Full Length Research Paper

ABSTRACT
This work aimed to evaluate the cytotoxic potential against cancer cells of Passiflora genus plant species cultivated in Brazil and identify the mechanism of cytotoxicity induced by the most promising extract. Ethanolic extracts from the leaves of 14 Passiflora species were obtained by accelerated solvent extraction and in vitro cytotoxicity evaluated against cancer cell lines using MTT assay at a single concentration of 50 μg/ml. Additionally, the IC50 of the Passiflora alata (ELPA) leaf extracts was determined against both cancer (HCT-116, SF-295, OVACAR-8, and HL-60), and non-cancer cells (PBMC). The ELPA flavonoids were identified by HPLC-DAD and UHPLC-MS/MS. The morphological analyses, using light and fluorescence microscopy, and cell cycle and DNA fragmentation analysis, using flow cytometry, were evaluated to study the mechanism of cell death induced by ELPA in HL-60 cells. Among the Passiflora leaf extracts evaluated; ELPA stood out with high cytotoxic activity, followed by Passiflora capsularis and Passiflora quadrangularis with varying high and low cytotoxic activity. ELPA presented high cytotoxic potency in HL-60 (IC50 19.37 μg/ml), and without cytotoxicity against PBMC, suggesting selectivity for cancer cells. The cytotoxic activity of ELPA may well be linked to the presence of ten identified flavonoids. Cells treated with ELPA presented the hallmarks typical of apoptosis and necrosis, with cell cycle arrest in the G2/M phase. From among the studied species, ELPA presented greater cytotoxic activity, possibly a consequence of synergistic flavonoid action, which induces cell death by apoptosis and necrosis.
Key words: Cancer, cytotoxicity, ethanolic extract, Passiflora, Passiflora alata.
INTRODUCTION
Cancer is a progressive disease, characterized by a gradual accumulation of genetic mutations and epigenetic change that causes cellular mechanism imbalances, which progressively transform normal cells into malignant cells (Hedvat et al., 2012; You and Jones, 2012). Together with their associated treatment modalities for curative or palliative purposes, there are more than 100 types of cancers (Ge et al., 2017; Palumbo et al., 2013) . Most therapeutic agents promote programmed cell death (apoptosis) in tumors. However, due to the relative similarity between malignant and normal cells, severe side effects are often observed; this is a limiting factor for therapeutics (Hedvat et al., 2012).
Despite existing treatment modalities, cancer is one of the most common causes of morbidity and mortality worldwide, with estimates that 14.1 million new cancers and 8.2 million cancer deaths occured in 2012, anticipating an increase of at least 70% by 2030, or 20 million new cases of cancer annually by 2025 (Antoni et al., 2016; Ferlay et al., 2015).
Finding more effective and selective compounds that reduce this growing public health problem is a challenge, and nature is the alternative (Dias et al., 2012). Historically, natural plant and animal products have been the origin of most medicinal preparations and natural products continue to provide discovery clues towards pharmacologically active compounds, particularly anti-cancer agents (Dias et al., 2012; Harvey et al., 2015). A continuing study beginning in 1981 through 2014 demonstrated that 83% of the world's registered anticancer drugs were in one form or another either natural products, based therein, or mimetics (Newman and Cragg, 2016). Despite these successes, most plant species have not been systematically investigated (Harvey et al., 2015) , and though Brazil has the greatest diversity of plant species in the world, discoverable cancer cell line cytotoxicities have been poorly studied (Ferreira et al., 2011).
The purpose of this study was to carry on the search for new natural product derived anticancer drugs. Within the gigantic diversity of species of the genus Passiflora, some such as Passiflora edullis Sims (Kuete et al., 2016), Passiflora ligularis Juss (Carraz et al., 2015), Passiflora incarnata (Kapadia et al., 2002; Ma et al., 2014; Sujana et al., 2012), Passiflora molíssima (Chaparro et al., 2015) and Passiflora tetandra (Perry et al., 1991)have been described in the literature as cytotoxic to cancer cell lines, with in vivo chemopreventive or antitumor activity. This raises the hope that related species could exhibit similar activities.
Facing treatments with low curative power, a rising cancer mortality rate in the world, a great variety of plant species not yet having been studied in relation to their antitumor activity, and the fact that certain species of the genus Passiflora present anticancer potential, this study aimed to evaluate the cytotoxic potential of leaf extracts from 14 plant species of the genus Passiflora against human cancer cell lines, and to identify the mechanism of cytotoxicity presented by most promising extract.
MATERIALS AND METHODS
Collection of plant
The leaves of 14 species of Passiflora were collected in July 2011 in the Empresa Brasileira de Pesquisa Agropecuária (EMBRAPA), in the city of Cruz das Almas, Bahia, Brazil (12°04’10”S, 39°06’22”W). The species were registered in the active germoplasm bank (BGP) with access number BGP 08 (Passiflora giberti N. E. Brown), BGP 32 (Passiflora maliformis L.), BGP 77 (Passiflora cincinnata Mast.), BGP 104 (Passiflora vitifolia Kunth), BGP 105 (Passiflora tenuifila Killip), BGP 107 (Passiflora morifolia Mast.), BGP 109 (Passiflora galbana Mast.), BGP 114 (Passiflora mucronata Sessé & Moc.), BGP 125 (Passiflora capsularis L.), BGP 152 (Passiflora suberosa L.), BGP 157 (Passiflora quadrangularis L.), BGP 163 (Passiflora alata Curtis), BGP 170 (Passiflora malacophylla Mast.), BGP 172 (Passiflora racemosa Brot.) and BGP 237 (Passiflora setácea DC.). The leaves of all species were placed in a drying oven at 30°C for seven days and then powdered (only particles between 0.5 and 1.0 mm were utilized for the extractions). The leaves of all species were placed in a drying oven (30°C) for 7 consecutive days and after spraying
Preparation of extracts and samples
Ethanolic extracts of the dried and powdered leaves were prepared by accelerated solvent extraction (ASE 100, Dionex Corporation, Sunnyvale, CA, USA). Each extract was prepared to weigh exactly 6.0 g P. alata, P. capsularis, P. cincinnata, P. gibertii, P. maliformis, P. malacophylla, P. morifolia, P. mucronata, P. quadrangularis, P. racemosa, Passiflora setacea, P. suberosa and 3.0 g of P. vitifolia and P. tenuifila. The plant material was extracted under optimized conditions in five extraction cycles with 64% (w/w) ethanol at 80°C, 1500 psi, and a static cycle timing of 10 min. The cells were then rinsed with fresh extraction solvent (100% of the extraction cell volume) and purged with N2 gas for 60 s. The solvent was removed under reduced pressure at 55°C to yield the corresponding crude extracts, which were stored under refrigeration and protected from light (Gomes et al., 2017). For HPLC analysis, hydroethanolic crude extract of P. alata was purified by solid-phase extraction (SPE) using the typical method with minor modifications (Pereira et al., 2005). A C18 cartridge (Agilent SampliQ, 3 ml/200 mg) was conditioned with 3 ml of methanol, followed by 1 ml of water. Next, 2 ml of a methanolic solution of the sample (5 mg/ml) was added, and the flavonoid fraction was obtained by elution with 3 ml of 60% (w/w) methanol. All samples were prepared and analyzed in triplicate.
Identification of flavonoids in the extract of P. alata
HPLC-DAD
High-performance liquid chromatography (HPLC) fingerprint analysis was carried out using an LC system (Thermo Scientific Dionex Ultimate 3000; MA, USA) consisting of a Thermo Scientific Dionex Ultimate 3000 diode array detector (DAD), quaternary pump, on-line degasser and automatic sampler. The chromatographic separation of sample was achieved on a reversed-phase HPLC column (Waters XBridge™, BEH C18, 100 mm × 3.0 mm I.D., 2.5 μm particle size). For HPLC fingerprint chromatographic analysis, a binary gradient elution system composed of 0.2% (w/w) formic acid in water (solvent A) and acetonitrile (solvent B) was applied as follows: 0 - 30 min, 5 - 20% B. There re-quilibration duration between individual runs was 30 min. The injection volume was 10 µL per sample, the flow rate was 0.6 ml/min and the column temperature was maintained at 30°C. The flavonoids were detected at 337 nm, and the UV spectra of individual peaks were recorded within a range of 190 to 400 nm. Data were acquired and processed with Chromeleon software. The main flavonoids were identified in the extract of P. alata based on their retention times (tR), coinjection of the samples with standards and comparison of their UV adsorption spectra (Gomes et al., 2017).
UHPLC-MS/MS
Ultra-high performance liquid chromatography tandem mass spectrometry (UHPLC-MS/MS) was utilized to identify and confirm chromatogram peaks, which comprised an ACQUITYTM UHPLC system (Waters Corp., Milford, MA, USA). Separation was performed with an ACQUITY BEH C18 column (50 mm × 2 mm, 1.7 µm; Waters, USA). The mobile phase was a mixture of 0.1% formic acid (A) and acetonitrile in 0.1% formic acid (B), with an linear gradient elution as follows: 0-11 min, 5-95% B. The injection volume was 4 µL. The flow rate was set at 0.30 ml/min. The UV spectra were registered from 190 to 450 nm. Eluted compounds were detected from m/z 100 to 1000 using a Waters ACQUITY® TQD tandem quadrupole mass spectrometer equipped with an electrospray ionization (ESI) source in the negative mode using the following instrument settings: capillary voltage 3500V; capillary temperature 320°C; source voltage 5 kV; vaporizer temperature 320°C; corona needle current 5 mA; and sheath gas nitrogen 27 psi. Analyses were run in the full scan mode (100-2000 Da). Ion spray voltage: -4 kV; orifice voltage: -60 V. The identification of flavonoids was based on the comparison of the molecular formula with that of the published data, further confirmation was performed by illuminating the quasi-molecular ions and key flavonoids fragmentations, in particular for those isomeric.
Human cancer cell lines and non-cancer cells
The cytotoxicity of these Passiflora species extracts was tested against colon carcinoma (HCT-116), glioblastoma (SF-295), ovarian adenocarcinoma (OVCAR-8) and promyelocytic leukemia (HL-60) human cancer cell lines, all obtained from the National Cancer Institute, Bethesda, MD, USA. The cells were grown in RPMI-1640 medium supplemented with 10% fetal bovine serum, 2 mM glutamine, 100 µg/ml streptomycin and 100 U/ml penicillin incubated at 37°C in a 5% CO2 atmosphere. Peripheral blood mononuclear cells (PBMCs) were isolated from a sample of about 3 ml of human blood plus 5 ml of saline. The steps up to isolation included the addition of 3 ml of Ficoll followed by 15 min of centrifugation at 1500 rpm, and aspiration of the PBMCs present in the intermediate region between the red blood cells and the plasma. The PBMC suspension was transferred to another tube which was added with saline to the 11 ml volume and centrifuged for 5 min at 1000 rpm. The supernatant was discarded, and the PBMC pellet was re-suspended in complete medium (RPMI 1640 plus 20% fetal bovine serum and 10 μg/ml ConA) and counted in a Neubauer chamber for further dilution and plating. The Research Ethics Committee of the Oswaldo Cruz Foundation (Salvador, Bahia, Brazil) approved the experimental protocol (# 031019/2013).
Determination of the cytotoxic effect of Passiflora extracts on cancer cell lines
The MTT assay was used to determine the cytotoxic effect of the 14 extracts of Passiflora leaves against cancer cell lines (Mosmann, 1983). For all experiments, HCT-116 cells (0.7 × 105 cells/ml in 100 µL medium), SF-295 and OVCAR-8 cells (0.1 × 106 cells/ml in 100 µL medium) were seeded in 96-well plates and incubated in a humidified chamber with 5% CO2 at 37°C for 24 h. After this, the 14 extracts of Passiflora were solubilized in dimethyl sulfoxide (DMSO, 0.7%), and added to each well at the concentration of 50 µg/ml. The cells were then incubated for 72 h at 37°C in a 5% CO2 atmosphere. After 72 h, MTT (0.5 mg/ml) was added, followed by incubation at 37°C in an atmosphere of 5% CO2 for 3 h. After incubation, the formazan product was dissolved in 150 μL DMSO, and absorbance was measured using a multi-plate reader (DTX 880 Multimode Detector, Beckman Coulter Inc.). The treatment’s effects were expressed as the percentage of control absorbance of reduced dye at 595 nm. All absorbance values were converted into a cell growth inhibition percentage (GI %) by the following formula:
[GI% = 100 − [(TE/NC) × 100%]
In which, NC is the absorbance for the negative control and TE is the absorbance in the presence of the tested extracts. The half maximal inhibitory concentration (IC50) was determined for the extract that caused the greatest GI% against HCT-116, SF-295 and OVCAR-8. The same protocol for the same cells, adding the cell line HL-60 (0.3 × 106 cells/ml in 100 µL medium) was used to determine the IC50, varying only the concentration of the compound from 0.39 to 50 µg/ml, and the potency of the extract. The experiment was performed as three independent experiments, and doxorubicin (0.3 μg/ml) was used as a positive control (Doxorubicin, purity > 98%; Sigma Chemical Co., St. Louis, MO, USA).
Determination of the effect of P. alata leaf extract in non-cancer cells
The alamar blue assay was used to determine the effect of extract from the leaves of P. alata (ELPA) against proliferation of non-cancer cells (PBMCs) obtained from peripheral blood from healthy human volunteers (Nakayama et al., 1997). Initially, the cells were plated in 96-well plates (100 μL/well of a solution of 3 × 106 cells/ml). After 24 h of incubation, extract from the leaves of P. alata (0.39 to 50 µg/ml) was dissolved in 0.3% DMSO, added to each well and incubated for 72 h. Doxorubicin (0.03 - 0.25 μg/ml) was used as a positive control and 0.3% DMSO was used as a negative control. 24 h before the end of the incubation period, 20 μL of alamar blue stock solution (0.312 mg/ml) (resazurin) was added to each well. Absorbances were measured at wavelengths of 570 nm (reduced) and 595 nm (oxidized) using a plate reader (DTX 880 Multimode Detector, Beckman Coulter™).
Determination of the cytotoxicity mechanism induced by P. alata leaf extract
Morphological analysis by light microscopy
Differential staining with hematoxylin/eosin was used for morphological analysis. The HL-60 cells, plated at 0.3 × 106 cells/ml were incubated for 72 h with P. alata leaf extract (9.69, 19.77, and 38.74 μg/ml) and examined under an inversion microscope. To observe the morphology, 50 μL of the cell suspension was added to the slide centrifuge (Shandon Southern Cytospin™). After cell adhesion on the slide, fixation with methanol for 1 min was completed, hematoxylin followed by eosin was used. Doxorubicin (0.3 μg/ml) was used as a positive control (Veras et al., 2004).
Morphological analysis by fluorescence microscopy
Acridine orange/ethidium bromide (AO/EB) (Sigma Aldrich) staining in HL-60 cells was performed to evaluate the pattern of cell death induced by P. alata leaf extract. Cells of the HL-60 strain were plated at 0.3 × 106 cells/ml, and then incubated for 72 h with the extract (9.69, 19.77, and 38.74 μg/ml). The cell suspension was transferred to a microtube and centrifuged for 5 min at low speed (500 rpm). The supernatant was discarded and the cells re-suspended in 20 μL of PBS solution. Then 1 μL of the ethidium bromide/acridine orange solution was added to each tube and placed under a fluorescence microscope to observe the cellular events. Doxorubicin 0.3 μg/ml was used as a positive control (Geng et al., 2003).
Analysis of cell cycle and DNA fragmentation
Cell cycle analysis was performed to determine cell content, which reflects the cell cycle phases (G0/G1, S, and G2/M). For this, HL-60 cells were plated in 24-well plates at the concentration of 0.3 × 106 cells/ml and incubated for 72 h with P. alata leaf extract (9.69, 19.77 and 38.74 μg/ml). Doxorubicin (0.3 μg/ml) was used as a positive control. After the incubation period, cells were harvested in a permeabilization solution containing 0.1% triton X-100 (Sigma Chemical Co. St Louis, MO, USA), 2 µg/mL propidium iodide (Sigma Chemical Co. St Louis, MO, USA), 0.1% sodium citrate and 100 µg/mL RNAse (Sigma Chemical Co. St Louis, MO, USA) and incubated in the dark for 15 min at room temperature (Militão et al., 2006). Finally, cell fluorescence was measured by flow cytometry with a BD LSRFortessa cytometer using BD FACSDiva Software (BD Biosciences) and FlowJo Software 10 (FlowJo Lcc; Ashland, OR, USA). Ten thousand events were evaluated per experiment, and cellular debris was omitted from the analysis.
Statistical analysis
Data are presented with mean ± standard error of the mean (SEM), standard deviation (SD), or IC50 values; confidence intervals (CI 95%) were obtained by nonlinear regression. Differences among the experimental groups were compared by one-way variance analysis (ANOVA), followed by Newman-Keuls test (p < 0.05). All analyses were carried out using the Graphpad program (Intuitive Software for Science, San Diego, CA, USA).
RESULTS AND DISCUSSION
Based on the fact that some species of the genus Passiflora have cytotoxic activity against cancer cell lines and that relatively few species have been evaluated for their possible anticancer activity, we selected 14 species of Passiflora cultivated in Brazil, produced by ASE method to evaluate their potential in vitro cytotoxic activity.
The cell growth inhibition percentage (GI%) of the Passiflora extracts was evaluated against three human cancer cell lines (HCT-116, OVACAR-8, and SF-295) in a single concentration of 50 µg/ml and measured by MTT assay after 72 h of incubation (Table 1). The results were analyzed for each cell line tested using a GI% scale as follows: samples with low cytotoxic activity have GI < 50%; samples with moderate cytotoxic activity have GI between 50 and 75%, and samples with high cytotoxic activity have GI > 75% (Mahmoud et al., 2011). The results showed that the extracts of P. cincinnata (3), P. gibertii (4), P. maliforms (5), P. mallacophyla (6), P. murchronata (7), P. morifolia (8) P. racemosa (10), P. setacea (11), P. suberosa (12), P. tenuifila (13) and P. vitifolia (14) have low or nonexistent cytotoxic activity in all human cancer cell lines tested. Extracts from P. capsularis (2) and P. quadrangularis (9) have high activity against HCT-116, low activity against SF-295, and cytotoxic activity from low (P. quadrangulares) to moderate (P. capsularis) against OVACAR-8. The extract from the leaves of P. alata (1) presented high cytotoxic activity in all human cancer cell lines analyzed. Therefore, among the Passiflora leaf extracts evaluated, P. alata (1) stands out with promising cytotoxic activity, followed by P. capsularis (2) and P. quadrangularis (9).
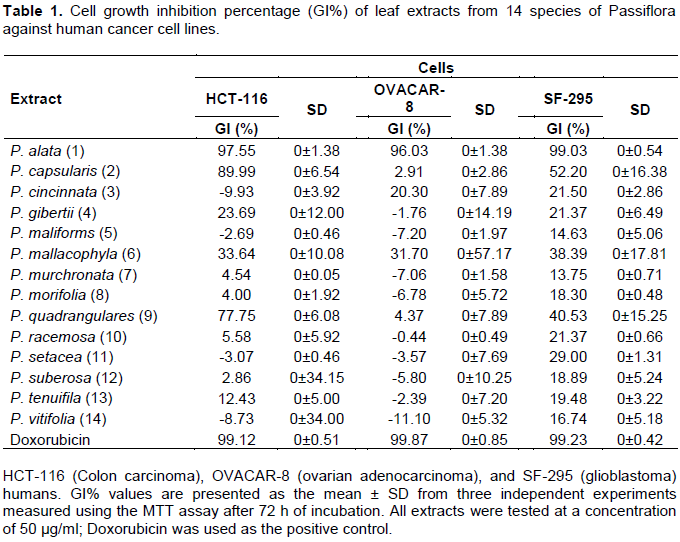
Although none of the three species highlighted in the present study had been previously evaluated for their cytotoxic activity against cancer cells, other species of the genus Passiflora have also been evaluated. The cytotoxic or antiproliferative activity of species of the genus Passiflora was evidenced for the first time by Perry et al. (1991) through study of P. tetandra leaf extract, demonstrating cytotoxic activity against murine leukemic cell line (P388), being possibly mediated by the extract component 4-hydroxy-2-cyclopentane. The ethanolic extract of P. ligularis Juss. and the methanolic extract of the P. edullis fruit have been respectively described as presenting activity against human hepatocellular carcinoma (Hep3B) and leukemic cell line (CCRF-CEM) (Carraz et al., 2015; Kuete et al., 2016).
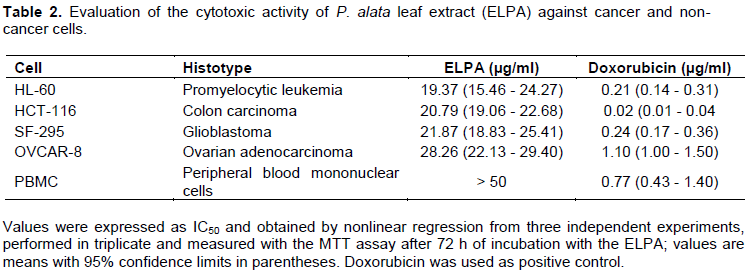
Based on the promising cytotoxic activity observed in the P. alata leaf extract (ELPA), we determined the median inhibitory concentration able to induce 50% of maximal effect (IC50) against four human cancer cell lines (HCT-116, HL-60, OVCAR-8, and SF-295) through the MTT method. The results demonstrated that ELPA has cytotoxic activity in the four cancer cell lines evaluated, respectively being most potent for the cell line HL-60 (IC50 of 19.37 µg/ml), followed by HCT-116 (IC50 of 20.79 µg/ml), SF-295 (IC50 of 21.87 µg/ml), and OVCAR-8 (IC50 of 28.26 µg/ml) (Table 2). According to the preclinical cytotoxic drug screening program based on the US National Cancer Institute program, only extracts with IC50 values below 30 μg/ml in assays with cancer cell lines are considered promising for the development of anticancer drugs (da Silva et al., 2016; Mahmoud et al., 2011). Thus, ELPA presents promising in vitro cytotoxic activity for further study. For a natural compound to progress as a potential anticancer drug candidate, it is necessary to determine the degree of specificity of the drug by evaluating in vitro cytotoxic activity against non-cancer cells (Andrade et al., 2015; da Silva et al., 2016). For this, ELPA was subjected to in vitro cytotoxic activity evaluation against non-cancer cells (PBMC); the results demonstrated that the compound shows no cytotoxic activity against PBMC (Table 2).
The cytotoxic activity of natural bioactive products against cancer cells is attributed to the chemical composition of the product, and this practice of identification is the most successful source of potential drug discovery and development (Dias et al., 2012).
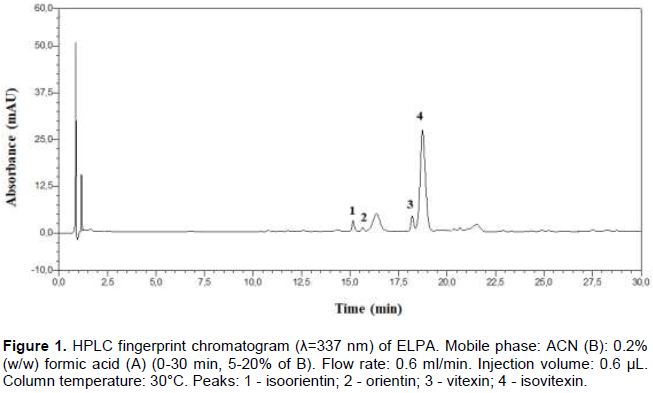
A total of 4 flavonoids were identified by HPLC-DAD, and 8 flavonoids were identified by UHPLC-MS/MS in the negative mode. They are summarized along with their retention time, molecular formula and MS/MS fragments as shown in Table 3. HPLC fingerprint chromatogram of ELPA with 4 identified compounds is shown in Figure 1.
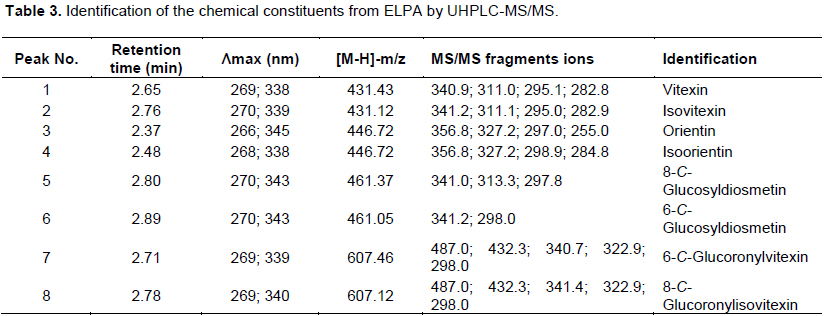
Flavonoid from Passiflora spp. are usually C-glycosylated with one or more sugar units. Different fragmentation patterns were observed in MS/MS experiments for flavone C-glycosides. Based on MS and UV/Vis spectra, all identified flavones were vitexin, isovitexin, orientin, isorientin, 8-C-glucosyldiosmetin, 6-C-glucosyldiosmetin, 6-C-glucoronylvitexin and 8-C-glucoronylisovitexin.
Thus, compounds 1 and 2 (tR = 2.65 and 2.76 min) were identified as vitexin and isovitexin, once the [M-H]- pseudo molecular ions were registerated at m/z 431 and the fragmentation pattern, compared with the literature are also in accordance with these two compounds. In the same way compounds 3 and 4 (tR = 2.37 and 2.48 min) were identified as orientin and isoorientin, also due the pseudo [M-H]- ions at m/z 446. These compounds (1-4) indicated similarities of fragmentation patterns due to dehydration and cross-ring cleavages of the glucose moiety, that is, 0.2 cross-ring cleavage (-120 amu) and 0.3 cross-ring cleavage (-90 amu) (Figure 2) (Davis and Brodbelt, 2004).
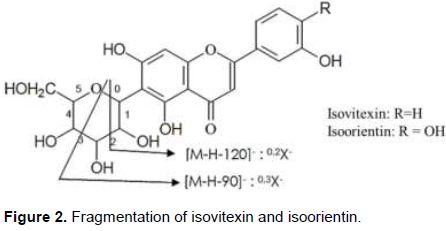
Compounds 5 and 6 (tR = 2.80 and 2.89 min) were identified as 8-C-glucosyldiosmetin and 6-C-glucosyldiosmetin, [M-H]- ions presented at m/z 461. In the MS/MS spectra the fragments were at m/z 341 0.2X-[M-H-120]-, which was attributed to the loss of a glucose moiety.
Compounds 7 and 8 (tR = 2.71 and 2.78 min) exhibited [M-H]- ions at m/z 607. They were identified as 6-C-Glucoronylvitexin and 8-C-Glucoronylisovitexin. In the MS/MS spectrum, the same fragment ion at m/z 487 0.2X-[M-H-120]- indicated the loss of a glucose moiety. The fragment ion at m/z 432 suggesting the loss of a glucuronic acid residue [M-H-175]- (Chen et al., 1998; Schutz et al., 2004; Wolfender et al., 1998).
The extractive process used for the ELPA was ASE, developed to maximize the yield of the extract and simultaneously the content of flavonoids (Gomes et al., 2017). According Pearson et al. (2010) (Pearson et al., 2010) and Saha et al. (2015), the process is promising, because it offers advantages over other extraction processes such as: easy automation, faster sample analysis, better reproducibility, less solvent required, and sample maintenance in an environment without light and free of oxygen. Accelerated extraction systems also allow the operator to control the temperature, pressure, extraction time and number of extractions, which may increase the number of compounds extracted from the plant when optimized. Thus, ASE could have influenced the optimization of extraction and increased the number of flavonoids extracted from ELPA.
The observed cytotoxic activity of ELPA demonstrated in this study might be attributed to the mixture of various flavonoids found in the extract. Flavonoids have the potential to modify many biological cancer events, such as apoptosis, vascularization, cell differentiation and proliferation (Batra and Sharma, 2013). Flavonoids found in ELPA such as vitexin and isovitexin also exert chemotherapeutic potential against breast, hepatic, colorectal, lung, skin, oral, prostate, cervix, ovary, esophagus and leukemic cancers (Ganesan and Xu, 2017). Orientin shows an inhibitory effect on the proliferation of esophageal cancer cells (EC-109) and isoorientin demonstrated the ability to inhibit proliferation of liver cancer cells (HepG2) (An et al., 2015; Xiao et al., 2016; Yuan et al., 2012). Diosmetin, found in ELPA conjugated with C-glucosyl groups (8-C-Glucosyldiosmetin and 6-C-Glucosyldiosmetin), is described as having in vitro anticancer activity against several cancer cells lines, with potent inhibition against leukemic cells (P-388), and lesser cytotoxic activity against HepG2, Hep3B, MDA-MB-231, MCF-7, A549, and Ca9-22 cancer cells (Patel et al., 2013). These data may justify the in vitro cytotoxic effect presented by
ELPA.
The presence of saponins in EFPA may be relevant for its antitumor effect. Saponins exert various pharmacological effects (cardiovascular protective activity, anti-inflammatory, antiviral and immunoregulatory effects) including significant anticancer activity, such as anti-proliferative, anti-metastatic, anti-angiogenic and reverses multiple drug resistance (MDR) effects through mechanisms which include induction of apoptosis and promotion of cell differentiation. In addition to these actions, it is described that saponins may reduce the side effects of radiotherapy and chemotherapy, suggesting that this class of compounds is a promising prospect for anticancer research and development (Man et al., 2010; Xu et al., 2016).
Thus, most of the secondary metabolites identified in the EFPA have proven cytotoxic or anticancer activities, which collaborate with the in vitro cytotoxic action presented as a result of the present work, emphasizing the hypothesis that the action of the EFPA is probably mediated by synergistic activity among its constituents.
For drug discovery, cell death related testing is paramount in oncology because resistance to dying is a hallmark of cancer cells (Méry et al., 2017). Knowing the cytotoxic activity demonstrated by ELPA, we decided to identify the mechanism of the cytotoxicity induced by the extract. For this, HL-60 neoplastic cells were selected for subsequent testing based on the fact that the lineage is among the most widely used cellular models of myeloid origin, and simultaneously was the lineage against which ELPA demonstrated it’s the highest cytotoxic potency. Three concentrations of ELPA, ½ IC50 (9.69 μg/ml), IC50 (19.37 μg/ml) and 2 × IC50 (38.74 μg/ml) were chosen.
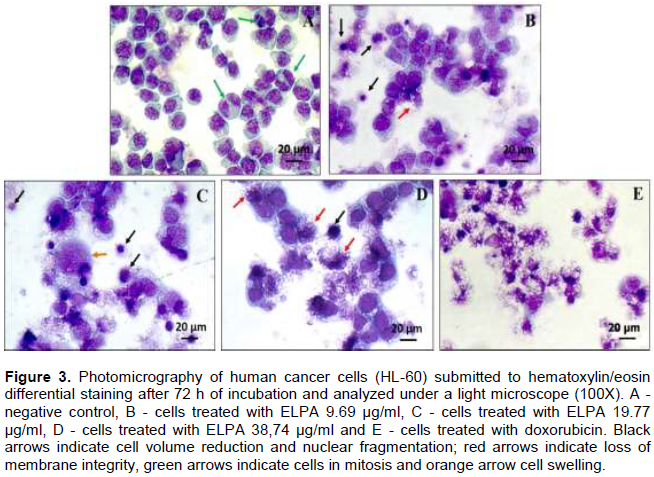
To ascertain the cellular death process in cancer cells, two tests were performed, hematoxylin-eosin coloration analyzed by light microscopy, and ethidium bromide/acridine orange coloration using fluorescence microscopy. After 72 h incubation, the effect of ELPA was evaluated based on cell morphology using hematoxylin-eosin. The HL-60 cells treated with ELPA showed the characteristic morphology of cell death induced by apoptosis (reduction in cell volume, chromatin condensation, and nuclear fragmentation), and necrosis (membrane disruption and cell swelling) as shown in Figure 3.
To confirm the light microscopy findings, we performed a morphological analysis of cells treated with ELPA by staining and examining cells with ethidium bromide/ acridine orange by fluorescence microscopy. The percentages of viable, apoptotic and necrotic cells were calculated. After 72 h, HL-60 cells treated with ELPA showed a reduction in the percentage of viable cells for the concentrations of 9.69 µg/ml (48.82 ± 3.76%), 19.77 µg/ml (41.88 ± 2.58%) and 38.74 µg/ml (34.74 ± 1.25%) when compared with the NC group (91.67 ± 1.72%). The percentage of cells in apoptosis increased for the concentrations of 9.69 µg/ml (30.74 ± 3.22%), 19.77 µg/ml (32.44 ± 3.23%) and 38.74 µg/ml (35.88 ± 3.06%) when compared twith the NC group (4.95 ± 0.98%). Another process of cell death observed was the increase in the percentage of cells in necrosis, for the concentrations of 9.69 µg/ml (20.44 ± 5.86%), 19.77 µg/ml (26.88 ± 5.03%) and 38.74 µg/ml (29.45 ± 3.92%) when compared with the NC group (3.17 ± 1.18%) as shown in Figure 4.
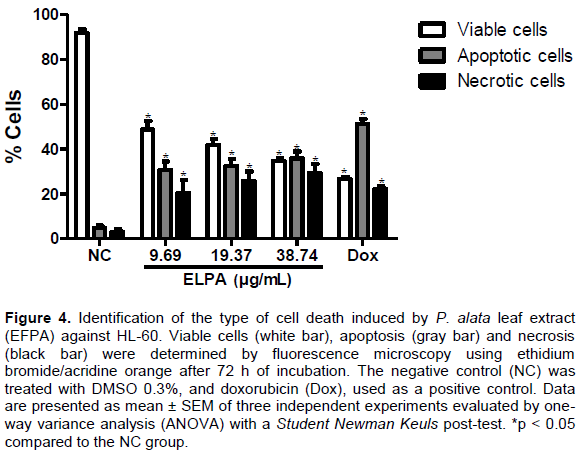
These data reaffirm the results found in the morphological analyses using light microscopy; demon-strating that ELPA can cause cell death by apoptotic and necrotic processes. Apoptosis is a programmed cell death process, and many studies suggest the activation of apoptosis as one of the most effective forms of chemotherapy to prevent the development and progression of cancer (Harvey and Cree, 2010; Yuan et al., 2014). Photodynamic treatment and certain antineoplastic agents, such as β-lapachone and DNA alkylating agents induce cell death by apoptosis and simultaneously induce cell death by necrosis in a variety of cancer cells (Li et al., 1999; Zong and Thompson,2006). Considering the two cell death processes involved, ELPA is a promising candidate for in vivo evaluation in animal experimental tumor models.
The morphological analysis in HL-60 indicated that apoptosis is one of the mechanisms of death involved in the cytotoxicity observed in ELPA, we resolved to determine DNA fragmentation and alteration in the cell cycle through flow cytometry. The results concerning cell cycle progression showed that the ELPA at all concentrations tested showed a significant increase in the percentage of cells in the G2/M phase, being indicative of cell cycle arrest in the specific phase (Table 4).
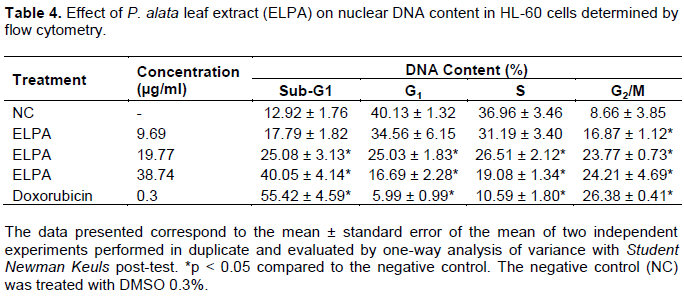
Assessing DNA fragmentation (Sub-G1), the data demonstrated a significant (p < 0.05) increase in fragmentation in the ELPA-treated groups at concentrations of 19.97 and 38.74 μg/ml (Table 4). Thus, cell cycle arrest may be related to ELPA-promoted DNA damage and subsequent DNA repair attempts. The lesion appears to be intense, as the DNA repair does not occur, and apoptosis is triggered.
CONCLUSION
The cytotoxic potential of extracts from 14 Passiflora spp. obtained by ASE was evaluated. ELPA presented greater cytotoxic potential against cancer cells, without cytotoxicity in non-cancer human cells. The in vitro cytotoxic activity may be a consequence of synergistic action between the flavonoids found in ELPA, promoting cell cycle arrest in the G2/M phase and inducing cell death by apoptosis and necrosis. This work thus demonstrates the potential of Brazilian plants and opens perspectives for ELPA evaluation in experimental in vivo tumor models.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
An F, Wang, S, Tian Q, Zhu D (2015). Effects of orientin and vitexin from Trollius chinensis on the growth and apoptosis of esophageal cancer EC-109 cells. Oncology Letters 10(4):2627-2633. |
|
|
Andrade LN, Lima TC, Amaral RG, Do Ó Pessoa C, De Moraes Filho MO, Soares BM, De Sousa DP (2015). Evaluation of the cytotoxicity of structurally correlated p-menthane derivatives. Molecules 20(7):13264-13280. |
|
|
Antoni S, Soerjomataram I, Møller B, Bray F, Ferlay J (2016). An assessment of GLOBOCAN methods for deriving national estimates of cancer incidence. Bulletin of the World Health Organization 94(3):174-184. |
|
|
Batra P, Sharma AK (2013). Anti-cancer potential of flavonoids: recent trends and future perspectives. 3 Biotech 3(6):439-459. |
|
|
Carraz M, Lavergne C, Jullian V, Wright M, Gairin JE, Gonzales De La Cruz M, Bourdy G (2015). Antiproliferative activity and phenotypic modification induced by selected Peruvian medicinal plants on human hepatocellular carcinoma Hep3B cells. Journal of Ethnopharmacology 166:185-199. |
|
|
Chaparro RDC, Maldonado Celis ME, Urango MLA, Rojano IBA (2015). Propiedades quimiopreventivas de Passiflora mollissima (Kunth) L. H. Bailey (curuba larga) contra cáncer colorrectal. Revista Cubana de Plantas Medicinales 20(1):62-74. |
|
|
Chen X, Zhong D, Jiang H, Gu J (1998). Characterization of some glucuronide conjugates by electrospray ion trap mass spectrometry. Yao Xue Xue Bao = Acta Pharmaceutica Sinica 33(11):849-854. |
|
|
Da Silva TBC, Costa COD, Galvão AFC, Bomfim LM, Rodrigues ACB, Mota MCS, Bezerra DP (2016). Cytotoxic potential of selected medicinal plants in northeast Brazil. BMC Complementary and Alternative Medicine 16(1):199. |
|
|
Davis BD, Brodbelt JS (2004). Determination of the glycosylation site of flavonoid monoglucosides by metal complexation and tandem mass spectrometry. Journal of the American Society for Mass Spectrometry 15(9):1287-1299. |
|
|
Dias DA, Urban S, Roessner U (2012). A Historical overview of natural products in drug discovery. Metabolites 2(4):303-336. |
|
|
Ferlay J, Soerjomataram I, Dikshit R, Eser S, Mathers C, Rebelo M, Bray F (2015). Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. International Journal of Cancer 136(5):E359-E386. |
|
|
Ferreira PMP, Farias DF, Viana MP, Souza TM, Vasconcelos IM, Soares BM, Carvalho AFU (2011). Study of the antiproliferative potential of seed extracts from Northeastern Brazilian plants. Anais Da Academia Brasileira de Ciencias 83(3):1045-1058. |
|
|
Ganesan K, Xu B (2017). Molecular targets of vitexin and isovitexin in cancer therapy: A critical review. Annals of the New York Academy of Sciences 1401(1):102-113. |
|
|
Ge C, Li R, Song X, Qin S (2017). Advances in evidence-based cancer adoptive cell therapy. Chinese Clinical Oncology 6(2):18-18. |
|
|
Geng CX, Zeng ZC, Wang JY (2003). Docetaxel inhibits SMMC-7721 human hepatocellular carcinoma cells growth and induces apoptosis. World Journal of Gastroenterology: WJG 9(4):696. |
|
|
Gomes SVF, Portugal LA, Dos Anjos JP, De Jesus ON, De Oliveira EJ, David JP, David JM (2017). Accelerated solvent extraction of phenolic compounds exploiting a Box-Behnken design and quantification of five flavonoids by HPLC-DAD in Passiflora species. Microchemical Journal 132:28-35. |
|
|
Harvey A, Cree I (2010). High-throughput screening of natural products for cancer therapy. Planta Medica 76(11):1080-1086. |
|
|
Harvey AL, Edrada-Ebel R, Quinn RJ (2015). The re-emergence of natural products for drug discovery in the genomics era. Nature Reviews Drug Discovery 14(2):111-129. |
|
|
Hedvat M, Emdad LK, Das S, Kim K, Dasgupta S, Thomas SB, Fisher P (2012). Selected approaches for rational drug design and high throughput screening to identify anti-cancer molecules. Anti-Cancer Agents in Medicinal Chemistry 12(9):1143-1155. |
|
|
Kapadia GJ, Azuine, MA, Tokuda H, Hang E, Mukainaka T, Nishino H, Sridhar R (2002). Inhibitory effect of herbal remedies on 12-O-tetradecanoylphorbol-13-acetate-promoted Epstein-Barr virus early antigen activation. Pharmacological Research 45(3):213-220. |
|
|
Kuete V, Dzotam, JK, Voukeng, IK, Fankam, AG, Efferth T (2016). Cytotoxicity of methanol extracts of Annona muricata, Passiflora edulis and nine other Cameroonian medicinal plants towards multi-factorial drug-resistant cancer cell lines. SpringerPlus 5(1):1666. |
|
|
Li YZ, Li CJ, Pinto AV, Pardee AB (1999). Release of mitochondrial cytochrome C in both apoptosis and necrosis induced by beta-lapachone in human carcinoma cells. Molecular Medicine 5(4):232-239. |
|
|
Ma WD, Zou YP, Wang P, Yao XH, Sun Y, Duan MH, Yu B (2014). Chimaphilin induces apoptosis in human breast cancer MCF-7 cells through a ROS-mediated mitochondrial pathway. Food and Chemical Toxicology 70:1-8. |
|
|
Mahmoud TS, Marques MR, Pessoa C do Ó, Lotufo LVC, Magalhães HIF, Moraes MO, Oliveira JE (2011). In vitro cytotoxic activity of Brazilian Middle West plant extracts. Revista Brasileira de Farmacognosia 21(3):456-464. |
|
|
Méry B, Guy JB, Vallard A, Espenel S, Ardail D, Rodriguez-Lafrasse C, Magné N (2017). In vitro cell death determination for drug discovery: A landscape review of real issues. Journal of Cell Death 10:1179670717691251. |
|
|
Militão GCG, Dantas INF, Pessoa C, Falcão MJC, Silveira ER, Lima MAS, Costa-Lotufo LV (2006). Induction of apoptosis by pterocarpans from Platymiscium floribundum in HL-60 human leukemia cells. Life Sciences 78(20):2409-2417. |
|
|
Mosmann T (1983). Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assay. Journal of Immunological Methods 65(1-2):55-63. |
|
|
Nakayama GR, Caton MC, Nova MP, Parandoosh Z (1997). Assessment of the Alamar Blue assay for cellular growth and viability in vitro. Journal of Immunological Methods 204(2):205-208. |
|
|
Newman DJ, Cragg GM (2016). Natural products as sources of new drugs from 1981 to 2014. Journal of Natural Products 79(3):629-661. |
|
|
Palumbo MO, Kavan P, Miller WH, Panasci L, Assouline S, Johnson N, Batist G (2013). Systemic cancer therapy: Achievements and challenges that lie ahead. Frontiers in Pharmacology 4:1-9. |
|
|
Patel K, Gadewar M, Tahilyani V, Patel DK (2013). A review on pharmacological and analytical aspects of diosmetin: A concise report. Chinese Journal of Integrative Medicine 19(10):792-800. |
|
|
Pearson CH, Cornish K, McMahan CM, Rath DJ, Whalen M (2010). Natural rubber quantification in sunflower using an automated solvent extractor. Industrial Crops and Products 31(3):469-475. |
|
|
Pereira CAM, Yariwake JH, McCullagh M (2005). Distinction of the C-glycosylflavone isomer pairs orientin/isoorientin and vitexin/isovitexin using HPLC-MS exact mass measurement and in-source CID. Phytochemical Analysis: An International Journal of Plant Chemical and Biochemical Techniques 16(5):295-301. |
|
|
Perry NB, Albertson NB, Blunt JW, Cole ALJ, Munro MHG, Walker RL (1991). 4-Hydroxy-2-cyclopentenone: An anti-pseudomonas and Cytotoxic Component from Passiflora tetrandra. Planta Medica 57(4):129-131. |
|
|
Saha S, Walia S, Kundu A, Sharma K, Paul RK (2015). Optimal extraction and fingerprinting of carotenoids by accelerated solvent extraction and liquid chromatography with tandem mass spectrometry. Food Chemistry 177:369-375. |
|
|
Schutz K, Kammerer D, Carle R, Schieber A (2004). Identification and quantification of caffeoylquinic acids and flavonoids from artichoke (Cynara scolymus L.) heads, juice, and pomace by HPLC-DAD-ESI/MS n. Journal of Agricultural and Food Chemistry 52(13):4090-4096. |
|
|
Sujana N, Ramanathan S, Vimala V, Sundaram M, Pemaiah B (2012). Antitumour potential of Passiflora incarnata against ehrlich ascites carcinoma. International Journal of Pharmacy and Pharmaceutical Sciences 4(5):10-13. |
|
|
Veras ML, Bezerra MZB, Braz-Filho R, Pessoa ODL, Montenegro RC, Do Ó Pessoa C, Costa-Lotufo LV (2004). Cytotoxic epimeric withaphysalins from leaves of Acnistus arborescens. Planta Medica 70(6):551-555. |
|
|
Wolfender J, Rodriguez S, Hostettmann K (1998). Liquid chromatography coupled to mass spectrometry and nuclear magnetic resonance spectroscopy for the screening of plant constituents. Journal of Chromatography A 794(1-2):299-316. |
|
|
Xiao J, Capanoglu E, Jassbi AR, Miron A (2016). Advance on the flavonoid C-glycosides and Health Benefits. Critical Reviews in Food Science and Nutrition 56:S29-S45. |
|
|
You JS, Jones PA (2012). Review cancer genetics and epigenetics : Two Sides of the Same Coin ? Cancer Cell 22(1):9-20. |
|
|
Yuan L, Wang J, Xiao H, Xiao C, Wang Y, Liu X (2012). Isoorientin induces apoptosis through mitochondrial dysfunction and inhibition of PI3K/Akt signaling pathway in HepG2 cancer cells. Toxicology and Applied Pharmacology 265(1): 83-92. |
|
|
Yuan L, Wei S, Wang J, Liu X (2014). Isoorientin induces apoptosis and autophagy simultaneously by reactive oxygen species (ROS)-Related p53, PI3K/Akt, JNK, and p38 signaling pathways in HepG2 cancer cells. Journal of Agricultural and Food Chemistry 62(23):5390-5400. |
|
|
Zong W, Thompson CB (2006). Necrotic death as a cell fate. Genes and Development 20:1-15. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


