Full Length Research Paper

ABSTRACT
This study evaluated the effect of ginger rhizome and its callus as well as callus elicited by yeast extract, glycine and salicylic acid on the production of pro-inflammatory (TNF-α, IL-1 and IL-6) and anti-inflammatory (IL-10 and TGF-β) cytokines in vitro. Petroleum ether (PE) and chloroform; methanol (1:1) (CM) extracts of rhizome and callus were shown to significantly (P < 0.05) suppress in a dose-dependent manner the LPS-induced production of TNF-α, IL-1 and IL-6. Both callus extracts showed significantly (P < 0.05) higher ability than the rhizome extracts. CM extract of ginger callus treated with elicitors showed significant (p < 0.05) capacity to inhibit IL-1, IL-6 and TNF-α secretion at highest concentration used (100 μg/mL) when compared to control (untreated callus). Elicitors improved significantly (P < 0.05) the callus capacity to produce the IL-10 and TGF-β anti-inflammatory cytokines. HPLC analysis showed that 6-gingerol and 6-shogaol were found in both extracts of rhizome, but were not detected in the callus extracts. Furthermore, gallic acid was found only in CM extracts of rhizome (34.05 ± 0.39 μg/mg) and callus (17.88 ± 0.01 μg/mg). Yeast extract, salicylic acid and glycine elicitors enhanced significantly (p < 0.05) the production of gallic acid in callus CM extract where the highest content was obtained from callus elicited with 100 mg/L of yeast extract followed by callus elicited with 50 mg/L of salicylic acid and 200 mg/L of glycine, respectively. Therefore, ginger callus could be included in nutraceutical formulations where it could provide valuable protection against inflammatory diseases.
Key words: Anti-inflammatory, ginger, callus, elicitors.
INTRODUCTION
Zingiber officinale or ginger (Zingiberaceae), is indigenous to tropical Asia, and occur particularly in southern China and India. It was introduced by the Arab traders to Africa continent (Semwal et al. 2015). In Yemen it is used as an aphrodisiac and an aromatic stimulant (Fleurentin and Pelt, 1982). A number of scientific studies have proven the biological properties of ginger such as antioxidant (Stoilova et al., 2007; Mošovskáa et al., 2015), anti-inflammatory (Minghetti et al., 2007; Barari, 2016), anticancer (Cheng et al., 2011) and anti-diabetic (Afshari et al., 2007) activities. The safety of Z. officinale has been investigated. Acute and chronic toxicity studies have demonstrated a broad safety of Z. officinale (Daily et al., 2015).The nutraceutical value of ginger is mainly attributed to gingerols, shogaols and zingiberene (Butt and Sultan, 2011).
Plant tissue and cell suspension cultures have been used to produce interesting bioactive secondary metabolites (Sák et al., 2014). Furthermore, it is reported that application of elicitors like salicylic acid, yeast extract and glycine induces the production of bioactive molecules from in vitro cultures (Namdeo, 2007). Elicitors including yeast extract, glycine and salicylic acid augmented significantly the total phenolic content of ginger callus, and consequently its antioxidant capacity (Ali et al., 2018).
The use of ginger infusions for relieving the pain of rheumatism and arthritis have prompted researchers to investigate the anti-inflammatory activity of ginger and the chemical compounds responsible for this activity (Tripathi et al., 2008; Ahn et al., 2009; Semwal et al., 2015; Srinivasan, 2017). However, there is no report on the effect of ginger callus or its callus treated with specific elicitors on the production of cytokines which play the major role in the inflammatory process. The present study was performed to investigate the effect of ginger rhizome and its callus/treated callus with different concentrations of yeast extract, glycine and salicylic acid on the in vitro production of pro-inflammatory and anti-inflammatory cytokines.
MATERIALS AND METHODS
Plant materials
Fully matured rhizomes of ginger were collected from the botanical garden at the Biology and Biotechnology Department, Faculty of Science and Technology, Al-Neelain University, Khartoum, Sudan. Rhizomes were well cleaned, cut into thin slices, and dried at room temperature.
Callus induction and proliferation
Ginger callus was initiated and proliferated as described by Ali et al. (2016). Highest fresh weight of callus was induced from shoot tip explants on MS medium fortified with 0.5 mg/L 2,4-dichloro-phenoxyacetic acid (2,4-D) and proliferated on the same medium and concentration of 2,4-D.
Elicitor’s treatments
Callus elicited by yeast extract (100, 300 and 500 mg/L), glycine (100, 200 and 300 mg/L) and salicylic acid (100 and 200 mg/L) was prepared using the protocol published by Ali et al. (2018).
Preparation of extracts
Two types of extracts were prepared by sequential maceration of each sample (rhizome, callus and treated callus) in petroleum ether (PE) and chloroform: methanol (1:1, v/v) (CM) (Ali et al., 2018).
Cell culture
THP cells (human acute monocytic leukemia cell line) were acquired from American type culture collection. Culture medium was Dulbecco’s modified eagle’s medium (DMEM) containing glucose (4.5 g/L), fetal bovine serum (10% v/v), nonessential amino acids (1%), glutamine (1%), streptomycin (10 μg/mL) and penicillin (100 U/mL). Cell line was cultured at 37°C with 5% humidified CO2. Cells were activated with Vitamin D3 (0.1 μM) and PMA (100 ng/mL). After that, the cells were exposed to different concentrations (25, 50 and100 μg /mL) of ginger rhizome, callus, treated callus extracts and 50 μg/mL of 6-gingerol and 6-shogaol in a fresh serum-free medium in the presence of LPS (1 μg/mL).
Cytokine enzyme linked immunosorbent assay (ELISA)
Sterile solutions of samples (DMSO dissolved) were incubated with THP cells for 72 h. Afterward, the levels of various cytokines in culture supernatants were measured using an ELISA via cytokine ELISA set (R and D Systems, USA). ELISA was performed according to the manufacturer’s protocol. The absorbances were read at 450 nm with a microtiter plate reader (Genetix GMB-580). Cytokine levels, expressed as pg/mL, were calculated from the standard curve (Ahmad et al., 2017).
HPLC analysis of ginger rhizome and callus extracts
Preparation of sample solutions
Accurate weight (20 mg) of ginger rhizome and callus extracts was dissolved separately in HPLC grade chloroform: Methanol 1:1 (10 mL). Volume of extracts was prepared with methanol up to 20 mL to give stock solutions (1 mg/mL), and filtered through 0.45 μ filter. Further, all solutions were subjected to HPLC analysis for simultaneous estimation of 6-Gingerol, 6-Shogaol and gallic acid. To prepare the standards solution, 6-gingerol, 6-shogaol and gallic acid (0.0625, 0.125, 0.250, 0.500 and 1 mg/mL) were dissolved in HPLC grade methanol. All standards solutions were stored at 4°C until used.
HPLC conditions
Analysis of ginger rhizome and callus extracts, along with reference compounds was performed using a gradient HPLC system (Waters, USA) as described by Mahadik et al. (2013). Liquid chromatographic separations were performed on an Inertsil ODS-3 column with a dimension of 250 x 4.6 mm, 5 μm particles size (Thermo Scientific, Waltham, USA). The mobile phase was consisting of acetonitrile and (0.05%) ortho-phospohoric acid (85:15, v/v). Flow rate was 1.0 mL/min, column temperature was 30°C and detection wavelength was set at 227 nm. All operations and data analysis were controlled using the Chemstation software (Empower). Identification of the three compounds in the extracts was achieved by comparison of their retention times with standards, UV spectra and UV absorbance ratios after co-injection of samples and standards. The content of 6-gingerol, 6-shogaol and gallic acid were calculated by using the standard calibration curves. The calibration curves of 6-gingerol, 6-shogaol and gallic acid were constructed with the correlation coefficients (R2) 0.9998, 0.9983 and 0.9994 respectively and regression equations; y=24356x + 296180, y=46021x - 108135 and y= 41274x + 155145 respectively.
Statistical analysis
All data were analyzed using SPSS version 19. The values were expressed as mean ± standard deviation (SD). Significant differences between samples were detected by analysis of variance (ANOVA) followed by Duncan’s multiple-range test (p < 0.05).
RESULTS AND DISCUSSION
Effect of ginger rhizome and callus extracts on pro-inflammatory and anti-inflammatory cytokines
Biological therapies for inflammatory diseases involve generally the suppression of members of the inflammatory cascade, like cytokines (Kim et al., 2013). In this study, PE and CM extracts of ginger rhizome and callus induced a dose-dependent inhibition of the production of pro-inflammatory cytokines (TNF-α, IL-1 and IL-6) by LPS activated macrophages.
Rhizome PE and CM extracts displayed more or less the same capacity to inhibit TNF-α. However, both callus extracts showed significantly (p < 0.05) higher activity than the rhizome extracts. At highest concentration (100 μg/mL) the activity of the PE and CM callus extracts to inhibit TNF-α production increased significantly (P < 0.05) by 19 and 58% respectively than their corresponding rhizome extracts as depicted in Figure 1A. Moreover, both rhizome extracts displayed significant (P < 0.05) capacity to inhibit IL-1 production with higher activity observed for PE. The ability of callus PE and CM extracts to inhibit IL-1 at concentration 100 μg/mL, increased significantly (P < 0.05) by 30.4 and 61.2% respectively compared to their corresponding rhizome extracts as shown in Figure 1B.
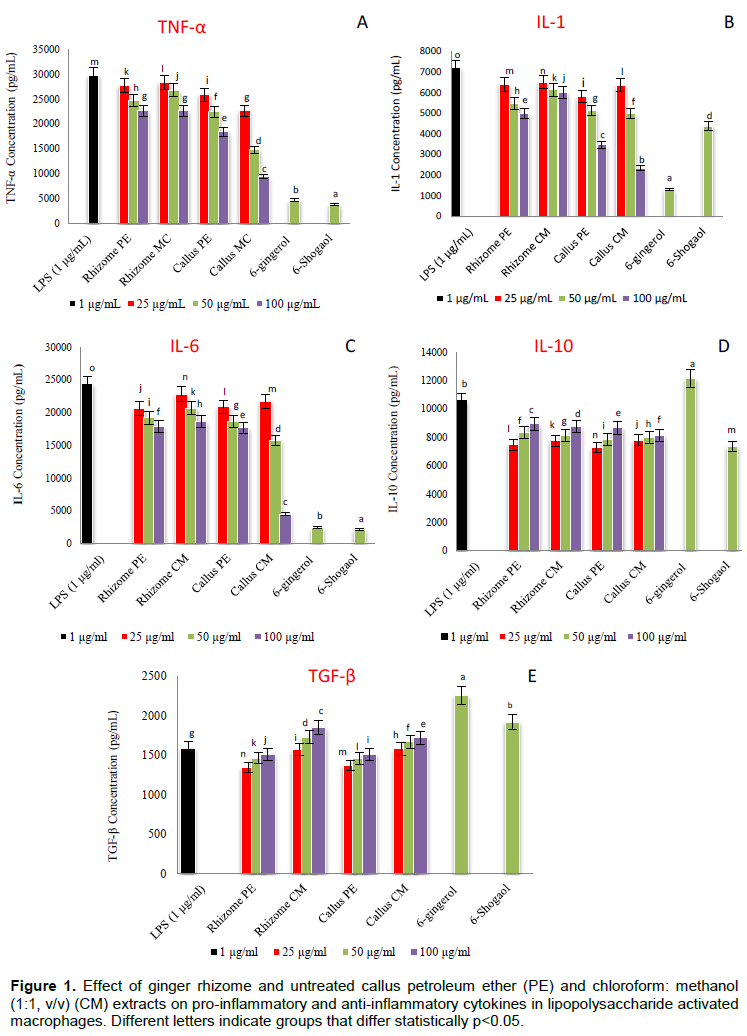
PE extracts of ginger rhizome and callus displayed more or less the same capacity to inhibit the IL-6 production; however, the inhibition capacity for IL-6 production of the CM callus extract at concentrations 50 and 100 μg/mL, increased significantly (P < 0.05) by 23.8 and 75.9% respectively than those of the rhizome as presented in Figure 1C. TGF-β involved in cell growth and differentiation where it is known to act as a multifunctional cytokine (Derynck et al., 2001). In addition, IL-10 and TGF-β are known to play an important role in tolerance induction and immune regulation (Van Kooyk et al., 2004). In this study, extracts of ginger rhizome and callus did not induce the production of IL-10 while CM extracts of rhizome and callus slightly, but not significantly, induced the production of TGF-β anti-inflammatory cytokine. These results supported earlier reports on the ginger anti-inflammatory effect. However, many previous studies showed that gingerols were the most typical active components contributing to the anti-inflammatory effect of ginger (Tripathi et al., 2008; Ahn et al., 2009; Semwal et al., 2015).
Some other studies demonstrated that shogaols were more potent than gingerols (Dugasani et al., 2010). Ha et al. (2012) reported that 6-shogaol displayed higher anti-inflammatory activity than 6-gingerol by inhibiting prostaglandin E2 and pro-inflammatory cytokines production. Other researchers like Ho et al. (2013) showed that 10-Gingerol as well as shogaols inhibited neuro-inflammation in a LPS-activated BV2 microglia. In our study, 6-gingerol and 6-shogaol showed strong significant (P < 0.05) activity to inhibit TNF-α, IL-1 and IL-6 production with higher capacity for 6-gingerol towards IL-1 inhibition.
Moreover, 6-gingerol promoted significantly (P < 0.05) the production of IL-10 and TGF-β, while 6-shogaol induced only TGF-β production and with lower effect than 6-gingerol as shown in Figure 1 D and F. It was also observed that the rhizome extracts possessed lower activity than the pure 6-gingerol and 6-shogaol suggesting the presence of molecules in rhizome that may interfere with activation of these compounds and their ability to suppress the production of pro- inflammatory cytokines; IL-1, IL-6 and TNF-α. Although 6-gingerol was not detected in both callus extracts, the callus showed significant (P < 0.05) higher capacity to inhibit the production of pro-inflammatory cytokines than that exhibited by the rhizome extracts suggesting the presence of other anti-inflammatory agents. The effect of callus tissues treated with different levels of yeast extract, glycine and salicylic acid elicitors on anti-inflammatory activity was also carried out and results are presented in Figure 2
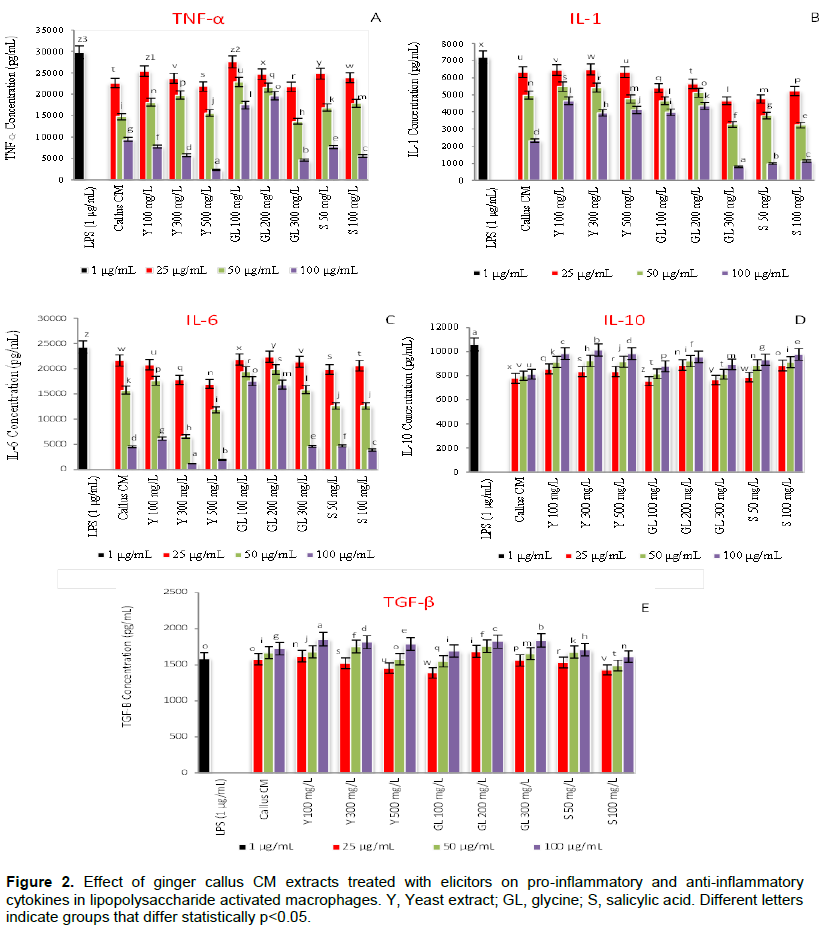
In general, CM extract of ginger callus treated with elicitors showed significant (P < 0.05) capacity to inhibit the production of pro-inflammatory cytokines (IL-1, IL-6 and TNF-α) at highest concentration used (100 μg/mL) compared to untreated callus. Callus elicited with 100, 300, 500 mg/L yeast extracts improved significantly (P < 0.05) its capacity, at 100 μg/mL to inhibit TNF-α production by 18, 39 and 75% respectively, while callus elicited with 50, 100 mg/L salicylic acid enhanced significantly (P < 0.05) its capacity, at 100 μg/mL by 19 and 41% respectively. However, the glycine only showed significant (P < 0.05) effect at concentration 300 mg/L where it improved the inhibition capacity of callus to produce TNF-α cytokine by 51% as presented in Figure 2A. On the other hand, glycine at 300 mg/L and salicylic acid (50, 100 mg/L) elicitors improved significantly (P < 0.05) the callus capacity, at 100 μg/mL to inhibit IL-1 production by 66, 58 and 52% respectively, as shown in Figure 2B.
Inhibition of IL-6 production was significantly (P < 0.05) obtained from callus elicited with 300 and 500 mg/L yeast and 100 mg/(P < 0.05) salicylic acid where it increased the callus capacity by 74, 59 and 14% respectively, as presented in Figure 2C. Interestingly, elicitors improved the callus capacity to produce the IL-10 and TGF-β anti-inflammatory cytokines. Callus treated with yeast extract (100, 300, 500 mg/L) increased significantly (P < 0.05) the production of IL-10 in a concentration dependent manner where highest increase (21, 25 and 21% respectively) was obtained at 100 mg/L of CM extract. Glycine (100, 200, 300 mg/L) improved significantly (P < 0.05) the IL-10 production of callus by 8, 18 and 10% respectively, while salicylic acid (50, 100 mg/L) by 15 and 20% respectively as shown in Figure 2D.
However, yeast (100, 300, 500 mg/L) and glycine (200 and 300 mg/L) elicitors slightly enhanced the capacity of callus at concentration 100 mg/L of CM extract to produce TGF-β anti-inflammatory cytokine by 8, 5 and 4% respectively for yeast elicitor and by 6 and 7% respectively for glycine elicitor as depicted in Figure 2E. These results suggested that the type and concentration of elicitor may have influenced the secondary metabolite synthesis and thus influence their anti-inflammatory potentiality (Karwasara et al., 2011).
HPLC profile of ginger rhizome and callus extracts
Qualitative and quantitative analysis for presence of 6-gingerol, 6-shogaol and gallic acid in ginger rhizome and callus extracts was also carried out using HPLC, and results are illustrated in Figures 3 and 4. Comparing the chromatograms of rhizome extracts with that of the standards, it was observed that 6-gingerol and 6-shogaol were only detected in both extracts (PE and CM) of ginger rhizome, while callus extracts were devoid of these two standards as illustrated in Figure 3. 6-gingerol was higher than that of 6-shogaol where the former had the highest concentration (191.21 ± 0.00 μg/mg) in the PE extract of the rhizome and the latter displayed highest content (80.46 ± 0.02 μg/mg) in the CM extract as illustrated in Table 1.
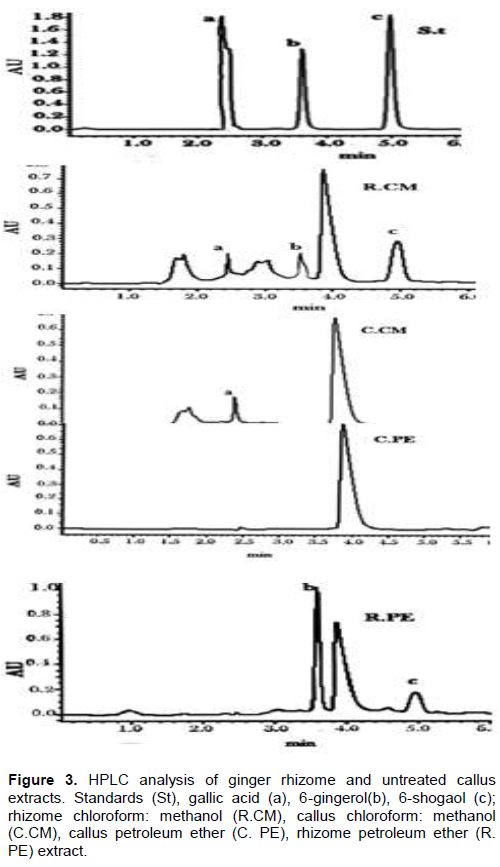
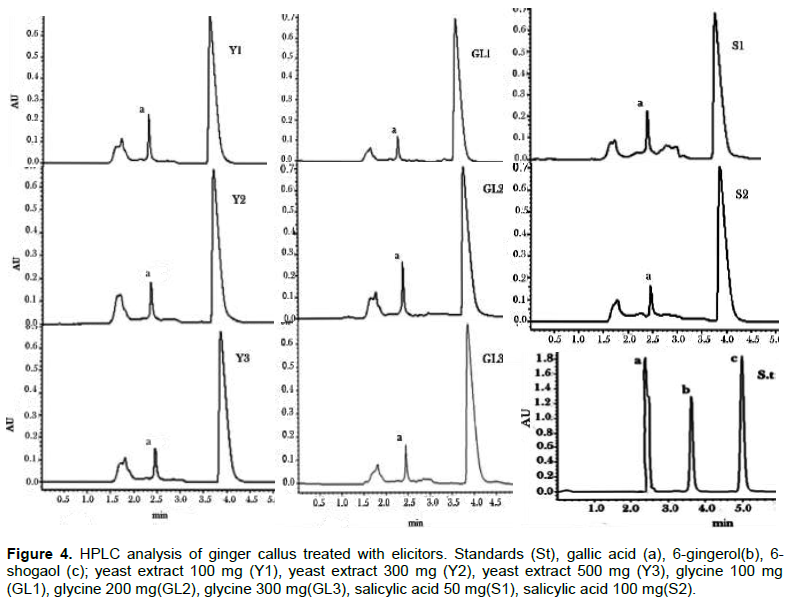
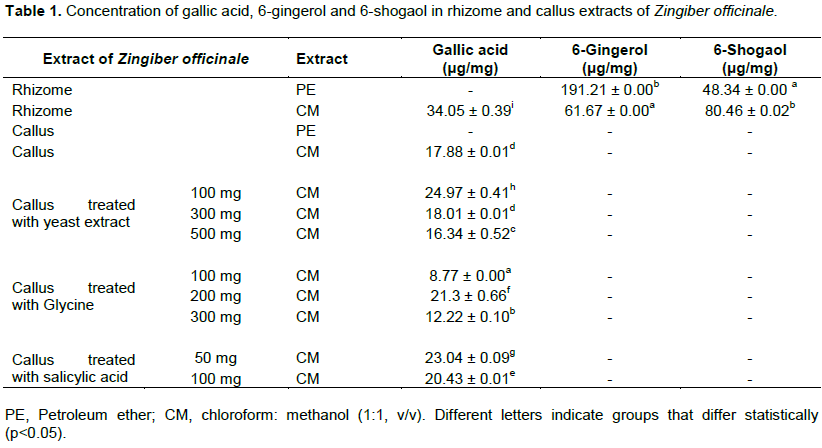
These results were consistent with those reported by Kizhakkayil and Sasikumar (2012) who determined the levels of gingerols and shogaols in the oils of 46 ginger accessions using HPLC. They found that 6- gingerol was the predominant one among the identified gingerols and 6-shogaol was also present in all the samples but with relatively low concentration when compared with 6-gingerol. The absence of 6-gingerol and 6-shogaol incallus extracts might be attributed to the dedifferentiation of callus culture. Pawar et al. (2015) investigated the content of 6-gingerol in ginger callus, micro-propagated rhizome and rhizomes growing conventionally using HPLC analysis. They found that, callus culture and micro-propagated plants contained lower amounts of 6-gingerol than that amounted in conventionally grown plants. These results suggested that, the dedifferentiation of callus and cells cultured in vitro is often associated with reduction of secondary metabolites content.
Furthermore, HPLC analysis of ginger rhizome and callus extracts showed that gallic acid was found in CM extracts of rhizome (34.05 ± 0.39 μg/mg) and callus (17.88 ± 0.01 μg/mg) and was not detected in their PE extracts as illustrated in Table 1. Gallic acid was reported as one of the most abundant phenolic acid in ginger (Ghasemzadeh and Ghasemzadeh, 2011). Treatment of callus with different concentrations of elicitors (yeast extract, glycine and salicylic acid) showed variable effect on the production of gallic acid in callus CM extracts as illustrated in Table 1 and Figure 4.
Highest content in gallic acid was obtained in callus elicited with yeast at concentration 100 mg/L where gallic acid content production increased significantly (P < 0.05) by 40% when compared with untreated callus. Elicitation of callus with yeast at concentration 300 mg/L did not affect the production of gallic acid while at higher concentration (500 mg/L), the production of gallic acid was reduced slightly. The effect of yeast extract, which is known as biotic elicitors mainly composed of amino acids, minerals and vitamins is attributed to its role in increasing the activity of phenylalanine ammonia lyase, a key enzyme of phenylpropanoid pathway that catalyses’ deamination of L-phenylalanine and trans-cinnamic acid production that links primary metabolism to the secondary one, and consequently the formation of vast secondary metabolites with phenylpropanoid skeleton (Seidel et al., 2002). Furthermore, the elicitation effects of yeast extract might be due to the presence of cations such as Ca, Zn and Co (El-Nabarawy et al., 2015).
Elicitation of callus with glycine showed that only at concentration 200 mg/L the production of gallic acid was increased significantly (P < 0.05) by 19%, while it reduced significantly (P < 0.05) at concentrations 100 and 300 mg/L. Molnár et al. (2011) reported that some amino acids like glycine, arginine and lysine, and vitamins such as nicotinic acid and thiamine, could serve as replacements for yeast extract, where it enhances growth in media containing relatively low concentrations of nitrogen or where vitamins are lacking.
The production of gallic acid was increased significantly (P < 0.05) by 29 and 14% in callus elicited with salicylic acid at concentrations 50 and 100 mg/L respectively. This result was in agreement with that of Ghasemzadeh and Jaafar (2012) who found that the content of phenolic acids including gallic acid increased significantly in two ginger varieties treated with different concentrations of salicylic acid. Moreover, they observed that the increase in phenolic compounds was associated with an increase in total soluble carbohydrate and a decrease in total flavonoids.
Thus, it was clear that yeast, salicylic acid and glycine elicitors enhanced the production of gallic acid where the highest content was obtained from callus elicited with 100 mg/L of yeast followed by callus elicited with 50 mg/L of salicylic acid and 200 mg/L of glycine respectively. Thus, it could be suggested that gallic acid participated in the anti-inflammatory activity of ginger callus.
CONCLUSION
In summary, the data of the present study demonstrated that, callus CM extract showed significant (P < 0.05) higher ability than the rhizome extracts to inhibit TNF-α, IL-1 and IL-6 production. Gallic acid was found in rhizome and callus while 6-ginger and 6-shogaol were only detected in the rhizome. Yeast, salicylic acid and glycine elicitors enhanced the production of gallic acid in callus and consequently its ability to modulate pro-inflammatory and anti-inflammatory cytokines release.
Furthermore, the results suggested that in addition to gingerols and shogaols, gallic acid might also contribute in the immunomodulatory effect of ginger rhizome, while in callus; gallic acid might play a major role in their anti-inflammatory activity. We speculate that, ginger callus could be included in nutraceutical formulations where it could provide valuable protection against inflammatory diseases.
Higher amounts of biomass accumulation could be obtained in cell suspension culture of ginger on a commercial level. Other biotic elicitors like glycoproteins and abiotic elicitors like ultraviolet radiation and heavy metals could be tested in order to generate more secondary metabolism from ginger culture for pharmaceutical and food industries.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
The authors are grateful to Dr. Alweya Abdallah (Faculty of Science and Technology, Al-Neelain University, Khartoum, Sudan) for taxonomic identification of the plant.
REFERENCES
|
Afshari AT, Shirpoor A, Farshid A, Saadatian R, Rasmi Y, Saboory E (2007). The effect of ginger on diabetic nephropathy, plasma antioxidant capacity and lipid peroxidation in rats. Food Chemistry 101:148-153. |
|
|
Ahmad F, Zubair S, Gupta P, Gupta UD, Patel R, Owais M (2017). Evaluation of aggregated Ag85B antigen for its biophysical properties, immunogenicity, and vaccination potential in a murine model of tuberculosis infection. Frontiers in Immunology 8:1-19. |
|
|
Ahn SI, Lee JK, Youn HS (2009). Inhibition of homodimerization of toll-like receptor 4 by 6-shogaol. Molecules and Cells 27:211-215. |
|
|
Ali AMA, El-Nour M EM, Yagi SM (2018). Total phenolic and flavonoid contents and antioxidant activity of ginger (Zingiber officinale Rosc.) rhizome, callus and callus treated with some elicitors. Journal of Genetic Engineering and Biotechnology 16:67-682. |
|
|
Ali AMA, El-Nour MEM, Yagi SM (2016). Callus induction, direct and indirect organogenesis of ginger (Zingiber officinale Rosc). African Journal of Biotechnology 38:2106-2114. |
|
|
Barari AR (2016). Effect of ginger supplementation and endurance training on serum levels of IL-1α and TNF-α. Medical Laboratory Journal 10:58-63. |
|
|
Butt MS, Sultan MT (2011). Ginger and its health claims: molecular aspects. Critical reviews in food Science and Nutrition 51(5):383-393. |
|
|
Cheng X, Liu Q, Peng Y, Qi L, Li P (2011). Steamed ginger (Zingiber officinale): Changed chemical profile and increased anticancer potential. Food Chemistry 129(4):1785-1792. |
|
|
Daily JW, Yang M, Kim DS, Park S (2015). Efficacy of ginger for treating Type 2 diabetes: A systematic review and meta-analysis of randomized clinical trials. Journal of Ethnic Foods 2:36-43. |
|
|
Derynck R, Akhurst RJ, Balmain A (2001). TGF-beta signaling in tumor suppression and cancer progression. Nature Genetics 29:117-129. |
|
|
Dugasani S, Pichika MR, Nadarajah VD, Balijepalli MK, Tandra S, Korlakunta JN (2010). Comparative antioxidant and anti-inflammatory effects of [6]-gingerol, [8]-gingerol, [10]-gingerol and [6]-shogaol. Journal of Ethnopharmacology 127:515-520. |
|
|
El-Nabarawy MA, El-Kafafi SH, Hamza MA, Omar MA (2015). The effect of some factors on stimulating the growth and production of active substances in Zingiber officinale callus cultures. Annals of Agricultural Sciences 1:1-9. |
|
|
Fleurentin J, Pelt JM (1982). Repertory of drugs and medicinal plants of Yemen. Journal of Ethnopharmacology 6:85-108. |
|
|
Ghasemzadeh A, Ghasemzadeh N (2011). Effects of shading on synthesis and accumulation of polyphenolic compounds in ginger (Zingiber officinale Roscoe) varieties. Journal of Medicinal Plants Research 5(11):2435-2441. |
|
|
Ghasemzadeh A, Jaafar HZE (2012). Effect of salicylic acid application on biochemical changes in ginger (Zingiber officinale Roscoe). Journal of Medicinal Plants Research 6(5):790-795. |
|
|
Ha SK, Moon E, Ju MS, Kim DH, Ryu JH, Oh MS, Kim SY (2012). 6-Shogaol, a ginger product, modulates neuroinflammation: a new approach to neuroprotection. Neuropharmacology 63(2):211-223. |
|
|
Ho SC, Chang KS, Lin CC (2013). Anti-neuroinflammatory capacity of fresh ginger is attributed mainly to 10-gingerol. Food Chemistry 141:3183-3191. |
|
|
Karwasara VS, Tomar P, Dixit VK (2011). Influence of fungal elicitation on glycyrrhizin production in transformed cell cultures of Abrus precatorius L. Pharmacognosy Magazine 7:307-313. |
|
|
Kim Y, Lee SK, Bae S, Kim H, Park Y, Chu NK (2013). The anti-inflammatory effect of alloferon on UVB-induced skin inflammation through the down-regulation of pro-inflammatory cytokines. Immunology Letters 149:110-118. |
|
|
Kizhakkayil J, Sasikumar B(2012). Characterization of ginger (Zingiber officinale Rosc.) germplasm based on volatile and non-volatile components. African Journal of Biotechnology 11(4):777-786. |
|
|
Mahadik KR, Nikam AR Sathiyanarayanan L (2013). Validation of reversed-phase high-performance liquid chromatography method for simultaneous determination of 6-,8-, and 10-shogaol from ginger preparations. International Journal of Pharmacy and Pharmceuetical Science 5:432-437. |
|
|
Minghetti P, Sosaz S, Cilurzo F, Casiraghi A, Alberti E, Tubaro A (2007). Evaluation of the topical anti-inflammatory activity of ginger dry extracts from solutions and plasters. Planta Medica 73:1525-1530. |
|
|
Molnár Z, Virág E, Ördög V (2011). Natural substances in tissue culture media of higher plants. Acta Biologica Szegediensis 1:123-127. |
|
|
Mošovskáa S, Novákováa D, Kaliňákb M (2015). Antioxidant activity of ginger extract and identification of its active components. Acta Chimica Slovaca 8:115-119. |
|
|
Namdeo AG (2007). Plant cell elicitation for production of secondary metabolites: A review. Pharmacognosy Reviews 1:69-79. |
|
|
Pawar P, Pai S, Nimbalkar M, Dixit G (2015). RP-HPLC analysis of phenolic antioxidant compound 6-gingerol from in vitro cultures of Zingiber officinale Roscoe. Plant Science Today 1:24-28. |
|
|
Sák M, Dokupilova' I, Mihalik D, Lakatosova J, Gubisova M, Kraic J (2014). Elicitation of phenolic compounds in cell culture of Vitis vinifera L. by phaeomoniella chlamydospora. Nova Biotechnologica et Chimica 2:162-171. |
|
|
Seidel V, Windho¨ vel J, Eaton G, Alfermann AW, Arroo RRJ, Medarde M, Petersen M, Wolley JG (2002). Biosynthesis of podophyllotoxin in Linum album cell cultures. Planta 215:1013-1039. |
|
|
Semwal R B, Semwal, D K, Combrinck, S, Viljoen, AM (2015). Gingerols and shogaols:Important nutraceutical principles from ginger. Phytochemistry 117:554-568. |
|
|
Srinivasan K (2017). Ginger rhizomes (Zingiber officinale): A spice with multiple health beneficial potentials. Pharma Nutrition 5:18-28. |
|
|
Stoilova I, Krastanov A, Stoyanova A, Denev P, Gargova S (2007). Antioxidant activity of a ginger extract (Zingiber officinale). Food Chemistry 102:764-770. |
|
|
Tripathi S, Bruch D, Kittu DS (2008). Ginger extract inhibits LPS induced macrophage activation and function. BMC Complementary and Alternative Medicine 8:1-7. |
|
|
Van Kooyk Y, Engering A, Lekkerkerker AN, Ludwig IS, Geijtenbeek TB (2004). Pathogens use carbohydrates to escape immunity induced by dendritic cells. Current Opinion in Immunology 16:488-493. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


