Herbs have been used for centuries by the world's population for treating a variety of diseases. Among them is
Hypericum perforatum which belongs to
Hypericaceae family
, popularly known as St. John's wort or hiperico (
Greeson et al., 2001;
Linde, 2009). Its main components are: naftodiantronas (hypericin), fluroglucinois (hyperforin), biflavonoids, essential oils and procyanidins (
Barnes et al., 2001;
Mennini and Gobbi, 2004).
The extract of
H. perforatum (EHP), widely distributed throughout the European, Asian and American continents, is used by the populationd ue to its action against depressive disorders that manifest mild to moderate episodes and against anxiety disorders, which become more pronounced during pregnancy. It also has other actions on the central nervous system (CNS), such as antinociceptive and anticonvulsant (
Barnes et al., 2001). The anticonvulsant effect is due to the activation of receptors for opioids and serotonin (5-HT), but mainly through activation of receptors for γ-aminobutyric acid (GABA), primarily located in the hippocampus (
Hosseinzadeh et al., 2005;
Ivetic et al., 2011). The antinociceptive effect results from activation of receptors for GABA, opioids, and 5-HT in both gray periaqueducal area (CPA), and in the spinal cord, besides inhibiting the release of nociceptive mediators associated with inflammation (
Uchida et al., 2008;
Ozdemir et al., 2012).
Studies in the laboratory (
unpublished data) showed that in pregnant Wistar rats at doses of 36, 72 and 144 mg/kg, EHP crossed the placental barrier and the blood-brain barrier and was found (seen through its flurorescence emitted by EHP) both in the mother's organs, as well as in the placenta, brain and liver of fetuses
. The same authors found that male rats born from pregnant rats treated with different doses of EHP (36, 72 and 144 mg/kg) demonstrated behavior in states compatible with antidepressant and anxiolytic responses, when they became adult (90 day old) (unpublished data
). These results suggest that EHP, when reaching the embryo, might be able to change the neurodevelopment of rats, whose maturation occurs several weeks after birth, and thus would be responsible for causing epigenetic modifications in a way that reprograms the CNS still under development (
Murrin et al., 2007).
Since the offspring of treated mothers showed altered antidepressant and anxiolytic responses, this study aimed to verify if the offspring of treated mothers also expressed antinociceptive and anticonvulsant activities different from those expressed in the young of control animals.
Animals and housing
The experiments were performed on Wistar rats (Rattus norvegicus) 90 days old, weighing 200 to 350 g, born to mothers treated with EHP during pregnancy. Males (F1) were used in other experiments. All animals came from the Vivarium of the Center for Reproduction Biology at the Federal University of Juiz de Fora (UFJF) (CIAPE 01.0048.2013). The rats were housed in plastic cages in air-conditioned cabinets, and kept under monitored conditions of temperature of 23±2°C, and dark/light cycle of 12 h. They were fed on rat chow pellets (NuvilabR) and received water ad libitum.
Five animals were placed in each cage. To be transported to the testing room, the rats were individually placed in their cages at least 60 min in advance of the tests to minimize possible behavior alteration. They were deprived of water and food 60 min before the test (
Almeida, 2006). The experimental procedures of this project were approved by the Committee for Ethics in the Use of Animals (CEUA / UFJF): 063/2013.
Treatment with extract of H. perforatum
Extract of H. perforatum
The soluble fraction of the hydro alcohol extract dry containing 0.3% hypericin was prepared by Mbpharma manipulações - Lot 10124778.
Dose calculation: Agência Nacional de Vigilância Sanitária (ANVISA) recommends that the dose of EHP in humans corresponds to the intake of 600 to 900 mg of standardized extract 0.3% hypericin per day (
Rodrigues et al., 2006;
ANVISA, 2008). Based on body surface area, the minimum dose to rats was 36 mg/kg (
Gregoretti et al., 2004).
Experimental groups: Control- pregnant rats received distilled water throughout the gestation period; Treatments- Pregnant rats were grouped into: Treated 1: received 36 mg/kg; Treated 2: 72 mg/kg, and Treated 3: 144 mg/kg of H. perforatum by gavage during all gestation.
Experimental procedure: After weaning (postnatal day 21), the offspring were separated according to sex and then females (F1) were housed in polypropylene cages, measuring 30 x 14 x 16 cm, provided with selected wood shavings for bedding, water and food ad libitum, in number, 5 per cage. The cages were kept on ventilated shelves (AlescoR), provided with control of air flow, temperature and humidity, and located in an environment with control of circadian cycle (12L: 12D) and temperature.
At 90 days of life, the rats were previously selected in diestrus phase, the period with the lowest plasma concentration of estrogen, to minimize the possible hormone interference with antinociceptive responses (
Stoffel et al., 2003). Later, they were euthanized with muscle relaxant xylazine (2% - Kensol
R) associated with the anesthetic ketamine (5% - Vetanarcol
R) (
DBPA;
Council., 2011), via
ip.,followed by diaphragm rupture.
Evaluation of antinociceptive activity
Hot plate test (hot plate)
The rats (F1) of the control and treated groups (T1, T2 and T3) were individually placed on the hot plate (LE 7406 – Panlab) heated to a temperature of 55 ± 1°C. The latency (in seconds), characterized as the time until the appearance of a first response to the thermal stimulus (licking of the paws), was recorded as the index of antinociception for the physical stimulation. The maximum evaluation time is set at 30 s (maximum latency) to minimize damage to the paws of the animals (
Kumar et al., 2001;
Abdel-Salam, 2005;
Almeida, 2006).
The writhing test induced by acetic acid
This is a model of chemical visceral pain induced by injection of glacial acetic acid intraperitoneally (
ip), which is the contraction of the abdominal musculature followed by hip rotation and stretching of the body and hind limbs. The acid acts directly on the fluid of the peritoneum, causing the release of endogenous inflammatory mediators such as prostaglandins E
2 (PGE
2) and I
2 (PGI
2), among others, thereby sensitizing the nerve endings of the region. The (F1) rats of the control and treated groups (T1, T2 and T3) received 10 mL/kg (
ip) 0.8% acetic acid (v/v solution), except the control group which received the same volume of distilled water (10 ml/kg). Five minute after the administration of acetic acid, rats were placed in individual boxes and the number of writhings was evaluated for 15 min. The antinociceptive activity was expressed as the reduction in the number of writhes (
Kumar et al., 2001;
Abdel-Salam, 2005;
Almeida, 2006).
Paw edema test
This consists of nociception induced by chemical stimulation caused by the injection of 50 uL of 2.5% formalin solution (0.92% formaldehyde) under the skin of the plantar surface of the right hind paw (
Almeida, 2006). After formalin injection, the rats (F1) of the control and treated groups (T1, T2 and T3) were placed in glass environments with mirrors on the sides to provide a complete view of animal behavior. Episodes of suspension of the hind paw, which received injection with formalin, were monitored and interpreted as a response to nociception (
Almeida, 2006). Two periods were considered: the first, known as the initial, or first stage, refers to the neurogenic pain stimulation phase, which begins immediately after formalin injection and lasts for 5 min. The second, known as a late stage, or second stage, refers to inflammatory pain and is between 15 and 30 min after the formalin injection. The antinociceptive activity was expressed as the reduction in the time of suspension of the paw. Moreover, the latency to initiate paw suspension was also evaluated; the increase of which was the parameter used to verify the antinociceptive action of the extract (
Almeida, 2006;
Uchida et al., 2008).
Evaluation of anticonvulsant activity
Seizures induced by pentylenetetrazol (PTZ)
PTZ induced seizures by blocking Cl
- channels associated with receptors for GABA
A (
Bukhari et al., 2004;
Hosseinzadeh et al., 2005;
Almeida, 2006). Females (F1) from the control and treated groups (T1, T2 and T3) were placed individually in a cage, where they were given intraperitoneal injection of PTZ (60 mg/kg) and observed for 25 min. The latency time for the onset of seizures, as well as the number and duration of seizures were evaluated.
Convulsion induced by pilocarpine (PLC)
Pilocarpine, a muscarinic agonist, produces motor seizures when administered to mice and rats (300 to 380 mg/kg,
ip) (
Almeida, 2006;
Ngoupaye et al., 2013). (F1) Females from the control and treated groups (T1, T2 and T3) were placed individually in a cage, where they were given intraperitoneal injection of PTZ (60 mg/kg) and observed for 25 min. The latency time for the onset of seizures was evaluated, as well as the number and duration of seizures.
Convulsion induced by auricular electroshock
The auricular electroshock corresponds to repetitive electrical pulses to induce, in different neuronal structures, a characteristic pattern of seizure activity which, when maintained, is called post-discharge (
Almeida, 2006). To evaluate the potential anticonvulsant activity, (F1) rats belonging to the control and treated groups (T1, T2 and T3) were subjected to the auricular shock with a current of 70 mA intensity, at a frequency of 150 pulses/s and duration of 2 s. The following were observed: latency to onset of seizures, number of animals with tonic-clonic seizures, and duration of the convulsive episode (flexion and extension of the legs) (
Almeida, 2006;
Pahuja et al., 2012).
Statistical analysis
The statistical analysis was developed using SPSS version 21. For verification of normality, the Kolmogorov-Smirnov and Shapiro-Wilk tests were used. Parametric data were analyzed by statistical tests ANOVA and T test, followed by post-hoc Tukey, and, for nonparametric data, Chi square test followed by Mann-Whitney. The graphics were made using the Graph Pad Prism program 5.0. For all tests, those ​​that presented significance levels of p<0.05 were considered significant.
Effect of dry extract of H. peforatum on antinociceptive activity
Antinociception assessed through the hot plate test
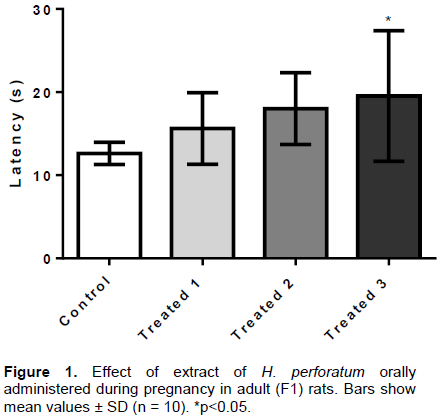
The physical stimulus (high thermal sensation) evaluated by the hot plate test triggers central action neurogenic responses. As shown in Figure 1, the EHP administered to pregnant rats at doses of 36, 72 and 144 mg/kg (T1 groups, T2 e T3, respectively) caused significant increase in latency time in the T3 group (19.5 ± 7.8 s) as compared to the control group (12.6 ± 1.3 s), in the adult F1 rat offspring.
Antinociception assessed by paw edema test
The paw edema test has a biphasic behavior, 2 periods of observation being considered after the administration of formalin. The first stage is characterized by the generation of neurogenic pain, the stimuli of which are primarily transmitted to the CNS by afferent neural fiber types A-δ and C. The second, known as late stage, occurs from the local release of inflammatory mediators such as prostaglandins, histamine, serotonin and bradykinin, and a sharp synaptic activation in the spinal cord and brain via the opioid system, for their control (
Uchida et al., 2008).
The observed results show the central nociceptive effect of EHP on F1 rats. Despite latencies for initiation of paw suspension not statistically different between treatment groups 1, 2 and 3 (Table 1), the rats of the T1 and T2 groups showed a significant reduction in the time for legs to remain suspended (p <0.05) during the initial test phase (phase 1 neurogenic pain) as compared to the controls, which highlights the central antinociceptive action of the extract (Table 1). However, the extract administration did not express such action during the late phase (phase 2 - inflammatory pain) of paw edema test in any of the treated groups as compared to the control (Table 1).
Antinociception evaluated by the writhing test
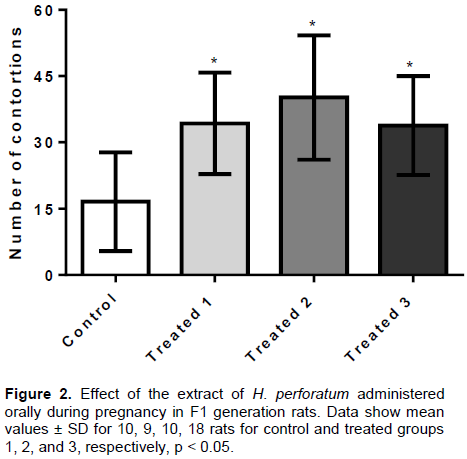
The EHP, administered orally (36, 72 and 144 mg/kg) in pregnant rats, significantly increased the number of writhes induced by acetic acid in adult F1 offspring in the three treated groups (T1: 34.3 ± 11.4 s; T2: 40.1 ± 14.0 s; T3: 33.8 ± 11.1 s) as compared to the control group (16.5 ± 11.1 s) (p <0.01) (Figure 2). These results suggest the occurrence of inflammatory hyperalgesia activity of the extract on the F1 generation rats as adults..
Effect of dry extract of H. peforatum on the anticonvulsant activity
Evaluation by pentilenotetrazol test (PTZ)
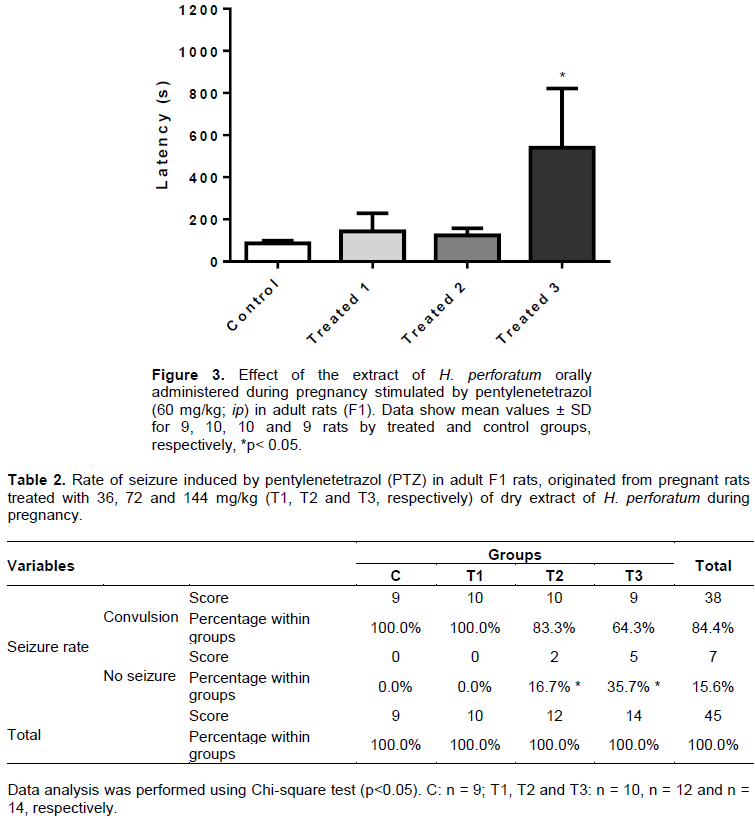
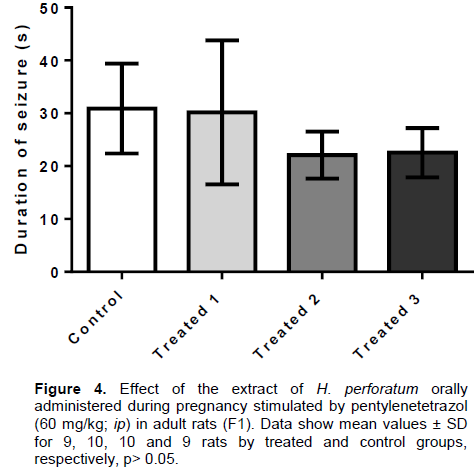
The pentylenetetrazol (PTZ) is a convulsant agent which acts by inhibiting the activation of GABA receptors in the CNS, besides reducing nitric oxide production. The EHP, administered orally (36, 72 and 144 mg/kg) in pregnant rats caused increased anticonvulsant activity in adult F1 rats by increasing the latency to onset of seizures in the group treated with the highest dose (T3: 540.2 ± 281.5 s) in the control group (86.3 ± 27.2 s) (Figure 3), and also reduced the seizure rate of T2 groups (16.7%) and T3 (35.7%), as compared to the control (Table 2). However, the duration of convulsive episodes were not statistically different between the treated groups (T1: 30.2 ± 13.6 s; T2: 22.1 ± 4.4 s; T3: 22.5 ± 4.6 s) and control (30.9 ± 8.5 s), despite the tendency of the T2 and T3 groups to present reduction of this parameter as compared to control rats (Figure 4). No animal deaths were observed in the control and treated groups.
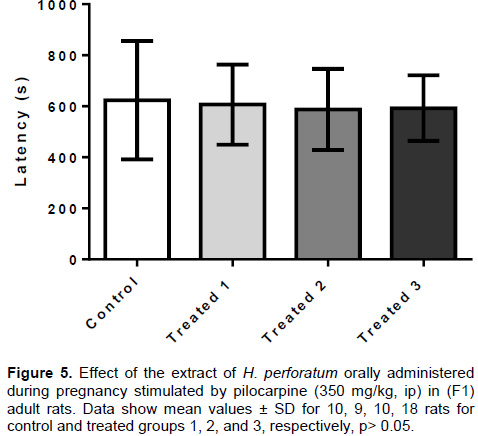
Evaluation by testing with pilocarpine (PLC)
The EHP, administered orally (36, 72 and 144 mg/kg) in pregnant rats during pregnancy, showed no anticonvulsant effect in F1 adult rats after induction of seizures by pilocarpine. The latencies (Figure 5)observed in treated groups T1 (606.6 ± 157.1 s), T2 (588.0 ± 159.4 s) and T3 (593.3 ± 128.3 s) did not differ significantly from the control group (624.0 ± 231.8s). Since all animals in the control and treated groups (except one animal of the group T1) expressed seizures induced by pilocarpine, the seizure protection rate was not evaluated.
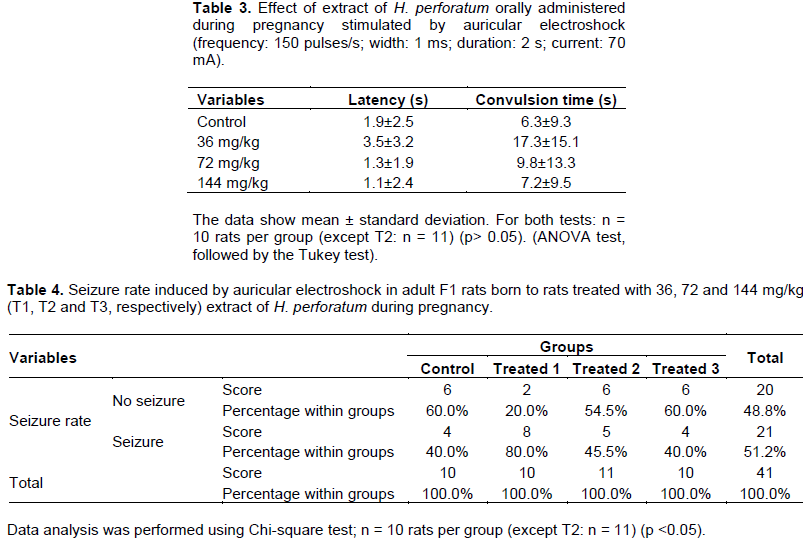
Evaluation by electroshock auricular test
EHP, orally administered in doses of 36, 72 and 144 mg/kg in pregnant rats during pregnancy, showed no anticonvulsant activity in adult F1 rats in relation to the control group, after induction by auricular electroshock. The parameters evaluated: latency (Table 3); seizure time (Table 3 and seizure rate (Table 4) did not differ significantly between the control and treated groups. No animal deaths were observed.
In this work, it was shown that the antinociceptive and anticonvulsant actions of EHP were enhanced in adult F1 rats originating from mothers treated with different doses (36, 72 and 144 mg/kg) throughout pregnancy. The antinociceptive activity of the EHP is promoted by activation of opioid receptors (
Galeotti et al., 2010)and GABAergic receptors (
Barnes et al., 2001) in the CNS (brain and spinal cord) and also by inhibiting the reuptake of 5-HT noradrenaline and dopamine (
Müller, 2003), while the anticonvulsant action results mainly from the activation of GABA receptors in the hippocampus (
Ivetic et al., 2011).
Anxiolytic and antidepressant effects of EHP in the F1 generation coming from mothers treated during pregnancy, as well as the absence of neuromotor and memory changes in neonatal rats were described by Campos and coworkers (unpublished data
). The assessment of the effects of EHP in the F1 generation is important for verifying toxicity and aims to contribute to the safe use of this herbal medicine for pregnant women, because its physiological action through the activation of CNS receptors opioids (
Schuurmans et al., 2015), serotonergic drugs (
Kroeze et al., 2016), noradrenergic drugs (
Murrin et al., 2007)and GABAergic drugs (
Xia and Haddad, 1992), may interfere with neurodevelopment (
Kroeze et al., 2016). This action can occur by different mechanisms and pathways, through the control of apoptosis, mitosis, cell differentiation, dendritogenesis and synaptogenesis in different stages of life and places, but especially in areas related to pain control and seizures (
Brunton et al., 2012).
Specifically,
H. perforatum has proven antinociceptive action from the activation of opioid receptors by its component hyperforin (
Bukhari et al., 2004;
Uchida et al., 2008)and the participation of hypericin (
Galeotti et al., 2010a)through dephosphorylation of protein kinase C (PKC), which has algesic action when phosphorylated. The analgesic effect of EHP reaches its maximum point 120 min after administration and tends to decrease thereafter (
Galeotti et al., 2010a). Vieira et al. (2013) demonstrated that EHP promoted reduction of states of anxiety and depression in rats treated throughout pregnancy, with effects that lasted up to 60 days after the end of treatment (
Vieira et al., 2013). In this case, it is believed that the maintenance of anti-nociceptive effects, and anticonvulsants, from the birth of rats born to treated mothers, is a result of cellular changes that overlap the pharmacokinetic effects of extract components, since these modifications persist into adulthood.
Hyperforin has antinociceptive action via receptor activation
mi, kappa and delta capable of hyperpolarize neuronal membranes, and also by reduction of PKC expression (γ and ε) the APG (
Galeotti et al., 2010a). During pregnancy, there is production of opioid receptors in the brain and spinal cord of rats (
Bernard et al., 1990)whose expression shows variations (
Van Praag and Frenk, 1991), an increased number occurring of κ receptors (
Barr et al., 1986), and μ receptors (
Auguy-Valette et al., 1978)until about 15 days post-natal.
It is proposed in this paper that the frequent availability of hyperforin during embryonic development has caused tolerance (down regulation) (
He et al., 2002)in the opioid receptors until the termination of pregnancy, reprogramming the brain areas related to pain control (APG and brainstem) and sensitive to this neurohormone, to a state with fewer receptors. However, with the end of treatment with EHP, it is assumed that there has been upregulation as a rebound effect so as to increase the number of receptors in F1 rats, above normal levels of brain growth for this period of post-natal neurogenesis (
Bouza, 2009), reprogramming these regions to a new state of normality. As the changes caused by EHP are lasting (
Vieira et al., 2013)and plasma concentrations of enkephalins are not altered by treatment with this extract (
Uchida et al., 2008), it is believed that the functional response of the receptor has become more marked, increasing the antinociceptive response in the offspring of mothers treated with EHP as compared to the control group young.
The continued use of hyperforin leads to decreased PKC expression in the APG, while hypericin is able to dephosphorylate PKC, inactivating it. Thus, it decreases the expression of transcription factors including Signal Transducer and Activator of Transcription (STAT-1), NF- and Cyclic AMP response element binding protein (CREB), involved in the activation of intracellular pathways in the APG and thalamus of mice (
Galeotti et al., 2010b;
Galeotti and Ghelardini, 2013). These mechanisms explain the synergistic interaction of hyperforin and hypericin present in the EHP to promote the antinociceptive response which is lasting and maintained into the adulthood of rats analyzed in this study.
Both 5-HT and norepinephrine are neurotransmitters directly involved in nociceptive response and neuro-development. Serotonin is one of the first neurotransmitters synthesized (
Gaspar et al., 2003)during embryogenesis, starting the second week of gestation and its production is maintained to the postnatal period. Norepinephrine is synthesized later, but is also involved in the development of the CNS (
Murrin et al., 2007). However, the activation maintained of these neurotransmitters generates tolerance (down regulation) (
Stahl, 1998;
Liang et al., 2007), which compromises this action. Since EHP increases the reuptake of serotonin and norepinephrine, and has been given throughout pregnancy, it is proposed that it acts to reprogram the functioning of the CNS, particularly of the brain stem, so as to increase the pain threshold. This would happen through the manifestation of tolerance mechanisms (prenatal down regulation followed by postnatal upregulation), which enhances the antinociceptive response when activated in adults. Studies have shown that there is a synergistic analgesic effect between morphine and serotonin and norepinephrine reuptake inhibitors (SNRI), when administered concurrently, besides the decrease occurring in morphine tolerance (
Ozdemir et al., 2012). Furthermore, α
2-noradrenergic receptors are located side by side in the cell membranes with μ receptors (MOR) in peripheral afferent nerves, which increases the chance of involvement of both in the response to treatment with EHP, by sharing some intracellular signaling pathways (activation of Gq protein - via protein kinase C - PKC - as second messenger) at this location (
Aley and Levine, 1997).
GABA also raises the threshold for triggering pain stimulus through hyperpolarization of the cell due to the opening of Cl
- channels (with glycine), mainly in the dorsal horn of the spinal cord (laminae I and II), hippocampus, thalamus, but not in the cerebral cortex (
Kharkevich and Churukanov, 1999;
Bonin and De Koninck, 2013). Some studies have shown that this action is stimulated by EHP, occurring mainly via the GABA
A receptor (
Turkmen et al., 2011). Following the reasoning above, from the prior knowledge that there is an increase of GABAergic fibers in rats during the first 2 weeks postnatal (
Xia and Haddad, 1992). It is believed that the activation of the GABA
A receptor maintained by EHP administered during pregnancy develops tolerance (down regulation) followed by upregulation postnatal in the regions mentioned above, which consequently reprograms fetal nerve cells to produce more GABA receptors. This would result in increased sensitivity in the expression of its natural effect, that is, decrease in pain (
Ito et al., 1996).
Thus, the administration of the EHP may have caused a reprogramming of neuronal cells in the CNS regions related to antinociceptive activity (hippocampus, APG, raphe nucleus, posterior horn of the spinal cord), which led the body to have reduced action to control pain and therefore to be less responsive to the detection of physiological and pathological changes. This would result in increased risk to the health and maintenance of homeostasis, featuring the neurotoxic action of EHP on the F1 generation (
He et al., 2002;
Nagi, 2011).
Nociceptive stimuli related to inflammation are generally associated with tissue damage resulting both from the painful agent, and the immune response against it. Their maintenance, then, is important for detecting changes in homeostasis. They involve the local release of autacoids such as prostaglandins, leukotrienes, 5-HT and others, which stimulate afferent fibers conducting nerve impulses (pain) to the CNS (
Brunton et al., 2012). Thereafter, the activation of the antinociceptive pathway occurs by the opioid system, serotoninergic system and others discussed above.
F1 rats born to mothers exposed to EHP showed greater sensitivity to the painful inflammatory agent acetic acid, as noted in the test writhes, though this extract can decrease the production of inflammatory mediators (
Albert et al., 2002;
Perazzo et al., 2008). As previously discussed, exposure of the F1 animals to EHP during neurogenesis caused increased central antinociceptive response but it is proposed that this could not prevent the generation and propagation of nociceptive stimuli to the central nervous system. Another possibility could be the enhancement of expression of nociceptive inflammatory mediators, also resulting from reprogramming the peripheral nerves, capable of generating pain stimulus greater than the responses for the control. However, this proposition needs to be proved from other studies.
The anticonvulsant action of EHP is well characterized in the literature and involves the activation of the receptors of the opioid system, the serotonergic system, but mostly the GABAergic system (GABA
A, via nitric oxide production) (Hosseinzadeh et al., 2005; Ivetic et al., 2011). The results of this study show that adult rats (F1) born to mothers treated with EHP have fewer seizures than untreated rats, when stimulated with PTZ, an antagonist of the GABA neurotransmitter. Therefore, it is assumed that neuronal reprogramming (down regulation prenatal then upregulation post-natal), which occurred during embryonic development, was responsible for alterations in neuron receptors located both in regions related to antinociceptive activity (discussed above), and in regions involved with anti-convulsive episodes (cortex and hippocampus), since the hippocampus presents GABAergic fibers from the beginning of neurogenesis, besides the increased number of GABAergic synapses during the first two postnatal weeks (
Swann et al., 1989).
The seizures induced by pilocarpine occur by its action on different receptors in the hippocampus, the striatum and frontal cortex: muscarinic-M
1 (mostly), 5-HT, NMDA and GABA (
Freitas et al., 2004). Studies relating seizures to the use of EHP are scarce in the literature. The results of this study allow the supposition that the action of the extract on such receptors in the CNS of F1 rats during embryonic development was not enough to prevent the triggering and maintenance of potential seizures, since the maturation of the cholinergic system occurs late in relation to opioid, serotonergic, noradrenergic and gabaergic systems (to the third postnatal week) (
Cavalheiro et al., 1987), besides having little affinity with the EHP.
The auricular electroshock promotes the generation of seizures by physical stimuli that facilitate reaching the threshold for triggering the convulsive potential (
Hosseinzadeh et al., 2005). It is believed that this stimulus is stronger than the biochemical stimuli from release of neurotransmitters for the generation of seizures because the originating neural reprogramming changes, through which the (F1) rats having contact with EHP passed, were not enough to promote an effective anticonvulsant response, as found in the results of this work.
In conclusion, the data obtained in this study suggest that treatment of pregnant rats with EHP interferes with neurodevelopment and neurofunction of F1 rats, reprogramming brain areas related to pain control and seizures, making them less responsive to their stimuli, probably due to the increase in opioid, serotonergic, noradrenergic and GABAergic receptors to above normal levels (upregulation postnatal). The treatment caused long lasting effects on the offspring, considering that tests were performed on adult female rats.