Full Length Research Paper

ABSTRACT
The present paper aimed to characterize the bioactivity being exerted on Sclerotium rolfsii Sacc. in vitro by seed aqueous extracts from 64 yam bean (Pachyrhizus spp.) genotypes as well as their maceration times (0, 48 and 72 h). Three experiments were carried out following a completely randomized design with 64 treatments (potato-dextrose-agar PDA+extracts) with two controls (PDA with no extract and PDA+Cabrio® Top in the dose of 4 g/L) with four replicates with one Petri dish per experimental unit. The 64 treatments were adjusted in the dose of 0.1%. The mycelial growth diameter was assessed on the fifth day. Findings showed the diameters of macerating extracts to be 34.3 - 67.0, 23.5 - 57.8, and 37.5 - 63.8 mm at 0, 48, and 72 h, respectively. PDA+Cabrio® Top and PDA with no extract controls ranged from 0.05 to 0.50 mm and 73.2 to 80.7 mm, respectively. P14, P1, P15, and P46 genotypes extracts achieved the lowest mean diameters (47 - 48% shorter). P61 was the genotype bearing the largest mean diameter tantamount to a reduction of 21%. Therefore, every aqueous extract has shown to be active against S. rolfsii, which indicates that genotypes exerting higher toxicity should either be tested against other pathogens or used in the field macerating them for 48 h.
Key words: Jacatupé, yam beam, secondary metabolites, toxicity, natural fungicides.
INTRODUCTION
The search for biodegradable or less harmful to environment pesticides has increased in the past years. Hence, Brazil, as a signatory of international treaties such as Kyoto, COP 21 Paris agreement, and the United Nations 2030 agenda has found that a lot of research should be developed concerning this subject.
The Brazilian plant species richness still needs to be further investigated to ascertain what each one of them might be used for. A large number of plants bearing bactericidal, fungicidal, and insecticidal properties are being used as traditional remedies (Lorenzi and Matos, 2008). But, to start this kind of endeavor in Amazonia one must search for plants the agronomical management of which is well developed and have shown to be effective for controlling pathogens. Thus, Pachyrhizus spp. Fabaceae, named jacatupé or feijão-macuco in Brazil; jícama in Central America and Mexico; and jíquima, asipa and ulillo in Peru (Valcárcel, 1984) has stood out, under this context. This genus holds three cultivated (Pachyrhizus tuberosus, Pachyrhizus erosus, and Pachyrhizus ahipa) and two wild (Pachyrhizus ferrugineus and Pachyrhizus panamensis) species (Bastidas, 1998). These plants are characterized by bearing toxic seeds, unfit for human (Yu et al., 2020) or animal feeding, the main metabolite of which is rotenone (Estrella-Parra et al., 2014; Upegui et al., 2014; Catteau et al., 2013) whose concentration ranges from 0.11 to 0.28% in dry matter (Lautie et al., 2012). This substance inhibits the mitochondrial respiratory chain by blocking the production of adenosine triphosphate (ATP), leading to cell death (Catteau et al., 2013).
The Sclerotium rolfsii Sacc. fungus is quite common in Amazonian soils and it mainly attacks the cultivation of Solanaceae such as tomato, sweet pepper, and cocona (Laborda et al., 2019; Soares et al., 2017). As a result, the use of yam bean seed extracts might help to control this pathogen. Preliminary in vitro tests have determined 0.1% aqueous extracts to be effective for controlling S. rolfsii (Martins and Ticona-Benavente, 2017).
The Germplasm Bank of the National, Amazonian Research Institute (INPA) preserves 64 genotypes of this bean, which are yet to be characterized as to their bioactivity against many pathogens and pests. The present work has aimed to characterize the bioactivity that 64 yam bean aqueous extracts exert on S. rolfsii in vitro; and from them select genotypes bearing the greatest effect for the control of this fungus, to be later used as a biodegradable fungicide.
MATERIALS AND METHODS
The experiments were conducted at the Phytopathology laboratory of the National, Amazonian Research Institute (INPA).
Preparation of the extracts
The extracts were prepared from seeds of 64 yam bean genotypes being preserved at the INPA’s germplasm bank. Seeds were oven-dehydrated at 50°C for 24 h, then crushed to a fine flour using cutting mill. The aqueous extract was then prepared by placing 0.1 g of the flour in 10 ml of distilled water (=1% mass/volume).
Obtaining of S. rolfsii
S. rolfsii sclerotia were obtained by hand, in the field, from infected plants of cocona (Solanum sessiflorum Dunal) at INPA’s ‘Alejo von der Pahlen’ experimental station (2°59’48’’S, 60°1’22’’W, altitude 60 m). They were disinfected in the laboratory by being dipped into 70% alcohol for 1/2 min and 2% sodium hypochlorite for 2 min (Personal communication by Luiz Alberto G. Assis). They were then washed twice with sterile distilled water and dried on filter paper. Sclerotia were multiplied by being placed in a PDA culture medium for 10 days at room temperature (~25°C) and permanent light (450 lumens).
Experimental design
Three experiments were carried out with different maceration times of yam bean flour in the water at 4°C: 0 (control), 48 and, 72 h. Following these times every extract was mixed with PDA adjusting to a final concentration of 0.1%. This solution was autoclaved for 20 min, ~117°C, and 0,1 MPa and placed on 9 cm-diameter Petri dishes within a laminar flow cabinet. Each experiment prepared 256 Petri dishes with 64 extracts + PDA plus 64 Petri dishes with the positive control (PDA + Cabrio® Top) and 64 with negative control (PDA) following a completely randomized design with 64 treatments, four replicates, with one Petri dish per experimental unit plus two additional treatments (controls).
Antifungal evaluation
Disinfected sclerotia were aseptically placed in the center of the Petri dish within a laminar flow cabinet. Then, the Petri dishes were sealed with plastic film and set at room temperature (~25°C). Following five days, the mycelial growth diameter of the two orthogonal axes was evaluated and its mean calculated. The percent reduction of mycelial growth was estimated using the formula (1-MDGEN/MDPDA)*100, where MDGEN= Mycelial diameter in genotype extracts + PDA, and MDPDA= mycelial diameter in PDA.
Statistical analyses
Data were subjected to individual variance analysis and means compared by the Scott-Knott test at 5% probability using the SISVAR 5.0 program (Ferreira, Universidade Federal de Lavras, MG). With the diameter averages of each maceration time a biplot, based on the principal component analysis, was constructed from the correlation matrix. JMP 10.0 (SAS Institute, Cary, NC) was the software being used. To detect the significance of treatments, maceration times, and treatments x maceration times (T x MT) effects, the joint variance analysis was also performed considering treatments and maceration times as fixed effects. In the case of T x MT is significant its decomposition was performed to identify extracts stability across maceration time. These analyses were made by using the SISVAR 5.0 (Ferreira, D.F., Federal University of Lavras-MG).
RESULTS AND DISCUSSION
Testing plant extracts bioactivity on pathogens, has become paramount for biodegradable pesticides development. As to Pachyrhizus spp. its toxic effect on fungi has been observed as well (Martins and Ticona-Benavente, 2017; Barrera-Necha et al., 2004). This points out that one is liable to find other genotypes that exert even greater toxicity on S. rolfsii. Joint variance analysis showed treatments, times (0, 48, and 72 h) and treatments x times are significant effects have modified the mycelial growth (Table 1).
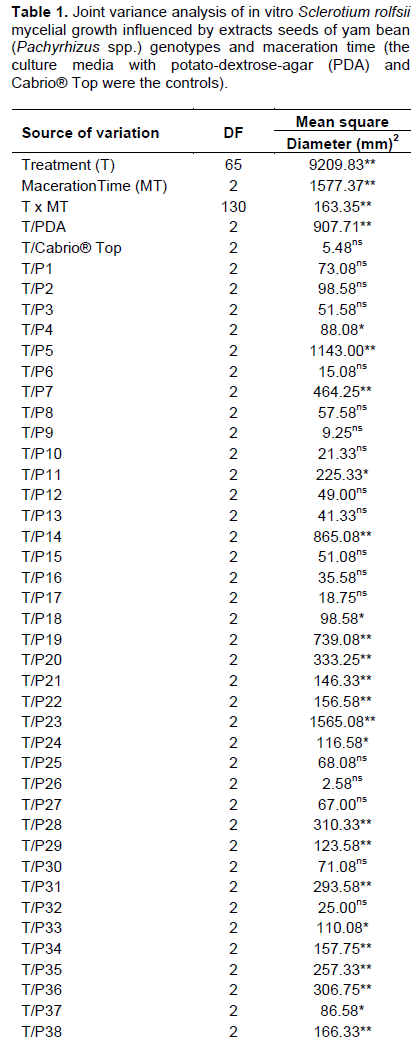
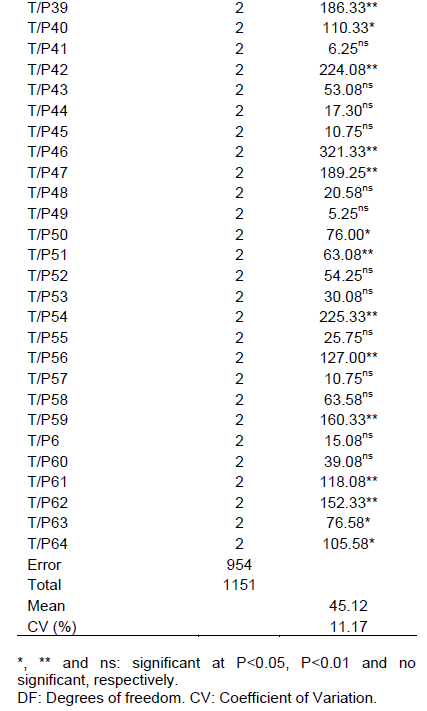
T x MT decomposition demonstrated that nearly half of the genotypes presented no significant variance between maceration times (Table 1). This indicates them to be stable up to 72 h at 4°C. Highlighting genotypes P1 (41.3 mm) and P15 (40.6 mm), presenting below average diameters (45.12 mm). Other tests, taking longer storage and maceration times at room temperature into account, will be needed to ease the use of these extracts out in the field.
On the other hand, genotypes P23 and P5 showed to be the ones contributing the most to interaction, since they presented the highest mean squares (Table 1). This means that maceration time has influenced their bioactivity. The extracts mycelial growth mean obtained with 0 h of maceration for the control treatments was 0.06 mm for PDA+Cabrio®Top and 76.84 mm for PDA with no extracts, and PDA+extracts it ranged from 34.3 mm (P46) to 67.0 mm (P19) (Table 2). This is equal to it being reduced by 12-55%. Highlighting genotypes P46, P20, and P39, whose diameters showed less than 38.50 mm (a reduction of between 50 and 55%). P19, P61, and P28 genotypes showed to be the ones bearing high mycelial growth ranging from 63.0 to 67.0 mm (reduction between 12 and 18%). These findings point out that P46, P20, and P39 genotypes aqueous extracts can be tested in the field for the control of S. rolfsii since they need no time to be macerated.
When comparing mycelial growth means at 48 h, one observes controls to hold diameters of 73.2 and 0.6 mm for PDA and Cabrio® Top respectively (Table 2), though extracts ranged from 23.5 mm (P23) to 57.8 mm (P31) (Table 2). This points out that extracts bear a toxic effect on S. rolfsii, but not on the same level as that being exerted by Cabrio® Top. Considering PDA as the maximum growth, one may infer that extracts reduce the diameter by 21.0 to 67.9%. Highlighting P23, P5, and P14 genotypes the diameters of which failed to reach 25 mm. This demonstrated these genotypes to control S. rolfsii more efficiently. P31 (57.8 mm), P34 (56.3 mm), and P28 (56.0 mm) showed to be the ones where the fungus presented the largest mycelial growth.

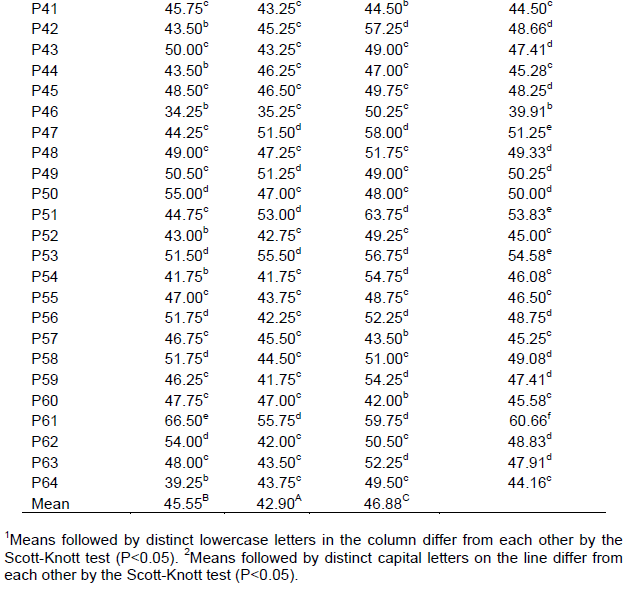
Controls held diameters from 0.05 to 80.7 mm for Cabrio® Top and PDA, respectively, at 72 h of maceration. Yet, extracts held diameters ranging from 37.5 mm (P35) to 63.8 mm (P51). This is tantamount to a 21-54%. P35 (37.5 mm); P25 (40.3 mm) and P21 (40.5 mm) showed to be the genotypes that did control the fungus efficiently. As a result, these genotypes are recommended to be used with 72 h of maceration. Conversely, P51, P61, and P40, hold diameters of between 58.8 and 63.8 mm, bore lower efficiency for controlling the above-mentioned fungus.
Other plant species have been used for controlling S. rolfsii. For instance: i) aqueous extracts from crajiru (Arrabidaea chica) and hot pepper (Kamila et al., 2009), both in the dose of 20%, inhibited injury on the bottom stem of the tomato (Kamila et al. 2009); ii) aqueous extracts from nettle (Urtica sp.) at 10, 20 and 50% concentrations were also effective in vitro, reducing mycelial growth by 90% (Soares, 2013); iii) aqueous extracts from rosemary (Helianthus annuus L.) at 15 and 20% concentrations reduced mycelial growth by 100% (Araújo et al., 2014). Nevertheless, yam bean seed extracts were at a concentration of 0.1% and reduced the mycelial growth by up to 67%, which indicates that extracts from this bean might bear greater toxicity against this fungus than that being exerted by crajirú, pepper, nettle, and rosemary.
Comparing 0, 48, and 72 h as the average time for all treatments (Table 2), one observes mycelial growth to be significantly lower at 48 h than at the other maceration times (42.9 mm) and higher (46.8 mm) at 72 h. Thus, this has demonstrated 48 h to be the best time for preparing aqueous extracts.
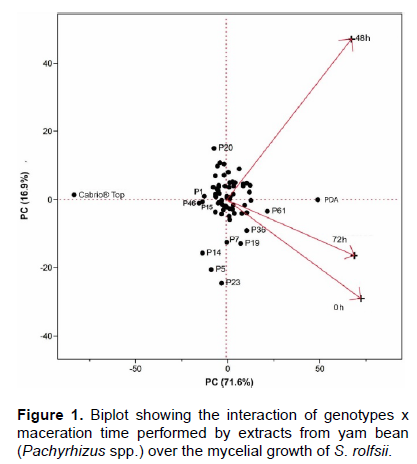
Based on a variance and covariance matrix regarding the genotypes’ mycelial growth means, at the three maceration times, a biplot that accounted for 88.5% of the total variation, was constructed (Figure 1), increasing the reliability of the geometrical interpretations. Biplot graph demonstrates that 48 and 72 h, and 48 and 0 h vectors tend to form a 90° angle pointing out 48 h mycelial growth not to be associated with that at 0 and 72 h. It also shows 0 and 72 h to be associated. These findings indicate toxicity to rise along with maceration time, up to 48 h, and to start coming down afterward. Regarding treatments distribution, one has noted extracts to have had a behavior intermediate to the performances played by Cabrio® Top and PDA with no extracts. When just taking extracts into account one observes their variability when controlling S. rolfsii. However, this variability concentrated around the origin of the vectors, indicates their performance to tend to the overall average. P23, P5, and P14 genotypes stood out due to their fungicidal effect at 48 h, confirmed by the 23.5; 23.8, and 24.5 mm mycelial growth means, respectively. Other promising genotypes are the ones geometrically closer to Cabrio® Top. Such as P46, P15, and P1, presenting 39.9; 40.6, and 41.3 mm overall means, respectively. P20 (35.5 mm) stands out when considering the aqueous extracts prompt use. Based on these findings P23, P5, P14, P46, P15, P1, and P20 genotypes might be recommended to either conduct novel toxicity tests with other pathogens or arthropods. The aqueous extracts showed to be promising for controlling S. rolfsii. However, other solvents should be tested when preparing them so as to enhance their efficiency.
On the other hand, the main active ingredient of these extracts is rotenone, which is water-insoluble (Sigma, 2019). Therefore, other studies should quantify the extracts’ rotenone contents in order to ascertain whether this metabolite would be associated with this fungus growth inhibition. Studies of rotenone decomposition by sunlight and ultraviolet irradiation showed rotenone to have a half-life of 10 min to 2.5 days (Cabizza et al., 2004; Chen et al., 2009). Meanwhile, rotenone’s lowest lethal dose in children via oral was 143 mg/kg (WHO, 1992). As rotenone concentration in Pachyrhizus seeds ranges from 0.11 to 0.28% in dry matter (Lautié et al., 2012), one child would need to eat at least 51 g/kg to die, although, there is one accidental intoxication case where the person died after eating 80 g of Pachyrhizus seeds (Fu and Wang, 2012). For this reason, World Health Organization considers rotenone as moderately toxic for humans (WHO, 1992). Therefore, Pachyrhizus extracts have a low impact on the environment and humans.
As Pachyrhizus extracts showed intermediary efficiency when compared with two controls, they could be used in agriculture to control S. rolfsii with minor impact over some pollinators such as bees (Xavier et al., 2010) and soil mesofauna (Guimarães et al., 2019). Other studies must be carried out to estimate the right occasion and optimal frequency of extracts application for each crop.
In order to produce pesticides based on this bean, it will be necessary to evaluate the P1, P5, P14, P15, P23, and P46 genotypes grain yield potential, as well as quantify their rotenone content. Then try to adapt the high-performance industrial extraction with supercritical CO2 used in Derris elliptica (Baldino et al., 2018), which showed to have an efficiency of 93% for extracting rotenoids.
CONCLUSION
Yam bean seed aqueous extracts should be used at 0.1% concentration, macerated for 48 h at 4°C to increase their efficiency in controlling S. rolfsii. P1, P5, P14, P15, P20, P23, P46, and, P64 genotypes should be investigated in the field to determine their agronomical performance and grain yield. Other solvents must be tested to evaluate extracts efficiency.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENT
The authors appreciate the Coordination for the Improvement of Higher Education Personnel - CAPES by Master’s scholarship. All authors thank FAPEAM (process N° 062.00855/2019) for funding the publication of this paper.
REFERENCES
|
Araújo JL, Oliveira ES, Teixeira FN (2014). Controle alternativo in vitro de Sclerotium rolfsii em girassol (Helianthus annuus L.) pelo uso de extratos vegetais e Trichoderma ssp. Essentia 15(2):25-35. |
|
|
Baldino L, Mariarosa S, Reverchon E (2018). Extraction of rotenoids from Derris elliptica using supercritical CO2. Journal of Chemical Technology and Biotechnology 93(12):3656-3660. |
|
|
Barrera-Necha LL, Bautista-Banos S, Bravo-Luna L, García-Suárez FJL, Alavez-Solano D, Reyes-Chilpa LL (2004). Antifungal activity of seed powders, extracts, and secondary metabolites of Pachyrhizus erosus L. against three postharvest fungi. Mexican Journal of Phytopathology 22(3):356-361. |
|
|
Bastidas CGT (1998). Caracterización morfológica y molecular de la diversidad genética de la colección de Pachyrhizus tuberosus (Lam.) Spreng. del Catie. Master Thesis, Centro Agronómico Tropical de Investigación y Enseñanza. |
|
|
Cabizza M, Angioni A, Melis M, Cabras M, Tuberoso CV, Cabras P (2004). Rotenone and rotenoids in cube resins, formulations, and residues on olives. Journal of Agricultural and Food Chemistry 52(2):288-293. |
|
|
Catteau L, Lautie E, Kone O, Coppee M, Hell K, Pomalegni CB, Quetin-Leclercq J (2013) Degradation of rotenone in yam bean seeds (Pachyrhizus sp.) through Food Processing. Journal of Agricultural and Food Chemistry 61(46):11173-11179. |
|
|
Chen XJ, Xu HH, Yang W, Liu SZ (2009). Research on the effect of photoprotectants on photostabilization of rotenone. Journal of Photochemistry and Photobiology B: Biology 95(2):93-100. |
|
|
Estrella-Parra EA, Gomez-Verjan JC, Gonzalez-Sanchez I, Vázquez-Martínez ER, Vergara-Castañeda E, Cerbón MA, Alavez-Solano D, Reyes-Chilpa R (2014). Rotenone isolated from Pachyrhizus erosus displays cytotoxicity and genotoxicity in K562 cells. Natural Products Research 28(20):1780-1785. |
|
|
Fu PK, Wang PY (2012) Toxic leukoencephalopathy due to yam bean seeds poisoning. The Neurologist 18(4):199-201. |
|
|
Guimarães RGV, Cevalho WC, Ticona Benavente CA, Pinto FR, Alencar DB (2019). Effect of Cabrio® Top and macuco bean (Pachyrhizus spp.) extracts on soil mesofauna. International Journal of Advanced Engineering Research and Science 6(12):343-347. |
|
|
Kamila FA, Santana Dezordi C, Coelho Netto RA, Hanada RE (2009). Avaliação do controle da podridão de Sclerotium rolfisii. em tomateiro (Solanum lycopersicum) por meio do uso de extratos de planta. INPA. |
|
|
Laborda PP, Nina AC, Silva Filho DF, Coelho Netto RA (2019). Incorporação de biomassa de fabáceas ao substrato de plantio para controle da podridão-de-escleródio (Sclerotium rolfsii) em cubiu (Solanum sessiliflorum). Summa Phytopathologica 45(4):399-405. |
|
|
Lautie E, Rozet E, Hubert P, Leclercq JQ (2012). Quantification of rotenone in seeds of different species of yam bean (Pachyrhizus sp.) by a SPE HPLC-UV method. Food Chemistry 131(4):1531-1538. |
|
|
Lorenzi H, Matos FJA (2008). Plantas medicinais no Brasil: nativas e exóticas. Nova Odessa, São Paulo: Instituto Plantarum. |
|
|
Martins AKB, Ticona-Benavente CA (2017). Extratos de feijão-macuco (Pachyrhizus spp.) controlam Sclerotium rolfsii in vitro. In Anais do VI congresso de iniciação cientifica do INPA - CONIC ISSN 2178 9665. |
|
|
SIGMA (2019). Product information rotenone. Available at: https://www.sigmaaldrich.com/content/dam/sigma-aldrich/docs/Sigma/Product_Information_Sheet/1/r8875pis.pdf Soares JVC (2013). Manejo da murcha de esclerócio (Sclerotium rolfsii Sacc.) em pimentão e seleção de acessos de Capsicum sp. resistente. Dissertation. Federal University of Amazonas. |
|
|
Soares JVC, Bentes JLS, Gasparotto L (2017). Reação de genótipos de Capsicum spp. à podridão do colo (Sclerotium rolfsii). Summa Phytopathologica 43(1):58-59. |
|
|
Upegui Y, Quiñones JGW, Torres F, Escobar G, Robledo SM, Echeverri F (2014). Preparation of rotenone derivatives and in vitro analysis of their antimalarial, antileishmanial and selective cytotoxic activities. Molecules 19(11):18911-18922 |
|
|
Valcárcel LE (1984). História del Perú antiguo através de las fuentes escritas. Lima, Peru: Juan Mejía. |
|
|
WHO-World Health Organization (1992) Rotenone health and safety guide. Geneva, Italy: Wissenschaftliche Verlagsgesdlschaft. |
|
|
Xavier VM, Message D, Picanço MC, Bacci L, Silva GA, Benevenute, JS (2010). Impact of botanical insecticides on indigenous stingless bees (Hymenoptera: Apidae). Sociobiology 56(3):713-725. |
|
|
Yu JH, Huang CF, Wang TH, Hung DZ, Mu HW, Pan CS (2020). Oxidative storm in a patient with acute rotenone-containing plant poisoning. The American Journal of Emergency Medical 38(6):1296e1-1296e3. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


