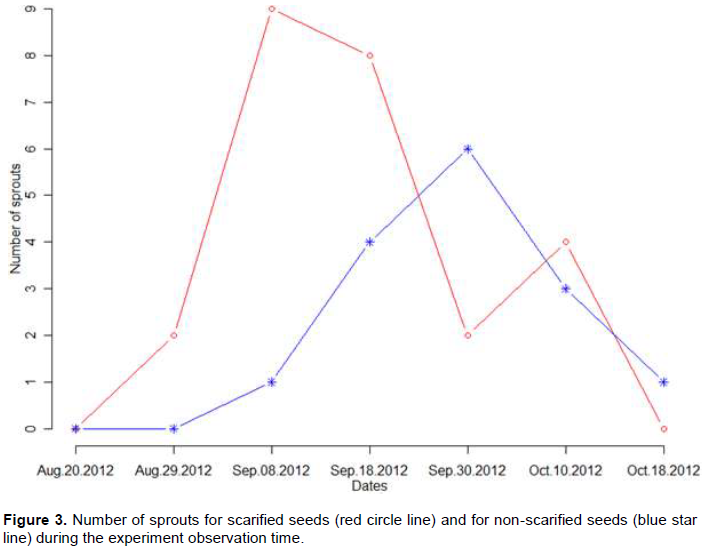
The germination of the seed of P. santalinoides L’Hér. ex DC. is of epigean type. The scarified and non-scarified seeds were sown on the same date. Two scarified seeds germinated 13 days after the sowing date, while one non-scarified seed germinated 30 days after sowing. The data obtained are shown in Figure 3. The measurements from the three parcels of land have been grouped for each treatment. A total of 25 scarified seeds germinated compared to 15 for the non-scarified seeds during the observation period of the experiment leading to a germination rate difference of 33.3%. To show the significance of this difference, we modeled the germination of the scarified seeds using a binomial distribution B(n, p) where n = 30 and p = 25/30. Our null hypothesis is: the germination rate for seeds T1 and T2 are the same. The p-value obtained, p-value=2.62E-5, shows a highly significant difference between the germination of the seeds T1 and T2. The odds ratio or cross-product-ratio was computed (Agresti, 2002): 25*15/15*5 = 5. This value means that the odds of success with scarified seeds are 5 times higher than non-scarified seeds. Figure 3 presents the number of sprouts obtained for scarified and non-scarified seeds. This Figure shows a peak on September 8 for the number of sprouts associated with the scarified seeds. The peak for the number of sprouts associated with the non-scarified seeds appears 22 days later. For comparison purpose, the data reported in (Diop et al., 2012) for the germination of shelled seed of the Jatropha curcas L. were re-analyzed, the p-value obtained was 0.0078. An odds ratio was also calculated and the value obtained was 3.27.
Branch cuttings
For each branch cuttings, two cases were considered to record data:
(a) The presence of bud and
(b) The total number of buds.
The counting data obtained are recorded in Table 1. In this table, values in column "buds presence" are the total number of branch cuttings having at least one bud. The values in column "buds total" are the total number of buds. The 10 replicas of each treatment were grouped for each repetition. The data in Table 1 were used to test the influence of cutting diameter and height on the budding rate (bud’s presence) and the total number of buds observed. An ANOVA was used to test equality of the average presence of buds for the 9 treatments. The p-value obtained, 0.00494, is significant and shows a difference between treatment results. Similarly, an ANOVA was used with the total number of buds. The p-value obtained, 0.000409, is significant and shows a difference between treatment results. We tested separately then together the influence of the diameter and the height on the budding rate on the one hand and on the total number of buds on the other hand.
Influence of the cutting diameter on the budding rate
An ANOVA was used with data in columns “buds presence” and “diameter” of Table 1. The p-value obtained, 0.0105 is significant meaning the budding rate varies with the diameter of the branch cuttings. Using a post-hoc analysis, we obtained two groups: the diameters 1.5 and 2.5 cm forming one and the diameter 3.5 cm the other. The 120 branch cuttings associated with each diameter combined with the values in Table 1 makes it possible to calculate the budding rate per diameter, that is, 30.0, 42.5 and 53.3% respectively for the diameters 1.5, 2.5 and 3.5 cm, respectively. Using the proportion test, we observed a significant difference (p-value=0.0012) between the budding rate for the three branch cuttings diameters. A post-hoc analysis (using proportion test) shows no significant difference (p-value = 0.105) between the budding rates of diameters d1 and d2, while a significant difference (p-value = 0.003) is observed between budding rates of diameters d1 and d3.
From these results, we conclude that cuttings of larger diameter (3.5 cm) have the highest budding rate compared with those of smaller diameter.
Influence of the cutting height on the budding rate
An ANOVA was used with data in columns “buds presence” and “height” of Table 1. The p-value obtained was 0.00895, which means the rate of budding also varies according to the height of the cuttings. Using a post-hoc analysis (t-test), we obtained 2 groups: heights of 15 and 25 cm in one group and height 35 cm forming the other.
The budding rates obtained were 47.5%, 50.0% and 28.3% respectively for the heights 15, 25 and 35 cm. Using the proportion test, significant difference (p-value = 0.00098) was observed between the budding rates associated with cutting heights. The budding rate associated with the cutting height h3 (35cm) is significantly different (proportion test, p-value < 0.0017) to those associated to h1 and d2.
Influence of the cutting diameter and height together on the budding rate
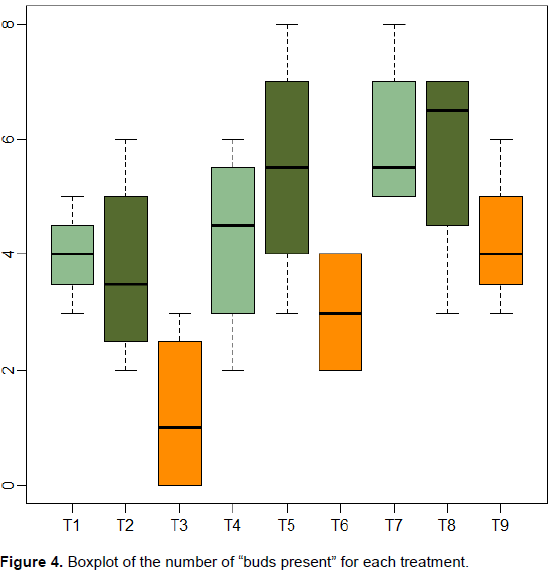
An ANOVA was used to test interaction of cuttings diameter and height on the budding rate. Significant results were obtained for diameter (p=0.004) and for height (p=0.0035) but a non-significant result was obtained (p=0.76) for diameter*height. Positive estimates for model parameters were obtained for diameter (2.0 for d3) and negative estimates for height (-2.75 for h3). Hence, a larger cuttings diameter size increases budding rate while a larger cuttings height penalizes the budding rate. Figure 4 shows the boxplot of the number of the “bud’s presence” for each treatment. Notice that height h3 is used in the treatments T3, T6 and T12. Figure 4 shows clearly a less good result for cutting height h3 compared to those obtained using h1 and h2.
Influence of the cutting diameter on the total number of buds
The total number of buds on each cutting was analyzed.
We observed a significant difference (ANOVA, p = 0.000136) between the average of the total number of buds when the diameter varies. A post-hoc analysis produced 2 groups: the diameters 1.5 and 2.5 cm in one with average values for the total number of buds equal to 6 and 7.7 respectively; and the diameter 3.5 cm forming the other with a average value for the total number of buds equal to 17.6. We can then state that the total number of buds depends strongly on the diameter of the cuttings.
Influence of the cutting height on the total number of buds
The results obtained with ANOVA reveal no significant difference (p = 0.1447) between the average value of the total number of buds when we vary the height from 15 to 35 cm.
Influence of the cutting diameter and height on the total number of buds
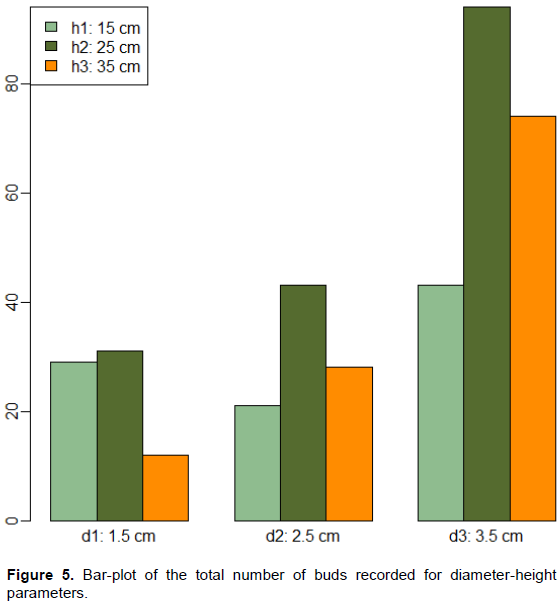
An ANOVA was used to test interaction of diameter and height on the total number of buds. Significant results were obtained for diameter (p=4.93E-5) and for height (p=0.033) but a non-significant result was obtained (p=0.218) for diameter*height. Positive estimate for model parameter was obtained for diameter (3.5 for d3) and negative estimate for height (-4.25 for h3). Hence, a larger cuttings diameter size increases the number of buds and a larger cuttings height acts in opposite direction. The highest model parameter estimate (12.25) was associated with d3*h2. This again revealed the importance of the cutting diameter on the success of the total number of buds. Figure 5 shows the bar plots of the total number of buds for diameter and height sizes. The combination of d3 and h2 seems to be the good choice for multiplication of P. Santalinoides L’Hér. ex DC. when branch cuttings are used.
DISCUSSION
Experimentation on the multiplication of P. santalinoides L’Hér. ex DC. concerning germination and branch cuttings concludes with the following 3 results. First, scarification has a positive effect on germination rate and emergence rate. This result is comparable with that of Hessou et al. (2009), on the germination of seeds of Caesalpinia bonduc (L.) Roxb. in Benin. These authors reported that untreated seeds take longer to germinate (16.4 days), than those removed from their pericarp (13.1 days). It is reported in (Diop et al., 2012), that no significant difference was observed regarding the germination of shelled seeds (87%) and unshelled seeds (73.3%) of the J. curcas L. However, a re-analysis of their data show a concordance with present result.
Second, the budding rate is proportional to the diameter of the P. santalinoides L’Hér. ex DC. branch cuttings. This result is comparable with that of (Ky-Dembélé et al, 2010) on Detarium microcarpum Guill & Perr. where it is reported that in dry environments, budding rates are higher for larger diameter sizes. This results is also comparable with that of (Ky-Dembélé et al., 2016) on the growth of the rooted cuttings of P. santaniloides L’Hér. ex DC. This result is also comparable with those of the in situ cuttings of Lawsonia inermis L. (henna) in which different lengths (15 and 40 cm) were used and where the basal cuttings of height 15 cm gave the best recovery rates (Sanogo et al., 2008).
The present result is also comparable with that for Vitex doniana Sweet (Mapongmetsem et al., 2012), where it is shown that whatever the length of the root segment in question, the budding rate of root cuttings of height 10 cm is higher than that of cuttings of height 15 cm. However, the present results contrast with those of (Stenvall et al., 2003), on the Populus hybrid in which budding decreases with increasing diameter.
Third, budding varies with the diameter of P. santalinoides L’Hér. ex DC. branch cuttings. This result is consistent with that reported by Detrez (1994), where it is shown that large-diameter cuttings yield larger-sized rejects than small-diameter cuttings. The present results are also comparable with that of (Mapongmetsem et al., 2012), for cuttings of the root segments of Vitex doniana Sweet and which maintain that in 24 months after cultivation, cuttings of 1.1 to 2.5 cm diameter (86.0±7.8 %), performed better than those of 0.5 to 1.0 cm (21±1.8%). It is also consistent with the morphological and growth characteristics of J. curcas L. seedlings (Diop et al., 2012). Branch segments close to the stem have a better yield (80%) than the apical segments (50%).
CONCLUSIONS
For P. santalinoides L’Hér. ex DC. scarification has a positive effect on the germination and emergence rates of seeds. The non-scarified seeds takes longer (30 days), to germinate than those scarified (13 days). Branch cuttings of the species is possible under very experimental conditions and the recovery rate is proportional to the diameter of the cuttings. Budding varies with the diameter of the cuttings. The control of the multiplication of woody species, characteristic of gallery, will permit a better protection of the riverbanks against degradation. The multiplication of woody species characteristic of a gallery should be undertaken so that the emerging results will permit the maintenance, protection and restoration of the galleries within the framework of a sustainable management of the natural resources linked to the rivers.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
This work was supported by the Mali Ministry of Higher Education and Scientific Research; the University of technical Sciences and Technologies of Bamako (USTTB); and the program TOKTEN (transfer of knowledge through expatriate nationals). We would like to express our sincere thanks to the people living in the galleries of the South in Mali for their collaboration. We thank S. BROOKS (IGBMC, Strasbourg) for reading and editing the manuscript. We thank A. GOITA (PhD student, FST/USTBB) for help with Figure 1.
REFERENCES
|
Ambebe TF, Akenji MJ, Mogho NMT (2017). Growth responses of branch cuttings of Cordia Africana to physiological age. Journal of Horticulture and Forestry 9(10):91-97. |
|
|
Agresti A (2002). Categorical Data Analysis, 2nd Ed, John Wiley & Sons. pp. 43-47. |
|
|
Amusa TO (2011). Effects of three ore-treatment techniques on dormancy and germination of seeds of Afzelia africana (Sm. Ex pers). Journal of Horticulture and Forestry 3(4):96-103. |
|
|
Arbonnier M (2000). Arbres, arbustes et lianes des zones sèches d'Afrique de l'Ouest. Centre de Coopération Internationale en Recherche Agronomique pour le développement /Muséum national d'histoire naturelle /Union mondiale pour la nature. 1st ed. CIRAD/MNHN/UICN). |
|
|
Breman H, De Ridder N (1991). Manuel sur les pâturages des pays sahéliens, Karthala-A.C.C.T.-Cabo-D.L.O.-C.T.A., 485p. Convention sur la Diversité Biologique Nations Unies 1992. |
|
|
Boudet G (1979). Quelques observations sur les fluctuations du couvert végétal sahélien au Gourma malien et leurs conséquences pour une stratégie de gestion Sylvio-pastorale. Bois et Forêts des Tropiques 184:31-44. |
|
|
Dembélé F (1996). Influence du feu et du pâturage sur la végétation et la biodiversité dans les jachères en zone soudanienne-nord du Mali. Cas des jeunes jachères du terroir de Missira (cercle de Kolokani). PhD. Univ. Aix-Marseille III. 182p. |
|
|
Detrez C (1994). Shoot production through cutting culture and micro grafting from mature tree explants in Acacia tortilis (Forsk.) Hayne subsp. radiana (Savi) Brena. Agroforestry Systems 25:171-179. |
|
|
Diop B, Samba SAN, Akpo LE (2012). Caractéristiques morphologiques et croissance de jeunes plants de Jatropha curcas L. International Journal of Biological and Chemical Sciences 6(2):677-691. |
|
|
Fredrick C, Muthuri C, Ngamau K, Sinclair F (2015). Provenance variation in seed morphological characteristics, germination and early seedling growth of Faidherbia albida. Journal of Horticulture and Forestry 7(5):127-140. |
|
|
Hessou C, Glele Kakaï R, Assogbadjo AE, Odjo T, Sinsin B (2009). Test de germination des graines de Caesalpinia bonduc (L.) Roxb au Bénin. International Journal of Biological and Chemical Sciences 3(2):310-317. |
|
|
Karembé M (1996). Etude de l'évolution de l'occupation des terres du terroir de Gouani en zone soudanien sud du Mali. Master, ISFRA, Mali 81p. |
|
|
Ky-Dembélé C, Tigabu M, Bayala J, Savadogo P, Boussim IJ, Odén PC (2010). Clonal propagation of Detarium microcarpum from root cuttings. Silva Fennica, 44(5):775-787. |
|
|
Ky-Dembélé C, Bayala J, Kalinganire A, Traoré FT, Koné B, Olivier A (2016). Clonal propagation of Pterocarpus santalinoides L'Her. Ex DC.: the effect of substrate, cutting type, genotype and auxin. Southern Forest: a Journal of Forest Science 78(3):193-199. |
|
|
Letouzey R (1982). Manuel de botanique forestière. Afrique tropicale, 2 tomes. Centre Technique Forestier Tropical. Nogent-sur-Marne, 654 p. |
|
|
Mapongmetsem PM, Fawa G, Bellefontaine R (2012). Bouturage des segments racinaires de Vitex doniana Sweet (Verbenaceae): technique de multiplication végétale à faible coût. Int Symp on Tree Product Value Chains in Africa, Yaoundé, Cameroon 26-28 November. |
|
|
MEATEU/DNCN (2000). Former pour une gestion durable des ressources naturelles au Mali. 1ère partie. Etats des lieux: Aspects environnementaux, institutionnels et ressources humaines. Bamako, juillet 2000. |
|
|
Sakiliba M (2006). Rôle des parcs botaniques dans la conservation de la biodiversité végétale dans les pays du Sahel: cas du parc botanique de Bamako. Master IPR/IFRA, 46p |
|
|
Sanogo D, Badji M, Akpo LE (2008). Possibilités de bouturage in situ de Lawsonia inermis L. (henné). Bois et Forêts des Tropiques, 297(3):35-41. |
|
|
Stenvall N, Aarlahti S, Pulkkinen P (2003). The difference between hybrid aspen clones in regeneration from root cuttings and root mass production in different growth circumstances. RitMógilsár Rannsóknasstödvar Skógraektar 17:21-27. |
|
|
Pathak DV, Singh S, Saini RS (2013). Impact of bio-inoculants on seed germination and plant growth of guava (Psidium guajava). Journal of Horticulture and Forestry 5(10):183-185. |
|
|
Penning De Vries FWT, Djiteye MA (1982). La productivité des pâturages sahéliens: une étude des sols, des végétations et de l'exploitation de cette ressource naturelle. PUDOC, Wagueningen, Agric. Res. Rep. (Verel Landbouvk, Onderz) 918, 525p. |
|
|
PIRT (1983). Les ressources terrestres au Mali. Tome I; II et III, TAMS, New York. |
|
|
Yossi H (1996). Dynamique de la végétation post-culturale en zone soudanienne du Mali. PhD thesis. Option Population-Environnement, ISFRA, Bamako, Mali 141 p. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


